

Summary
Preassembled Cas9 kullanan karmaşık Ribonükleoprotein (RNP) kesin, verimli genom düzenleme için güçlü bir yöntemdir. Burada, biz yararını hücreleri ve organizmalar, geniş aralığında birincil insan hücreleri ve her iki klasik de dahil olmak üzere ve model organizmalar gelişmekte olan vurgulayın.
Abstract
Siteye özgü ökaryotik genom CRISPR (kümelenmiş düzenli olarak interspaced kısa palindromik tekrarlar) ile düzenleme-Cas (CRISPR ilgili) sistemleri hızlı bir şekilde çok çeşitli biyolojik soruları takip araştırmacılar arasında bir olağan hale geldi. Kullanıcıların en sık Streptococcus pyogenes kolayca programlanmış bir rehber RNA (gRNA) ile birlikte elde edilen Cas9 protein istihdam. Bu bileşenleri hücrelerin içine tanıtıldı ve çift iplikçikli DNA (dsDNA) genom tamamlayıcı bir bölge ile eşleştirme bir Bankası enzim iki iplikçikli mola (DSB) oluşturmak için her iki ipliklerini cleaves. Sonraki onarım rasgele ekleme veya silme olayları (indels) veya deneyci tarafından sağlanan DNA mola yerinde birleşme yol açar.
Bir RNP oluşturmak için preassembled ve doğrudan hücrelere, teslim bir saflaştırılmış tek-Kılavuzu RNA ve Cas9 protein, kullanımı son derece verimli gen düzenleme elde etmek için güçlü bir yaklaşımdır. RNP düzenleme özellikle gen ekleme, sık sık elde etmek zordur bir sonucu oranını artırır. Bir plazmid üzerinden gönderim karşılaştırıldığında, Cas9 RNP hücre içinde daha kısa kalıcılığı daha az hedef kapalı olaylarına yol açar.
Avantajları rağmen CRISPR gen düzenleme birçok sıradan kullanıcılar bu tekniği ile daha az aşina bulunmaktadır. Giriş engel düşürmek için biz farklı uygulamalar ve farklı yararları vurgulayarak çeşitli bağlamlarda, RNP strateji uygulamak için detaylı iletişim kuralları özetlemektedir. Biz birincil insan hücreleri, T hücreleri ve hematopoetik kök/progenitor hücreler (HSPCs) iki tür düzenleme kapsar. Biz de ne kadar Cas9 sağlar düzenleme RNP modeli kabuklu, Parhyale hawaiensisson zamanlarda klasik modeli yuvarlak kurt Caenorhabditis elegans ve daha fazlasını içeren tüm organizmalar facile genetik manipülasyon tanıttı gösterir.
Introduction
Adına CRISPR-Cas9 sistemi herhangi bir genom1hedef bölgeleri değiştirmek bilim adamları sağlar. Bu hızlı ve ucuz teknoloji temel araştırma devrim yarattı ve kişiselleştirilmiş hastalığı tedaviler, hassas tarım ve2ötesinde geliştirme üzerinde derin bir etki yaratmak vaat ediyor. CRISPR düzenleme oluşturuluyordu bir araçtır ve sistem yeni bir laboratuarda uygulama genom mühendislik, sadece temel moleküler biyoloji becerileri hiçbir belirli uzmanlık gerektirir. Araştırmacılar şimdi daha önce dirençli organizmaların genetik manipülasyon3,4için birkaç alternatif yolları ile eğitim görebilirsiniz. Son beş yıl içinde tek başına, CRISPR genom düzenleme 200'den fazla farklı omurgalılar, omurgasız, bitki ve mikrobiyal türler mühendisi için kullanılmıştır.
CRISPR prokaryotik savunma yolu uyarlanmış, siteye özgü genom düzenleme için gerekli temel unsurları Cas9 protein, genellikle S. pyogenes vardır ve kodon-bir eklenen nükleer yerelleştirme sinyal (NLS) ve onun özel ile optimize RNA Kılavuzu5,6. Burada tartışılan değil de, diğer Cas9 orthologues veya CRISPR endonucleases de kullanılabilir. Doğal olarak meydana gelen gRNA iki ayrı ayrı kopya etmek parça, CRISPR RNA (crRNA) ve trans aktive oluşan crRNA (tracrRNA)7. Bu RNA'ların tek-Kılavuzu RNA (sgRNA)8olarak bilinen tek bir transkript içine erimiş. Çift-kılavuzu da olsa akıcı sgRNA9, genom düzenleyicilerinin çoğu seçin düzenli olarak kullanılan10,11. Denemecileri bir 20-nükleotit (nt) genomik DNA hedef, bu kısa bir lisans imza protospacer bitişik motifi (PAM) denilen Cas9 tanıma için gerekli yanında yatıyor sağlanması seçin ve tamamlayıcı sıra12 içerir bir gRNA Tasarla .
Bir kez hücre içinde karmaşık RNP genomik hedefini bulur, gRNA baz çifti ile tamamlayıcı DNA strand ve enzim cleaves iki iplikçikli oluşturmak için her iki DNA iplikçikleri2kır. Hücre onarım makineleri en az iki yoldan biri tarafından DSB giderir: hataya sigara-homolog son katılma (NHEJ) yol veya Homoloji yönetmen onarım (HDR) yolu ile hangi sorunsuz Homoloji her iki tarafında break ' silah' içeren DNA içermektedir. İkinci Denemecileri eklemek veya DNA dizileri1değiştirmek izin verirken eski onarım yolu genellikle INDEL oluşumu ve bunun sonucunda gene bozulma, yol açar.
Düzenleme verimliliği ve doğruluk hangi tarafından Cas9 ve gRNA hücre içine girmek anlamına gelir bağlıdır. Bu bileşenler kültürlü hücrelere, embriyo veya organizmalar nükleik asit veya formdaki bir preassembled RNP karmaşık13,14,15olarak teslim edilebilir. Yaygın nükleik asit alan teslim yöntem viral iletim, transfection veya Elektroporasyon mRNA veya plazmid DNA olabilir. Cas9 protein ve RNA Kılavuzu sonra hücre içinde üretilir ve onlar bir kompleks oluştururlar, ilişkilendirmek.
RNP doğrudan teslim Cas9 protein ve RNA Kılavuzu ayrı arıtma gerektirir. Bu-ebilmek kılınmak içinde-ev ya da protein ve sgRNA çeşitli ticari satıcılar birinden satın alınabilir. Bir kez satın aldı, Cas9 ve gRNA enzimatik yetkili RNP kompleks oluştururlar, karışık ve hücrelere döllenmiş yumurta/embriyo, lipid tabanlı transfection16veya Elektroporasyon içine direkt enjeksiyon tarafından tanıtıldı. RNP düzenleme ilk rapor C. elegans gonads17içine enjeksiyon dahil. Etkili Elektroporasyon fare18,19 ve sıçan20 embriyo yerleştirilmesi göstermiştir rağmen mikroenjeksiyon halen RNP embriyo ve bütün organizmalar, giriş tercih edilen araçtır. Biz doğrudan RNP C. elegans gonads ve P. hawaiensis embriyo enjekte için protokol tanımlamak ve Elektroporasyon birincil insan hücreleri düzenlerken RNP sunmak için özel bir tür öneririz. Bu yöntem, nucleofection, en iyi duruma getirilmiş Elektroporasyon programları ve hücre türüne özgü çözümler içerir ve RNP sitoplazma ve çekirdek21girmek izin verir.
Genom RNP ile düzenleme birkaç farklı avantajlar sunar. Protein ve RNA bileşenleri ön montajı ve kalitede teslim önce sağlanmış olur çünkü RNP düzenleme birçok tuzaklar nükleik asit tabanlı teslimatla ilişkili önler. Yani, ana bilgisayar genomu DNA Cas9 kodlama entegrasyonu riski yoktur, mRNA yıkımı için asla maruz ve in vivo gRNA veya protein ifade, katlama ve Derneği22,23sorunları kaçınmanızı sağlar. Ayrıca, RNP kullanarak alt toksisite ve plazmid tabanlı ifade, bir sonuç RNP'ın daha kısa Half-Life hücre24,25,26,27içinde daha çok daha az hedef dışı olaylar yol açar.
Son olarak, RNP düzenleme demonstrably yüksek düzenleme oranları insan hücre hatları, primer hücre çeşitli pluripotent kök hücreler (iSPCs), HSPCs, fibroblastlar, embriyonik kök hücre (ESCs), indüklenen ve16,24T hücreleri gibi yol açar, 25,26,27,28,29; omurgasızlar C. elegans, P. hawaiensisve meyve sinekleri3,17,30da dahil olmak üzere; zebra balığı, fare ve sıçan31,32gibi omurgalı türleri içinde; tür Arabidopsis, tütün, marul, pirinç, asma, apple, Mısır ve buğday33,34,35,36dahil bitki; ve içinde Chlamydomonas, Penicilliumve Candida türleri37,38,39. Plazmid teslim karşılaştırıldığında RNP kullanırken INDEL oluşumu sıklığı daha yüksek olabilir ve HDR-aracılı DNA ekleme25,27,29elde etmek daha kolay olabilir.
Burada açıklanan protokol Cas9 RNP kullanır ve çok çeşitli biyolojik sistemleri40,41, özellikle aksi halde çalışmak zor hücrelerdeki uygulamak basit bir etkili ve kolayca uyarlanabilir tekniği ile ve köklü sistemleri için kesin genetik manipülasyon olmadan organizmalarda. Biz tasarım, edinme ve farklı bir model hücre tipleri ve organizmalar arasında kullanımı kapsayan önce Cas9 RNP araya açıklayarak başlar. (HSPCs) hematopoietik kök/progenitor hücreler ve T hücreleri birlikte adım 2 ve 3 Bu protokol kapsamında nucleofection, aynı yöntemi kullanarak düzenlenmiş bulunmaktadır. Yordamlar Ciçin düzenleme. elegans adım 4 ve 5, açıklanan ve P. hawaiensis düzenleme 6 ve 7 numaralı adımları kaplıdır. Son olarak, herhangi bir organizma bir gen düzenleme deney başarısı genotip sıralama tarafından değerlendirildi bu yana, tüm hücreleri ve iletişim kuralında tanımlanan organizmalar için olası analiz yöntemlerinin açıklandığı adımlar 8. adımda özetlenmiştir.
Protocol
1. RNP derleme
-
De önceden, önceden tüm RNA, DNA ve protein bileşenleri edinme deney tasarlayın. Bir ilk geçiş olarak tablo 1'de listelenen olumlu denetimlerinden birini deneyin ve malzemeler tablo açıklanan ticari reaktifler güvenilir deneysel tasarım ve malzemelerin bütünlüğünü sağlamak için kullanın. Yeni bir genom düzenleme deneme planlama ek ipuçları için bkz: Bu konu12,42,43at kağıtları.
Not: sonraki adımlarda açıklandığı gibi monte sonra önceden hazırlanmış RNPs-80 ° C'de depolanmış olabilir- Hedef için hangi gen seçtikten sonra ücretsiz çevrimiçi araçlar biri bir en uygun gRNA44,45,46,47,48tasarlamak için kullanın. Bir nakavt oluşturmak için umut Eğer bir exon hedeflemek emin olun.
Not: Bir hedef site bir bitişik S. pyogenes ile PAM tanımlamak için bu araçları yardımcı olacak serisi, yüksek kalite puanı ve düşük hedef kapalı puanı. - S. pyogenes Cas9 protein ile yayınlanan yöntemleri8arındırmak veya ticari bir satıcıdan satın.
- Tipik bir Cas9 arabellek RNA seyreltme, RNP hazırlık ve 20 mM HEPES pH 7.5, 150 mM KCl, % 10 gliserol ve TCEP 1 mM içeren protein depolama için hazırlayın. Her zaman resuspend veya RNA yıkımı önlemek için sulandırmak için kullanılan arabellekleri nükleaz ücretsiz su kullanın.
- Yayınlanan yöntemleri kullanarak bir vitro transkripsiyon yoluyla RNA (tracrRNA ve crRNA veya sgRNA) Kılavuzu üretmek veya bir nükleik asit sentezi Şirket17,21,49, satın 50 , 51.
- Bir gen ekleme, sentez veya bir donör DNA şablon satın almak.
- Protein ve RNA aliquots-80 ° C ve tezcan buz kullanımdan hemen önce mağaza.
Not: Her donma-çözülme biraz verimini düşürür. Ayrıntılı, açık erişim protokolleri Cas9 arıtma52 ve sgRNAs53 , vitro transkripsiyonu için başka bir yerde kullanılabilir.
- Hedef için hangi gen seçtikten sonra ücretsiz çevrimiçi araçlar biri bir en uygun gRNA44,45,46,47,48tasarlamak için kullanın. Bir nakavt oluşturmak için umut Eğer bir exon hedeflemek emin olun.
- C. elegansile çalışıyorsanız, 1.5 adıma atlayın. P. hawaiensis iletişim kuralı için 1,6 adıma atlayın. SgRNA kullanıyorsanız, 1.4 adıma atlayın. Birincil hücre düzenleme için bir gRNA bir araya 1.3 adıma geçin.
-
Bir gRNA tracrRNA ve crRNA ekimolar miktarda karıştırılarak topla. Yaklaşık 50 genom düzenleme deneyler için 80 µM gRNA stok 100 µL olun.
- GRNA 30 dk 37 ° C'de kuluçkaya ve sonra yavaş yavaş oda sıcaklığına kadar soğumasını bekleyin.
-
HSPC ve T hücre düzenleme için RNP hazırlık: araya bir RNP karmaşık bir 1-2 x molar miktar gRNA toplam hacmi 10 µL. Cas9 proteinin 200 pmol için çok yavaş bir şekilde karıştırarak ekleyin konsantre Cas9 (Cas9 arabellekte önceden seyreltilmiş) gRNA için yaklaşık 30 için s , Hızlı daireler son Cas9 konsantrasyon 20 µM için getiren pipet, yapma.
- Elektroporasyon cuvettes hazırlayın.
Not: Bu protokolü Malzemeler tablobaşvurulan ticari sistem özgüdür, ancak RNP düzenleme de diğer Elektroporasyon cihazlar ile elde edilebilir. - 5 µL (100 pmols, T hücreleri) veya 10 µL (200 pmol, HSPCs) RNP her küvet için ekleyin.
- Bir eleme yapmak yerine yeni DNA'ları ekleme eklemek Eğer 100 µM (100 pmol) tek iplikçikli oligonükleotid donör DNA (ssODN)25,54,55 1 µL cuvettes veya kuyu plaka.
- Adım 2 için birincil hücre düzenleme sonraki bölümündeki yönergeleri atla protokolü.
- Elektroporasyon cuvettes hazırlayın.
-
C. elegans düzenleme RNP hazırlık: karmaşık RNP 20 µL (son konsantrasyonları parantez içinde belirtilmiştir) son bir birim oluşturmak için aşağıdaki ayıraçlar ekleyerek birleştirin: Cas9 (2 µM), HEPES pH 7.5 (10 µM), KCl (115 µM), crRNA (12 µM) , tracrRNA (40 µM) ve onarım şablonlar gerekirse (0.5 µM ssDNA veya 350 ng/µL dsDNA kadar).
Not: Cas9-aracılı DSB-şablonu esas alan bir onarım verimliliğini dsDNA onarım yapı konsantrasyonu orantılıdır; Bu nedenle, onarım şablon, daha verimli daha yüksek konsantrasyon şablonu esas alan onarım. Ancak, bir enjeksiyon 350 ng/µL dsDNA, büyük içeren karışımları canlılık enjekte solucanların azaltmak için gösterilmiştir. Böylece, e kadar en fazla 350 ng/µL dsDNA karışımında onun ölümcül en aza indirerek onarım verimliliği en üst düzeye çıkarmak için ama kullanmak en iyisidir.- Aynı anda birden fazla loci hedef, adım 5.4'açıklanan co-CRISPR/co-conversion tarama yaklaşım için gerektiği gibi için birden çok crRNAs ekleyin. Birden fazla crRNA eklerken, her ardışık olarak ana karışıma ekleyin.
Not: Her crRNA miktarı aynı olması gerekmez ve hatta crRNAs ana karışımında toplam konsantrasyonu Cas9 konsantrasyonu değiştirmeden iki katına mutagenesis belirli bir odağı, frekans ile müdahale değil gibi görünüyor. Örnekler Paix vd ayrıntılarıyla açıklanmıştır 56. - Pipetting tarafından mix ve 16.000 x g 5 için de RNP çözüm spin s çözüm tüpün dibinde toplanan emin olmak için.
- Çözüm için 15 m 37 ° C'de kuluçkaya.
- 16.000 x g ince-sıkılmış mikroenjeksiyon iğne yapışmasına neden herhangi bir parçacık cips 1 dk da örneğine santrifüj kapasitesi. Süpernatant sonraki adımları kullanın.
- Adım C. elegans Protokolü'nün 4'e kalan atlayın.
- Aynı anda birden fazla loci hedef, adım 5.4'açıklanan co-CRISPR/co-conversion tarama yaklaşım için gerektiği gibi için birden çok crRNAs ekleyin. Birden fazla crRNA eklerken, her ardışık olarak ana karışıma ekleyin.
-
P. hawaiensis düzenlemek için RNP hazırlık: tek kullanımlık Cas9 aliquots onları nükleaz ücretsiz su ve fenol (için enjeksiyonları görüntülenmesi) kırmızı Cas9 ve % 0.15 fenol kırmızı 6.25 µM son bir konsantrasyon sulandrarak hazırlamak.
- Karmaşık RNP 2-5 x 6 µl. Ekle 12 pmol Cas9, gRNA, 2 µM, 4-8 µM için gRNA konsantrasyon ve fenol red konsantrasyonu % 0.05 için son Cas9 konsantrasyon getirmek için toplam hacmi Cas9 protein gRNA molar fazlalığı karıştırılarak topla.
- Karışımı 10 dakika için karmaşık oda sıcaklığında RNP kuluçkaya.
- Adım 6 P. hawaiensis düzenleme sonraki talimatlar için atlaprotokolü.
2. hücre kültür ve hazırlık
Not: 2.1.1 için 3.3.3 biyolojik emanet dolabı adımları izleyerek.
-
Satınalma cryopreserved insan periferik kan CD34 seferber+ HSPCs bir satıcıdan.
- ~ 1 çözülme x106 HSPCs 37 ° C suda banyo 3 dakikadır ve 15 mL konik tüp aktarabilirsiniz. 10 mL serum-Alerjik genişleme orta ticari bir kaynaktan ekleyin ve 10 dk. süpernatant kaldırmak ve 2 mL ilave SFEM hücrelerde resuspend 100 x g karisimin spin. 6-iyi plakaları hücrelerde plaka ve onları 37 ° C kuluçka için 24-48 h RNP Elektroporasyon önce kültür.
- Bir hemasitometre içeren hücreleri saymak ve transfer HSPCs toplam sayısı (electroporated olmak 150.000-200.000 HSPCs küvet başına) bir santrifüj tüpü için gerekli.
- Tüp 100 x g hücreleri cips için 10 dakika için de spin.
-
Satın insan birincil CD4+ T hücreleri bir satıcıdan veya onları tarafından yoğunluk gradient Santrifüjü29bütün kan grubundan ayırmak.
- T Hücre aktivasyonu önce 48-şey kültür pilakalar ve αCD3 önceden kat (UCHT1) ve αCD28 (CD28.2). 10 µg/mL αCD3 ve en az 2 h 37 ° C'de için PBS içinde 10 µg/mL αCD28 500 µL pilakalar kat
Not: bazı loci için NHEJ öncesi stimülasyon elde edilebilir, ama verimlilik bu adımı da dahil olmak üzere en üst düzeye çıkarır. - ΑCD3/αCD28 antikor bağlı plakaları [RPMI-1640 ile HEPES, 5 mM 2 mM L-glutamin, ticari alternatif 50 µg/mL penisilin/streptomisin, 2-mercaptoethanol, 5 mM, 50 µM takıma bir RPMI tam orta 37 ° C'de 48 h için kültür T hücreleri gerekli olmayan amino asit, sodyum pyruvate ve % 10 (vol/vol) FBS 5 mM]. Medya 48-şey plaka kuyu başına 500 µL 2.000.000 T hücrelerinin bir yoğunluk itibariyle kültür T hücreleri.
- Bir hemasitometre ve transfer kullanarak hücreleri hesaplama T T Hücre çoğalmasıyla için gerekli toplam sayısı (electroporated olmak küvet başına 100.000-1.000.000 T hücreleri) bir santrifüj tüpü için denemeler yapın.
- 90 x g hücreleri cips için 8 dakika için de tüp spin. Hücreleri yoğunluğu 2 gün içinde degrade ayrılmış olması, onları vasıl 200 x g 8 min için spin.
- T Hücre aktivasyonu önce 48-şey kültür pilakalar ve αCD3 önceden kat (UCHT1) ve αCD28 (CD28.2). 10 µg/mL αCD3 ve en az 2 h 37 ° C'de için PBS içinde 10 µg/mL αCD28 500 µL pilakalar kat
-
Her iki hücre türü için süpernatant bir pipet/vakum, herhangi bir kabarcık çıkarma ile Aspire edin.
- Yavaşça Elektroporasyon arabelleği 20 µL hücrelerle küvet resuspend.
- Hücre (150.000-200.000 HSPCs veya 100.000-1.000.000 T hücreleri) 20 µL RNP ve mix 10 µL de yukarı ve aşağı kabarcıklar oluşturmadan pipetting tarafından zaten her küvet ekleyin.
3. RNP Elektroporasyon
- Electroporate bir nucleofector yerleştirerek sonra cuvettes. HSPCs için ER100 darbe kodu kullanın. T hücreler için EH-115 darbe kodu kullanın.
-
Yalnızca HSPCs: Eklemek 100 µL (37 ° C sıcak) ilave SFEM orta her küvet hemen sonra Elektroporasyon ve 10-15 dk. için hücreleri kurtarmak izin vermek için
- Onları bir 96-şey yuvarlak-alt plaka ve ilave SFEM orta 24 h için ek bir 100 µL eklemek kültür hücreleri aktarın.
- Onlara taze bir ilave SFEM orta olarak değiştirin ve onları bir ek 24-72 saat için kuluçkaya.
- Genotipleme için hücreleri kaldırmak onları 48-96 h mesaj-Elektroporasyon. Senaryo Özeti 300 x g 5 min için spin ve süpernatant DNA ekstraksiyon (adım 8.2) başlamadan önce kaldırın.
-
T hücreleri sadece: RPMI 80 µL tamamlamak kültür 37 ° C ile çok kanallı pipet (gerekirse) kullanarak havzanın her küvet veya iyi, önceden sıcak ortam ekleyin.
- Onları 15dk için 37 ° C'de kuluçkaya.
- Hedef plate(s) için uygun ortam, antikor, sitokinler, vb eklemek ve onları önceden 37 ° C kuluçka makinesine sıcak.
- Electroporated hücrelerinin 107 µL kuyulardan çok kanallı pipet (gerekirse) kullanarak yuvarlak-alt 96-şey plaka aktarın.
- Düzenleme sonuçları değerlendirme hakkında daha fazla bilgi için 8 adıma atlayın.
4. C. elegans hazırlık
-
mikroenjeksiyon öncesinde 1 gün: özel yastıkları mikroenjeksiyon için hazırlamak.
- %3 (w/v) özel çözüm özel su ekleyip kaynatın sıcak bir tabak veya bir mikrodalga çözüm getiren suda olun.
- 24 mm x 50 mm x 1.5 mm cam slayt bir tabloda düzenlemek ve bir cam Pasteur pipet özel çözüm bir slayda bir küçük (~ 15 µL) damla eklemenizi sağlar. Hızlı bir şekilde özel damla üstüne başka bir coverslip vermek suretiyle düzleştirin. Kuvvetlendirmek ve coverslips birini kaldırmak özel izin.
- Özel kaplamalı coverslip yüz-up bir masa üzerinde kuru gecede bırakın. 24 saat sonra özel yastıkları temiz, Kuru bir kapta saklayın.
Not: Bunlar süresiz olarak kullanılabilir.
- Mikroenjeksiyon iğneler çekin: borosilikat cam kılcal filamentler ile kullanarak (dış çap 1.0 mm ve iç çapı 0,58 mm), Mello ve yangın57 ve58diğer kaynaklara dayalı iğneler çekin. Şişleri hemen kullanılabilir veya kil destekler tarafından braced temiz, Kuru bir kap içinde saklı.
- Solucanlar bakımı için Petri kalıplara dökülür ve OP50 bakteri ile benekli Yuvarlak solucanlar büyüme medya (NGM) agar hazırlamak (Standart C. elegans protokolleri için bakım ve büyüme medya, tariflerini görmek Stiernagle59).
- Solucanlar mikroenjeksiyon için sahne: 12-24 h mikroenjeksiyon öncesinde L4 sahnelenen hermoafroditler için yeni bir NG-agar plaka OP50 bakteri ile almak ve onları bir gecede 20 ° C'de kuluçkaya Her Cas9 hedef/enjeksiyon karışım için ~ 30 solucanlar plaka almak.
-
Mikroenjeksiyon günü: RNP süpernatant 1.5 adımda hazırlanan solüsyona çekti mikroenjeksiyon iğneyle yük.
- Süpernatant kapiller pipet adımından 1.5.4 çekti kapiller pipet ve backfill çözüm içine üzerinden (genellikle daha az 0.1 µL takılması) hazırlanan mikroenjeksiyon iğne pipette.
- Bir micromanipulator için bağlı mikroenjeksiyon aparatı üzerine yüklenen iğne bağlayın. 250 kPa ve 25 kPa denge basınç enjeksiyon aparatı basınç ayarlı.
-
Geri yüklenen iğne ucu keskin iğne kenar oluşturmak için kır. Bir 15 x 15 mm x 1.5 mm kare coverslip bir 24 mm x 50 mm x 1.5 mm coverslip üstüne yerleştirin.
- Bir kare coverslip kenarına halocarbon yağla 700 yerleşimi.
- İğne 15 mm kare coverslip kenarında petrol yerleştirin.
- Yardım kullanma kılavuzu mikroskop sahne ve coverslip, fırça slaydı yukarı ve iğne kenarı boyunca enjeksiyon pedal/Farenizde süre için. İğne ucu geri, iğne dışında sıvı akışı artan kır. Bir en iyi akış hızı ~ 1 kabarcık/s şekillendirme iğne, kenarı boyunca akışı karıştırmak enjeksiyon yaparak elde.
- 12-24 h mikroenjeksiyon öncesinde aldı L4 solucanlar enjeksiyon günde gelişimsel olarak sahnelenen genç yetişkinler olduğunu onaylayın. Genç Yetişkin solucanlar OP50 bakteri yoksun bir NG-agar plaka almak ve 5 min için sürünmem kurmalarına izin. Bu iğne takunya minimize enjeksiyon yastık transfer bakteri miktarını azaltır.
- Bir özel enjeksiyon pad/coverslip bir diseksiyon kapsamı yerleştirin. Bir solucan pick kullanarak, küçük bir parça halocarbon petrol bir yastık kenarı boyunca yatıyordu.
-
Yağda kaplı solucan pick kullanarak, NG-agar plaka kapalı ve petrol kavramını içine birkaç solucanlar kaldırın. Bir kirpik ya da kedi bıyık gibi bir pipet bağlı güzel bir saç ile paralel, yavaşça solucanlar özel yastık iterek içinde solucanlar getirin. Mikroenjeksiyon yordamı ile rahat kadar sadece dağ ve bir solucan bir kerede enjekte.
Not: Kuru özel onları belgili tanımlık yastık için bağlı kalmak neden solucanlar, nemden fitil. Sonuç olarak, bir solucan yazılıolduğu gibi hızlı bir şekilde çalışmalıdır.- Bir kez içinde konumlandırın ve solucanlar solucan ucundan halocarbon yağ (~ 20 µL) birkaç damla almak başka bir ile belgili tanımlık yastık için bağlı, yer paylaşımı.
5. C. elegans erbezi mikroenjeksiyon ile RNPs ve sonrası enjeksiyon bakım
Not: Mikroenjeksiyon Protokolü Mello ve yangın57uyarlanmış ve60,61başka bir yerde ayrıntılı olarak açıklanmıştır.
-
Coverslip enjeksiyon mikroskop üzerine monte solucanlarla yerleştirin. Düşük büyütme (5 X amaç, 10 X oküler) altında enjeksiyon iğneyi dik solucanları getirin.
- Yüksek büyütme (40 X amaç, 10 X oküler) geçiş bitişik hücre çekirdeği içinde - için geç-pachytene yakınındaki bölgesine karşılık gelen erbezi kolu iğne yeniden konumlandırın.
- Micromanipulator kullanarak, manikür biraz iç karartıcı solucan karşı iğne taşıyın. Sonra bir elinizle manikür iğne sarsıntı için mikroskop sahne alanı'nın yan dokunun. Enjeksiyon pedal/düğmesine basın ve yavaş yavaş erbezi kol enjeksiyon karışımı ile doldurun ve iğne kaldırma.
- Erbezi kol ile bu işlemi tekrarlayın.
-
Bir zamanlar solucanlar enjekte edilir, coverslip/özel yastık kaldırmak ve diseksiyon mikroskop altında yerleştirin.
- Çekti kapiller pipet kullanarak, solucanlar yağdan bir M9 arabellek üzerlerine pipetting tarafından yerinden. Solucanlar agar üzerinden serbest bırakmak için bu tedavi gerçekleştirmek.
- Solucanlar arabellekte etrafında seyretmiştin zaman 10 dakika sonra onları çekti kapiller pipet kullanarak OP50 bakteri ile bir NG-agar plakasına taşıyın. Solucanlar iyileşti ve etrafında hareket ediyor kadar 2-3 h için 20 ° C'de plaka koyun.
- Sonra kurtarılan, ayrı ayrı ve plakaları için 25 ° C kuluçka aktarım solucanlar NG-ağar kaplamalar OP50 ile transfer.
-
P0izin-enjekte solucanlar büyümeye ve Döl 3 gün boyunca yatıyordu. Ekran F1 çoluk çocuk.
- Eğer Co-CRISPR veya ortak dönüşüm62,kullanarak63,64,65, sonra aday solucanlar referans gen mutasyona uğramış fenotip olup tabanlı tarama için seçin. Tek tek yeni NG-ağar kaplamalar OP50 ile işaretli bu solucanları aktarmak ve F2 Döl 20 ° C'de düzenlemek izin
Not: bir co-CRISPR tarama veya seçimi için kullanılan fenotip Cas9 düzenleme başarı için erken tahmini sağlamalıdır. - Co-CRISPR fenotip yoksa, mikroenjeksiyon verimliliği geliştirmeye yardımcı olmak için bir olumlu denetim plazmid microinject.
Not: Örneğin, mCherry öğesini MYO-2 kodlar enjeksiyon karışımında bir plazmid dahil enjeksiyon etkinliğini değerlendirmek yardımcı olacaktır. Başarılı bir şekilde pCFJ90 ile enjekte solucanlar bazı yavrular ile floresan pharynxes olacaktır.
- Eğer Co-CRISPR veya ortak dönüşüm62,kullanarak63,64,65, sonra aday solucanlar referans gen mutasyona uğramış fenotip olup tabanlı tarama için seçin. Tek tek yeni NG-ağar kaplamalar OP50 ile işaretli bu solucanları aktarmak ve F2 Döl 20 ° C'de düzenlemek izin
- F1 solucanlar istediğiniz düzenlemeleri varlığı için inceleyin. F1 anne bir 96-şey plaka tek tek bir şey için almak, onu parçalayıcı ve DNA'sı Ekle özel PCR güçlendirme, DNA dizi analizi veya arazi nükleaz tahlil (CEL-1)66inceleyin.
Not: Bu deneyleri bir co-CRISPR/co-conversion veya diğer tarama ya da seçim rejimler65,66,67,68kullanılırken gerçekleştirilebilir. - Düzenleme sonuçları değerlendirme hakkında daha fazla bilgi için 8 adıma atlayın.
6. P. hawaiensis hazırlık
- 1 gün önce mikroenjeksiyon, önceki gece bir 'çift tankı' ayarlayarak için erken embriyo zenginleştirmek; yeni ayrılmış kadın taze döllenmiş embriyo içerir. Rehm et al. bkz: 69 detaylar için.
- Mikroenjeksiyon günü, tek hücreli Parhyale embriyo toplamak (0-4 s sonrası döllenme) %0.02 karanfil yağı deniz suyu ile gravid kadın anesthetizing ve yavaşça embriyo ventral onu damızlık kese alev çekti kullanarak dışarı kazıma ve yuvarlak cam pipet ve #3 forseps sıkıcı bir çift.
7. P. hawaiensis embriyo mikroenjeksiyon ile RNPs ve sonrası enjeksiyon bakım
- Backfill yaklaşık 1 µL RNP enjeksiyon karışımı bir çekti Kılcal boru yukarıdakiler.
-
Sıkıştırılmış azot her embriyo microinject Rehm vd içinde açıklandığı gibi kullanmak 69.
- Parhyale embriyo bir microinjector ve bir micromanipulator kullanarak bir diseksiyon mikroskop altında enjekte. Enjeksiyon Mix 1,5 µL çekti kılcal tüp (4 inç - 1.0 mm ile filamentler, aparatı çekerek bir micropipette kullanarak çekti) geri yüklemek microloader pipet ucu kullanarak.
- Diseksiyon kapsamına forseps çifti kullanılarak iğne (çok küçük bir miktar) ucunu kırmak ve iğneyi Enjeksiyon cihazları üzerinde ayarlayın. 700 halocarbon yağın içine enjekte ve kabarcık çapı ölçme teslim birim kalibre.
- Bir tıraş bıçağı kullanarak kür ajan dışında bir 'yalak' kes. Yarıya kadar filtre sterilize deniz su ile doldurun ve Parhyale embriyo yalak stabilize etmek için sıraya girin.
- Her embriyo forseps bir çift ile enjeksiyon sırasında sabitleme mikroenjeksiyon kurulumu kullanarak embriyo enjekte. Enjeksiyon sonra cam transfer pipet embriyo üzerinde yarım filtre sterilize deniz suyu ile dolu bir taze 60 mm Kültür yemek aktarmak için kullanın.
-
Birinci Lig forma 2 hücreli embriyo (4-6 h sonrası döllenme) önceden olmuşsa, tamamen mutant hayvanlar her iki blastomeres enjekte edilerek oluşturmak. 2-hücre sahne alanı'nın toplam bir bölünme emin olmak için blastomeres işbirliği ile FITC veya TRITC dextran enjekte ve sinyal kapsamı sonra enjeksiyon anatomi bir floresan altında tek bir blastomere sınırlı olduğunu gözlemlemek.
- Alternatif olarak, 'yarım-mutant' hayvanlar sadece bir iki blastomeres (kabaca bölünmüş sol-sağ doku ve pozisyon A-P eksen boyunca bağlı olarak) 2-hücre aşamada enjekte edilerek oluşturmak.
- Tek hücre tek bir mikrop katman düzenleme kısıtlamak için bir 8 hücreli embriyo (7,5-9 h sonrası döllenme) olarak enjekte. Gerberding ve ark. görmek erken blastomere soy haritası için 70 .
-
60 mm Kültür yemekleri (en fazla 25 çanak başına), yarı dolu 'bir akvaryum bubbler kullanarak önceden oksijenli' filtre sterilize deniz suyu, ya da şiddetle sallayarak embriyo kuluçkaya.
- Embriyo yemekleri nem korumak ve onları 12 h koyu döngüsü ile 26 ° C kuluçka yer ıslak kağıt havlu ile kaplı bir gevşek mühürlü plasticware yerleştirin.
- Deniz suyu yemekleri birkaç günde temizlemek için hayatta kalan embriyolar transfer.
Not: her ne kadar onlar çok daha yavaş gelişecektir embriyo oda sıcaklığında kültürlü.
-
İncelemek ve embriyoların bir ifade analizi için çeşitli aşamalarını in situ hibridizasyon veya antikor boyama (bkz: tarafından Browne ve ark. düzeltmek 71 hazırlama kılavuzu ve diseksiyon ve fiksasyon72, in situ hibridizasyon73ve74boyama antikor için ek başvurular için).
- Tungsten tel bükülmüş bir parçası yaklaşık iş parçacığı tarafından diseksiyon iğneler yapmak bir insülin iğne ucunu uzunluğu 0.5. Sodyum hidroksit bir akım altında iğneyi keskinleştirmek. 1 mL şırınga diseksiyon iğne kolu kullanın.
- 3-şey cam yemeğin yarıya kadar bir şey bir taze yapılmış çözüm 9 parça PEM arabellek (0.1 M boru pH 6,95, EGTA, MgSO41 mM 2 mM), 1-Bölüm 10 x PBS ve 1 yarı % 32 ile doldurmak PFA. 3-5 embriyo çanak yerleştirin ve dışarı akmasını sarısı ve çalışması için sabitleştirici izin stabilize etmek, poke ve biraz dulled bir keskin tungsten iğne kullanarak her embriyo küçük bir delik poke.
- Bir çift bilenmiş tungsten iğne kullanarak, yavaşça uzakta Parhyale embriyo çevreleyen dış iki membran alay. Onları embriyolar daha güçlü olması için sabitleştirici teşrih ama hızlı bir şekilde membran membran kaldırma daha zor hale getirir embriyo için sabit olma tutmak için çalışmak. Embriyo antikor boyama için 15-20 dk. ya da in situ hibridizasyon için 40-50 dk toplam düzeltmek izin verir.
- Canlı yavru görüntü ve morfolojik ve davranışsal fenotipleri için çözümlemek veya düzeltmek ve onları daha detaylı analizler için leke. Nakavt ve transgenik çizgiler (bkz: Kontarakis ve Pavlopoulos75 yavru Bakımı ve diğer yararlı bilgi) kurmak için 2-3 ay içinde yavru cinsel olgunluk için kaldırın.
8. değerlendirme sonuçları düzenleme
- (Varsa), görsel ya da işlevsel fenotip düzenlenmiş hücreler veya organizmalar arayın.
Not: Bu işlem uygulama tarafından büyük ölçüde değişecektir ve onların ilgili Protokolü merdiven yukarıda sonunda bazı örnekler açıklanmaktadır. HSPCs orak hücre mutasyon düzelttikten sonra hemoglobin üretim HPLC (Şekil 1A) kullanarak farklı erythroblasts tarafından analiz. Bir nakavt IL-2 reseptör gen T hücrelerinde yüzey boyama tarafından onaylandı ve akış sitometresi (Şekil 1B). C. elegans ve P. hawaiensis fenotipleri değerlendirmek için hayvan morfolojisi ve davranış (rakamlar 1 c ve 1 D) bir ışık veya floresan mikroskop altında gözlemlemek. - Verimlilik ve oluşturulan genomik düzenlemeleri türünü belirlemek için düzenlenmiş hücreler takımlık parçalayıcı ve genomik DNA'ları bir ticari ekstraksiyon kiti21kullanarak ayıklayın.
-
Hızlı bir tahmini INDEL oluşumu için PCR-yükseltmek en az 200 baz çifti kesme etrafında site ve T7 endonuclease1 gerçekleştirmek (T7E1)76 veya arazi (CEL-1 nükleaz) tahlil77.
- Cas9-cut sitesi veya başarılı HDR INDEL oluşumu oluşturmak veya bilinen kısıtlama site kaldırma, Restriksiyon enzimi sindirim düzenleme verimliliği6tahmin etmek için kullanmayı düşünün. Kısıtlama parça Uzunluk Polimorfizmi (RFLP) tahlil kullanılabilir olması için olursa etkinliğini denetlemek için kullanışlı bir yol olabilir.
- Düzenleme verimliliği doğru bir miktar ve baskın düzenleme sonuçlar belirlenmesi için PCR amplicon için standart bir Sanger göndermek ileriye ve geriye doğru astar ile sıralama.
Not: bir tek klon veya organizma analiz, Sanger sonuçları analiz Şekil 2içindeAgösterildiği şekilde basittir. Sonra bir havuzu hücre analizi, chromatograms ile online araç78, Şekil 2' deBgösterildiği gibi analiz. - Tam bir miktar ve sonuçları düzenleme dizileri için derin sıralama27,54, Şekil 2' deCgösterildiği gibi gerçekleştirin.
- Hedef kapalı değişiklikleri, PCR-yükseltmek öngörülen hedef kapalı siteleri belirli bir dizi değerlendirmek ve onları NGS için göndermek için. Kromozomal translokasyonlar algılamasını etkinleştirmek için Kılavuzu-seq79 veya yüksek üretilen iş, genom-wide translocation sıralama (HTGTS)80gerçekleştirin. Hedef kapalı düzenlemeleri klonal bir popülasyondaki tam bir resim için bütün-genom sıralama (WGS)81,82,83gerçekleştirin.
Not: Orada on - ve off-hedef genom düzenlemeleri, açıkladı miktarının daha fazla makaleleri çeşitli İnceleme84,85,86için çeşitli yöntemler vardır.
Representative Results
Bu deneyler gösteri nasıl ön montajı Cas9 RNP primer hücre ve bütün organizmaların genleri işlemek için kullanılabilir. Araştırmacılar arındırmak veya Cas9 protein ve sgRNA satın almak, karmaşık önceden oluşturmak üzere iki parçaları birleştirmek ve kendi hücreleri veya organizma ilgi RNP tanıtmak. Oluşur ve (varsa) doğmak çoluk çocuk-in yeni nesil için fenotipleri için kontrol ve/veya hücreler Genotipleme için toplamak için düzenlemek için yeterli zaman izin sonra. Fenotipleri fonksiyonel deneyleri, ifade deneyleri, görselleştirme (göz veya mikroskobu ile) veya diğer yöntemine bağlı olarak deneme yoluyla gözlenen.
Örneğin, orak hücre hastalığı neden olur β globin mutasyon düzeltmek için düzenlenmiş HSPCs eritrositler farklılaşmış olabilir ve sağlıklı üretim veya orak hemoglobin27,87 (resim 1 için denetlesinler A). T hücreleri düzenlenmiş yüksek benzeşme IL-2 reseptör gen, CD25 yıkmak için (IL2RA), yüzey boyama ve akış sitometresi88tarafından analiz ve işlevsel olarak IL-2 stimülasyon (Şekil 1B sinyal bir yanıt bulmak için analiz ). T hücreleri de HIV enfeksiyonu89 etkinliği de dahil olmak üzere farklı fenotipleri değerlendirilmesi gereken birçok klinik olarak önemli yönden programlanabilir ve11 in vivo antitümör etkinliğini araba-T hücreleri.
Bir co-CRISPR/co-conversion tarama yaklaşım kullanarak, C. elegans solucanlar aynı anda iki loci62düzenlenmiş bulunmaktadır. HDR bir ssODN kullanarak dpy-10 referans gen, bir baskın dpy-10 kolayca attı işlevi kazanç mutasyon sonuçlarında şablon onarın. Heterozigoz F1dpy-10(gof) hayvan silindir (Rol) ve bodur (Dpy) homozigoz dpy-10(gof) hayvanlardır. Fenotip varlığı Cas9 düzenleme bu hayvanlara oluştu gösterir ve bir düzenleme olay Rol veya Dpy F1 hayvanlarda ikinci odağı olarak tanımlayan oran geliştirir. Başarılı bir düzenleme deneme Rol veya Dpy90olan 20 veya daha fazla F1 çoluk çocuk verimli enjekte P0 solucanlar % 33-50 sonuçlanmalıdır. O zaman Rol olmayan hayvanlar dpy-10 wildtype için dönmek ve faiz homozigoz düzenleme için seçmek için seçmek mümkündür. Pratik bir kural olarak, co-CRISPR başvuru gen hedefleme crRNA konsantrasyonu yarısı crRNA gen ilgi hedefleme olmalıdır. Bir düzenleme ilgi gen kurtarılır değil, iki CRISPR RNA'ların oranları istenen mutasyon kurtarma olasılığını artırmak için ayarlanabilir. Örneğin, crRNA başvuru gen crRNA göre faiz gen için miktarını artırarak düzenlemeleri de düzenlemeleri, referans gen locus sahip solucanlar nüfusu içinde ilgi gen sahip solucanlar yüzdesi artacaktır. Ortak dönüşüm frekanslar değişir, ancak genellikle % 20-60 kez F1 nesil (Şekil 1C) homozigoz düzenlemeleri verimli oranları.
P. Abdominal-B gen (Abd-B) yıkmak için düzenlenmiş hawaiensis yavru görüntülemek açık morfolojik anormallikler3 (resim 1D). Bu gen doğru karın desenlendirme için gereklidir ve torasik türü atlama ve bacaklar genellikle yüzme ve çapa bacakları yerine yürüyüş bozulma sonuçları karın bölgesinde mevcut.
Sonuçlar genotypic düzeyinde düzenleme genom belirleme sıralama veya sıra değişiklikleri algılar bir vitro tahlil gerekir. İşte, temsilcisi sıralama veri bizim model hücre türleri ve organizmalar, miktar düzenleme için farklı yaklaşımlar vurgulayarak göstermektedir. Çünkü tüm yöntemleri burada gösterilen herhangi bir biyolojik sisteme uygulanabilir Şekil etiketleri Genelleştirilmiş unutmayın.
Dayandırılmış yaklaşımlar teknik karmaşıklık ve sonuçları derinliği değişir. Klonal düzenlenen nüfus veya kolayca ayrılabilir bireysel organizmalar için düzenlenen bireyler genomik DNA ekstraksiyon aşağıdaki sıralı. Standart Sanger sıralama sonuçları (Şekil 2A) işlevini bozabilir varsayımsal frameshifts ile belirli bir birey Cas9 kesimli sitedeki sıra değişiminde ortaya çıkaracaktır. Sıralama için kullanılan online alet bireysel mutant78yerine karışık nüfus için uygulanabilecek başka bir Sanger-dayandırılmış bir yaklaşımdır. Sıraları genel düzenleme verimliliği hem de baskın sıra sonuçları yaklaşık bir çevrimiçi araç ile analiz edilir. Temsilcisi veri Şekil 2' deBgösterilir.
Burada açıklanan en kapsamlı sıralama yöntemi derin sıralama (bazen yüksek üretilen iş veya yeni nesil sıralama olarak da adlandırılır) olduğunu. Bu yöntem DNA dizileri nüfus içinde bireysel genleri üzerinden sağlar. Bu gibi veriler çeşitli şekillerde gösterildiği. Burada, düzenleme sonucu (Şekil 2C) göre düzenlenmiş hücrelerden bireysel sıralama okuma gizli. En hücreleri genellikle gene bozulma sonuçları NHEJ yolu üzerinden düzenlenmiş bulunmaktadır. Bazılarında, hedef gene dışarı HDR27ile alternatif sürümü için takas olmuştur.
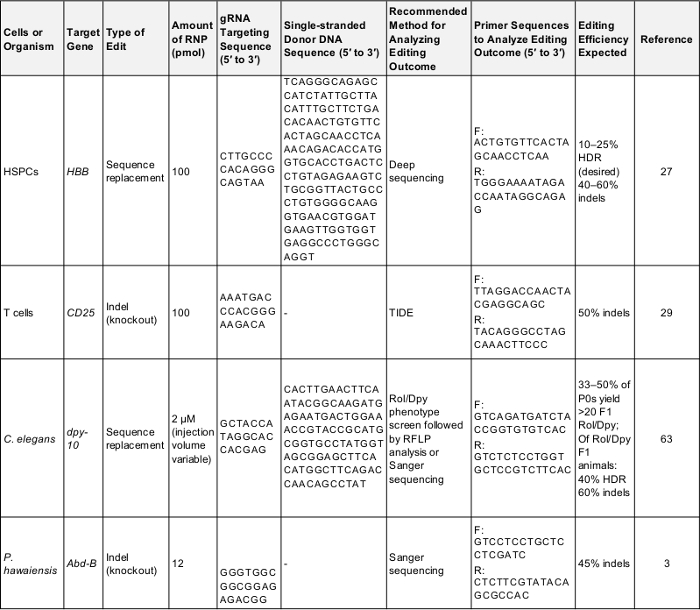
Tablo 1: pozitif kontrol deneyleri düzenleme ön genom için. Bu tabloda her hücre ve bu protokol için açıklanmıştır organizmaların Deneme düzenleme ilk genom gerçekleştirmek için gereken anahtar bilgileri gösterir. Bu parametreler aşağıdaki protokol sınamak için kullanılan başarılı bir sonuç vermeye olasıdır veya bir kez karşılaştırma için bir temel olarak deneyci bir gen kendi ilgi hedefliyor. F: ileri, R: ters, HDR: Homoloji-yönetmen onarım. Bu tabloyu indirmek için buraya tıklayınız.

Resim 1 : Temsilcisi fenotipik sonuç Cas9 RNP birincil insan hücreleri ve organizmaların düzenleme. (A) bu, başarılı genom Son aşama erythroblasts ayrılır HSPCs düzenleme, hemoglobin orak daha daha işlevsel hemoglobin ürettikten sonra gösterilen bir HPLC izidir. Başarılı bir şekilde düzenlenmiş hücreleri sağlıklı hemoglobin (HbA ve HbA2) fetal hemoglobin (HbF) yanı sıra üretecek iken mutant eritrositler orak hemoglobin (HbS), üretmek. Absorbans rasgele birimlerinde (au) Grafiği çizilecek. Bu panel ilk DeWitt et al. içinde yayınlandı 27. bilim ilerleme Amerikan Derneği'nden izni ile yayımlanmaktadır. (B) solda, her koşul için akış sitometresi veriler CD25 gen RNP ile nakavt sonra T hücre yüzey lekeli CD25 ifade yok olduğunu gösteren bu panel gösterir. CD25 bereket ile belgili tanımlık cep büyüklük y ekseni üzerinde x ekseni üzerinde çizilen. Her koşul için sağdaki Fosfo-Stat5 (pStat5) miktar bir indüksiyon ile IL-2 sonra bu panel gösterir. IL-2 reseptör (CD25 KO) olduğunda sinyal düşürülür. PStat5 bereket x ekseni üzerinde çizilen ve IL-2 giriş üç farklı düzeylerinden kaynaklanan veri dikey olarak karşılaştırılır. (C) Bu panel dpy-10 ortak dönüşüm marker olarak hedefleme bir Caenorhabditis elegans co-CRISPR/co-conversion ekran gösterir. İki kılavuz RNA'lar hedef iki loci, dpy-10 ve senin favori gen (yfg), içinde aynı P0-enjekte hayvan. HDR dpy - 10 bir Rol ya da Dpy fenotip sonuçlanır. Rol veya Dpy F1 hayvanlar yelpazesi düzenlemeleri ikinci odağı, tanımlama şansını artırır. (D) Bu panel wildtype Parhyale hawaiensis yavru yüzme ve çapa bacaklar ile normal abdomens olduğunu gösterir. Abd-B knock-out yavru (F0 bireyler) toraks doğru dönüştürülmüş bir karın geliştirmek. Böylece, yüzme ve çapa bacaklar gitti ve bir normal göğüs ile ilişkili atlama ve yürüme bacakları yerine. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.

Resim 2 : Sonuç analizi yöntemleri düzenlemesini tipik sonuçları. (A) Bu panel Sanger bireysel F1 P. hawaiensis organizmalar wildtype sırası ve açık okuma çerçevesi ilerletmeniz gen işlevini bozabilir üç farklı indels de dahil olmak üzere, sıralama sonuçlar gösterilmektedir. (B) Bu gelgit sonuçları göster çeşitli eklemeler ve sıralı T hücrelerinin bir havuzda bir Cas9-hedef sitede silme olayları. X ekseni verilen ekleme veya silme nükleotit uzunluğunu gösterir. (C) bu derin sıralama sonuçlar hiçbir genom nucleofection veya gRNA düzenleme göstermek ve başarılı olduğu gibi Cas9 RNP ile düzenleme, DNA onarım sonucu HSPCs. tarafından gruplandırılmış Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
Discussion
İletişim kuralı düzenleme sağlam genom kurulması bir hücrede satır veya organizma ilgi optimizasyonu gerektirir ve ampirik birkaç anahtar parametrelerinin test, bu bölümde açıklanan. Burada sunulan genel yaklaşımlar birkaç varyasyon çalışan son derece teşvik edilmektedir. Anahtar bu protokolü bu yöntemler diğer hücrelere başvuruda kısıtlamasıdır veya organizmalar okudu tür bağlı olarak farklı bir sonuç neden olabilir ve yüksek verimli gen nakavt yol açar bir deneysel tasarım DNA ekleme reklamını yapamaz. Böylece, biz burada sunulan ve aşağıda açıklandığı gibi sorun giderme yöntemleri ile başlayan öneririz.
Reaktif kalite düzenleme genom sorunlarını giderme
Oluşturmadan veya yüksek kaliteli Kimyasalları satın alma iletişim kuralını düzenleme herhangi bir genom kritik bir adımdır. Cas9 protein laboratuarda saf veya ticari olarak satın alınmış. Etkinlik düzenleme en iyi gen kaynağına bağlı olarak değişir bireysel herhangi bir Cas9 protein hazırlık özel faaliyetin bağlı olacaktır, ancak birçok iletişim kuralları son bir konsantrasyon Cas9 için RNP tariflerde unutmayın. Burada sunulan Protokolü çalışıyor sonra RNP en iyi bir konsantrasyon kurmak için titrating Cas9 düzeyleri tarafından kullanılan miktarını en iyi duruma getirme düşünün: neden gereksiz hedef kapalı bölünme olmadan son derece belirli hedef DNA bölünme sağlayan aşırı Cas940.
Kılavuzu RNA saflık ve homojenliği da başarı22düzenleme genom belirleyicileri olabilir. Satın alınan sgRNAs ya da ayrı crRNA ve tracrRNA bileşenleri genellikle yüksek kaliteli reaktifler ve kimyasal değişiklikler çeşitli RNA Bozulması sorunları ile mücadele için veya RNP91için ek özellikler aşılamak için kullanılabilir. Kimyasal olarak değiştirilmiş gRNAs standart genom deneyler düzenleme için gerekli olmayabilir iken, bazı gruplar daha yüksek verimliliği ile böyle reaktifler işleminin mastering sonra denemeye değer olabilir ve/veya zaman düzenleme gözlemledim gRNA bozulma bir sorunu22,91gibi görünüyor. Vitro transkripsiyon ve sonraki jel arıtma bir rutin genom deneyler17,21,49,50düzenleme için yeterli olabilir ucuz bir alternatif. Ayrıca, homojen gRNA nüfus içinde vivoribozyme ve tRNA tabanlı eksizyon bireysel kılavuzların da dahil olmak üzere, üretmek için yaygın olarak uygulanan, vitro için süpürge üretmek için RNA hazırlık uzatılabilir birkaç yaklaşım Ürünler92.
Tasarım İpuçları Kılavuzu RNA ve donör DNA:
Kılavuzu RNA seçimi son derece verimli hedef üzerinde hedef kapalı bölünme olasılığını en aza indirerek düzenleme ulaşmada önemli bir faktördür. Kılavuzu seçiminde yardımcı olmak için çeşitli çalışmalarda yeni nesil sıralama ile birleştiğinde yüksek üretilen iş ekranlar sıra özellikleri başarılı kılavuzları47,79,93,94derlemek için kullandık, 95,96. Bu özellikler akıllı algoritmalar ve Kılavuzu seçimi44,45,46,47,48yılında yardımcı olan çevrimiçi araçlar geliştirmek için kullanılmaktadır. Böyle algoritmalar DNA tabanlı sistemler kullanma kılavuzu için RNA ifade ekranlarda cezalısın. Kılavuzları Pol III organizatörü kullanılarak ifade edilir ve bu nedenle onların ifade urasil97,98izleri karşılaşıldığında erken sonlandırma gibi Pol III transkripsiyon ile ilgili sınırlamalar eğilimli, 99. Ancak, RNPs kullanımı ile vitrogetirdi-sentezlenmiş Kılavuzu RNA'ların endişeleri atlar ve Kılavuzu tasarım kısıtlamalar kolaylaştırır. Bu algoritmalar kadar ortaya çıkan ve son derece etkili genom düzenleme ile çok sayıda çalışma onaylamıştır bir ortak özelliğidir bir pürin, özellikle bir guanin, kılavuz hedef özgü sıra 3'ucunda varlığı. Bu kılavuz özelliği memeliler C. elegans, meyve sinekleri ve zebra balığı65,100,101kadar organizmalar arasında çok başarılı olmuştur. Buna ek olarak, C. elegansiçin kılavuz hedefleme bölge 3'ucunda bir GG dinükleotit kılavuzlarıyla tasarımı son derece etkili Kılavuzu RNA'ların65tahmin için etkili bir strateji olduğunu. İdeal olarak, birden çok kılavuzları paralel hangisinin belirli bir uygulama için en başarılı olduğunu belirlemek için sınayın.
Bir DNA dizisi genom tanıtmak çalışılırken, donör veya şablon DNA tasarımını da çok önemlidir. Tek iplikçikli oligonükleotid bağış (ssODNs) diğer tipik onarım şablonları, doğrusal çift iplikçikli ve plazmid DNA54,55,102daha güvenilir bir şekilde eklenir. Bazı loci sigara hedefe tamamlayıcı veya DNA dizisi yerlerinden ve uzunluğu27,55asimetrik Homoloji silah sahip ssODNs ile HDR verimliliği artırılabilir. Onarım şablon kesim sitesinde eklenir ve hedeflenen sırası içerir beri donör DNA önce veya sonra genomik ekleme bulandı Cas9 engellemek için adımların atılması gerekir. Bu sessiz mutasyonlar PAM sıra veya tohum bölge, tanıma Cas9 tarafından eklenen gen21,103fonksiyonun ise istinat kaçınarak yaparak gerçekleştirilir. PAM'yi bile tek nükleotid değişiklikler bağlama104kaldırılması olasılığı olmasına rağmen güvenli olması için en az dört nükleotit değiştirmeye çalışın.
Önemi ve gelecekteki uygulamalar:
CRISPR-Cas9 ile düzenleme genom facile genetik manipülasyon herhangi bir organizmanın sağlayan güçlü bir yöntem ortaya çıkmıştır. Cas9 RNP ile düzenleme ilk başta biraz daha fazla çaba alır ama reaktifler ve protokolleri bir laboratuarda oluşturulduktan sonra kullanmaya devam edebilirsiniz basittir. Daha az hedef kapalı etkileri24,25,26 ile HDR, üzerinden ulaşmak zor gen ekleme de dahil olmak üzere daha yüksek genel düzenleme verimliliği ön montajı RNP plazmid DNA yerine hücrelerle düzenleme yol açar , 27 , 29. Ayrıca, Denemecileri gen ekspresyonu, RNA bozulması, protein katlanması ve gRNA ayrı olarak sentezlenmiş içinde hücre22,23Cas9 moleküller arasındaki ilişkiyi sorunlardan kaçınmak. RNP düzenleme de güvenlik kaygıları hakkında insertional mutagenesis kaçınmanızı sağlar ve viral teslim yöntemleri olduğunda ortaya çıkabilecek sürekli ifade klinik olarak14kullandık. Bu avantajlar nedeniyle pek çok bilim adamı önceden klinik iletken RNP düzenleme insan tedavi uygulamaları için prototip deneyler lehine. Tedavi etmek ya da bile tedavi koşulları, Duchenne kas distrofisi105 ve orak hücre hastalığı27 gibi genetik hastalıklardan çeşitli geliştirme vivo içinde ve ex vivo genom RNP tabanlı düzenleme yaklaşımlar vardır HIV1129 ve kanser. İlginçtir, Cas9 RNP giderek 'DNA-ücretsiz' bitkiler33,34,36toplam düzenleme sağladığından tarımsal Mühendislik için Teslimat yöntemi olarak istihdam edilmektedir.
Disclosures
Yazar Alexander Marson ve Jacob E. Mısır kurucu spot tedavi vardır. Jacob E. Mısır misyon Therapeutics danışmanı ve AstraZeneca ve Pfizer onun laboratuvar araştırma projeleri destek aldı. Alexander Marson Juno Therapeutics ve Paktı Therapeutics danışmanı olduğunu ve onun laboratuvar araştırma projeleri destek Juno Therapeutics, Epinomics ve Sanofi aldı. Onun laboratuvarı da Cas9 RNP teknoloji ile ilgili patentler için başvurdu.
Acknowledgments
Birçok önceki üyeleri bizim labs ve Bay Area genom düzenleme topluluk bu yöntemler gelişmesine katkılarından dolayı teşekkür ediyoruz. Ross Wilson eleştirel Bu makale okumak için teşekkür ederiz.
Alexander Marson'ın araştırma Jake Aronov bir hediye tarafından desteklenmektedir ve ulusal bir multipl skleroz Derneği (CA 1074-A-21) verin. Alexander Marson Burroughs Wellcome fondan tıp bilim adamları için bir kariyer Ödülü tutar ve bir Chan Zuckerberg Biohub Dedektif. Jacob E. Corn'ın araştırma için rejeneratif tıp Li Ka Shing Vakfı, miras tıbbi araştırma Tıp Enstitüsü ve California Institute tarafından desteklenir. Behnom Farboud ve Barbara J. Meyer araştırma kısmen Barbara J. Meyer, Howard Hughes Tıp Enstitüsü'nün bir dedektif için NIGMS grant R01 GM030702 tarafından finanse edilmektedir. Erin Jarvis ve Nipam H. Patel'ın araştırma kısmen NSF grant IOS-1257379 tarafından finanse edilmektedir ve Erin Jarvis bir NSF GRFP ve Philomathia yüksek lisans bursu destek kabul eder.
Materials
| Name | Company | Catalog Number | Comments |
| Reagents/Materials | |||
| DNA oligonucleotides | Integrated DNA Technologies | - | IDT will provide custom DNA sequences, including those in Table 1 |
| Guide RNAs | Synthego | - | Synthego will provide high-quality sgRNAs for S. pyogenes Cas9, including custom sgRNAs containing the targeting sequences included in Table 1 |
| Purified Cas9 protein (EnGen Cas9 NLS, S. pyogenes) | New England Biosciences | M0646T | If possible, purifying Cas9 in-house or purchasing from local core facilities is a less expensive option |
| Normal peripheral blood CD34+ stem/progenitor cells | AllCells | PB032-2 | |
| StemSpan SFEM | StemCell Technologies | 09650 | |
| StemSpan CC110 | StemCell Technologies | 02697 | |
| P3 Primary Cell 4D-Nucleofector X Kit | Lonza | V4XP-3032 | |
| RPMI-1640 Medium, With sodium bicarbonate, without L-glutamine, liquid | Sigma | R0883-6X500ML | |
| EasySep™ Human T Cell Isolation Kit | Stemcell | 17951 | |
| cell culture plate, 96 wells, round | Fisher Scientific | 3799 | |
| CTS (Cell Therapy Systems) Dynabeads CD3/CD28 | Life Tech | 40203D | |
| Reombinant Human IL-2 | UCSF Pharmacy | NA | |
| SepMate-50 500-pack IVD | Stemcell Technologies | 85460 | |
| OP50 Escherichia coli | Caenorhabditis Genetics Center | OP-50 | https://cgc.umn.edu/ |
| Nematode Growth Media agar in petri dishes | - | - | See Stiernagle, T (ref. 59) |
| Standard borosilicate glass capillaries with filament: 4 in (100 mm), 1/0.58 OD/ID | World Precision Instruments | 1B100F-4 | |
| Single-barrel standard borosilicate glass capillaries: 6 in (152 mm), 2/1.12 OD/ID | World Precision Instruments | 1B200-6 | |
| Cover glass; 24 × 50 mm | Thermo Fisher Scientific | 12-544E | |
| Cover glass; 22 × 22 mm | Thermo Fisher Scientific | 12-518-105K | |
| Apex LE agarose | Genesee Scientific | 20-102 | |
| Halocarbon oil 700 | Sigma-Aldrich | H8898-100ML | |
| pCFJ90 plasmid | Addgene | 19327 | |
| Compressed nitrogen | - | ||
| 60 mM culture dishes | BD | ||
| Capillary tubes with filament: 4 in (1.0 mm) | World Precision Instruments | T2100F-4 | |
| Sylgard 184 | Dow Corning | ||
| Petri dishes (100 × 15 mm) | - | ||
| Tungsten wire (0.005 in. diameter) | Ted Pella | ||
| Perfluoroalkoxy alkane (PFA) | - | ||
| Marine salt | - | ||
| 9" pasteur pipettes | - | ||
| Phenol red | - | ||
| Nuclease-free water | - | ||
| Equipment | |||
| 4D Nucleofector | Lonza | AAF-1002X | |
| MZ75 Stereomicroscope | Leica | Out-of-production. Current model is the M80 Stereomicroscope | |
| Axio Vert35 inverted phase contrast fluorescent microscope | Zeiss | Out-of-production. Current model is the Axio VertA.1 | |
| Laser-based micropipette puller (for C. elegans protocol) | Sutter Instrument | FG-P2000 | |
| Picoliter Microinjector (for C. elegans protocol) | Warner Instruments | PLI-100A | |
| Three-axis Joystick oil hydraulic micromanipulator | Narishige International | MO-202U | |
| Coarse manipulator | Narishige International | MMN-1 | |
| Micropipette puller (for P. hawaiensis protocol) | Sutter Instrument | P-80/PC | |
| Microinjector (for P. hawaiensis protocol) | Narishige | IM300 | |
| Microloader pipette tips | Eppendorf | 5242956003 | |
| NG-agar |
References
- Komor, A. C., Badran, A. H., Liu, D. R. CRISPR-based technologies for the manipulation of eukaryotic genomes. Cell. , 1-17 (2016).
- Barrangou, R., Horvath, P. A decade of discovery: CRISPR functions and applications. Nature Microbiology. 2, 1-9 (2017).
- Martin, A., Serano, J. M., et al. CRISPR/Cas9 mutagenesis reveals versatile roles of Hox genes in crustacean limb specification and evolution. Current Biology. 26 (1), 14-26 (2016).
- Goldstein, B., King, N. The future of cell biology: emerging model organisms. Trends in Cell Biology. 26 (11), 818-824 (2016).
- Mali, P., Yang, L., et al. RNA-guided human genome engineering via Cas9. Science. 339 (6121), 823-826 (2013).
- Cong, L., Ran, F. A., et al. Multiplex genome engineering using CRISPR/Cas systems. Science. 339 (6121), 819-823 (2013).
- Deltcheva, E., Chylinski, K., et al. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature. 471 (7340), 602-607 (2011).
- Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., Charpentier, E. A Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 337 (6096), 816-821 (2012).
- Nowak, C. M., Lawson, S., Zerez, M., Bleris, L. Guide RNA engineering for versatile Cas9 functionality. Nucleic Acids Research. 44 (20), 9555-9564 (2016).
- Jiang, W., Cox, D., Zhang, F., Bikard, D., Marraffini, L. A. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nature Biotechnology. , 1-9 (2013).
- Rupp, L. J., Schumann, K., et al. CRISPR/Cas9-mediated PD-1 disruption enhances anti-tumor efficacy of human chimeric antigen receptor T cells. Scientific Reports. 7 (1), 737 (2017).
- Graham, D. B., Root, D. E. Resources for the design of CRISPR gene editing experiments. Genome Biology. 16, 260 (2015).
- Wang, H., La Russa, M., Qi, L. S. CRISPR/Cas9 in genome editing and beyond. Annual Review of Biochemistry. 85, 2270 (2016).
- Nelson, C. E., Gersbach, C. A. Engineering delivery vehicles for genome editing. Annual Review of Chemical and Biomolecular Engineering. 7, 637-662 (2016).
- Yin, H., Kauffman, K. J., Anderson, D. G. Delivery technologies for genome editing. Nature Reviews Drug Discovery. 16 (6), 387-399 (2017).
- Zuris, J. A., Thompson, D. B., et al. Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo. Nature Biotechnology. 33 (1), 73-80 (2015).
- Cho, S. W., Lee, J., Carroll, D., Kim, J. -S., Lee, J. Heritable gene knockout in Caenorhabditis elegans by direct injection of Cas9-sgRNA ribonucleoproteins. Genetics. 195 (3), 1177-1180 (2013).
- Wang, W., Kutny, P. M., et al. Delivery of Cas9 protein into mouse zygotes through a series of electroporation dramatically increases the efficiency of model creation. Journal of Genetics and Genomics. 43 (5), 319-327 (2016).
- Chen, S., Lee, B., Lee, A. Y. -F., Modzelewski, A. J., He, L. Highly efficient mouse genome editing by CRISPR ribonucleoprotein electroporation of zygotes. Journal of Biological Chemistry. 291 (28), 14457-14467 (2016).
- Remy, S., Chenouard, V., et al. Generation of gene-edited rats by delivery of CRISPR/Cas9 protein and donor DNA into intact zygotes using electroporation. Scientific Reports. 7 (1), 16554 (2017).
- DeWitt, M. A., Corn, J. E., Carroll, D. Genome editing via delivery of Cas9 ribonucleoprotein. Methods. , 1-7 (2017).
- Hendel, A., Bak, R. O., et al. Chemically modified guide RNAs enhance CRISPR-Cas genome editing in human primary cells. Nature Biotechnology. 33 (9), 985-989 (2015).
- Thyme, S. B., Akhmetova, L., Montague, T. G., Valen, E., Schier, A. F. Internal guide RNA interactions interfere with Cas9-mediated cleavage. Nature Communications. 7, 11750 (2016).
- Kim, S., Kim, D., Cho, S. W., Kim, J., Kim, J. -S. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Research. 24 (6), 1012-1019 (2014).
- Lin, S., Staahl, B. T., Alla, R. K., Doudna, J. A. Enhanced homology-directed human genome engineering by controlled timing of CRISPR/Cas9 delivery. eLife. 3, 04766 (2014).
- Liang, X., Potter, J., et al. Rapid and highly efficient mammalian cell engineering via Cas9 protein transfection. Journal of Biotechnology. 208, 44-53 (2015).
- DeWitt, M. A., Magis, W., Bray, N. L., Wang, T. Selection-free genome editing of the sickle mutation in human adult hematopoietic stem/progenitor cells. Science Translational Medicine. 8 (360), (2016).
- Ramakrishna, S., Kwaku Dad, A. -B., Beloor, J., Gopalappa, R., Lee, S. -K., Kim, H. Gene disruption by cell-penetrating peptide-mediated delivery of Cas9 protein and guide RNA. Genome Research. 24 (6), 1020-1027 (2014).
- Schumann, K., Lin, S., et al. Generation of knock-in primary human T cells using Cas9 ribonucleoproteins. Proceedings of the National Academy of Sciences of the United States of America. 112 (33), 10437-10442 (2015).
- Lee, J. -S., Kwak, S. -J., et al. RNA-guided genome editing in Drosophila with the purified Cas9 protein. G3: Genes, Genomes, Genetics (Bethesda, MD). 4 (7), 1291-1295 (2014).
- Sung, Y. H., Kim, J. M., et al. Highly efficient gene knockout in mice and zebrafish with RNA-guided endonucleases. Genome Research. 24 (1), 125-131 (2014).
- Menoret, S., De Cian, A., et al. Homology-directed repair in rodent zygotes using Cas9 and TALEN engineered proteins. Scientific Reports. 5, 14410 (2015).
- Woo, J. W., Kim, J., et al. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nature Biotechnology. 33 (11), 1162-1164 (2015).
- Malnoy, M., Viola, R., et al. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Frontiers in Plant Science. 7, 1904 (2016).
- Svitashev, S., Schwartz, C., Lenderts, B., Young, J. K., Mark Cigan, A. Genome editing in maize directed by CRISPR-Cas9 ribonucleoprotein complexes. Nature Communications. 7, 13274 (2016).
- Liang, Z., Chen, K., et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nature Communications. 8, 14261 (2017).
- Shin, S. -E., Lim, J. -M., et al. CRISPR/Cas9-induced knockout and knock-in mutations in Chlamydomonas reinhardtii. Scientific Reports. 6, 27810 (2016).
- Pohl, C., Kiel, J. A. K. W., Driessen, A. J. M., Bovenberg, R. A. L., Nygård, Y. CRISPR/Cas9 based genome editing of Penicillium chrysogenum. ACS Synthetic Biology. 5 (7), 754-764 (2016).
- Grahl, N., Demers, E. G., Crocker, A. W., Hogan, D. A. Use of RNA-protein complexes for genome editing in non-albicans Candida species. mSphere. 2 (3), (2017).
- Rivera-Torres, N., Kmiec, E. B. A standard methodology to examine on-site mutagenicity as a function of point mutation repair catalyzed by CRISPR/Cas9 and ssODN in human cells. Journal of Visualized Experiments. (126), (2017).
- Nandal, A., Mallon, B., Telugu, B. P. Efficient generation and editing of feeder-free IPSCs from human pancreatic cells using the CRISPR-Cas9 system. Journal of Visualized Experiments. (129), (2017).
- Mohr, S. E., Hu, Y., Ewen-Campen, B., Housden, B. E., Viswanatha, R., Perrimon, N. CRISPR guide RNA design for research applications. The FEBS Journal. 283 (17), 3232-3238 (2016).
- Bauer, D. E., Canver, M. C., Orkin, S. H. Generation of genomic deletions in mammalian cell lines via CRISPR/Cas9. Journal of Visualized Experiments. (95), e52118 (2015).
- Hsu, P. D., Scott, D. A., et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nature Biotechnology. 31 (9), 827-832 (2013).
- Heigwer, F., Kerr, G., Boutros, M. E-CRISP: fast CRISPR target site identification. Nature Methods. 11 (2), 122-123 (2014).
- Moreno-Mateos, M. A., Vejnar, C. E., et al. CRISPRscan: designing highly efficient sgRNAs for CRISPR-Cas9 targeting in vivo. Nature Methods. 12 (10), 982-988 (2015).
- Labun, K., Montague, T. G., Gagnon, J. A., Thyme, S. B., Valen, E. CHOPCHOP v2: a web tool for the next generation of CRISPR genome engineering. Nucleic Acids Research. 44, 272-276 (2016).
- Haeussler, M., Schönig, K., et al. Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome Biology. 17 (1), 148 (2016).
- Lo, T. -W., Pickle, C. S., et al. Precise and heritable genome editing in evolutionarily diverse nematodes using TALENs and CRISPR/Cas9 to engineer insertions and deletions. Genetics. 195 (2), 331-348 (2013).
- Bassett, A., Liu, J. -L. CRISPR/Cas9 mediated genome engineering in Drosophila. Methods. 69 (2), 128-136 (2014).
- Prior, H., Jawad, A. K., MacConnachie, L., Beg, A. A. Highly efficient, rapid and co-CRISPR independent genome editing in Caenorhabditis elegans. G3: Genes, Genomes, Genetics. , Bethesda, MD. (2017).
- Hirsh, A. Cas9 expression and purification protocol. protocols.io. , (2017).
- DeWitt, M. A., Wong, J. In vitro transcription of guide RNAs. protocols.io. , (2017).
- Yang, L., Guell, M., et al. Optimization of scarless human stem cell genome editing. Nucleic Acids Research. 41 (19), 9049-9061 (2013).
- Richardson, C. D., Ray, G. J., DeWitt, M. A., Curie, G. L., Corn, J. E. Enhancing homology-directed genome editing by catalytically active and inactive CRISPR-Cas9 using asymmetric donor DNA. Nature Biotechnology. 34 (3), 339-344 (2016).
- Paix, A., Folkmann, A., Seydoux, G. Precision genome editing using CRISPR-Cas9 and linear repair templates in C. elegans. Methods. 121-122, 86-93 (2017).
- Mello, C., Fire, A. DNA transformation. Methods in Cell Biology. 48, 451-482 (1995).
- Sutter Pipette Cookbook. , Available from: https://www.sutter.com/PDFs/pipette_cookbook.pdf (2017).
- Stiernagle, T. Maintenance of C. elegans. WormBook: the online review of C. elegans biology. , (2006).
- Evans, T. C. Transformation and microinjection. WormBook: the online review of C. elegans biology. , (2006).
- Berkowitz, L. A., Knight, A. L., Caldwell, G. A., Caldwell, K. A. Generation of stable transgenic C. elegans using microinjection. Journal of Visualized Experiments. (18), (2008).
- Kim, H., Ishidate, T., et al. A co-CRISPR strategy for efficient genome editing in Caenorhabditis elegans. Genetics. 197 (4), 1069-1080 (2014).
- Arribere, J. A., Bell, R. T., Fu, B. X. H., Artiles, K. L., Hartman, P. S., Fire, A. Z. Efficient marker-free recovery of custom genetic modifications with CRISPR/Cas9 in Caenorhabditis elegans. Genetics. 198 (3), 837-846 (2014).
- Ward, J. D. Rapid and precise engineering of the Caenorhabditis elegans genome with lethal mutation co-conversion and inactivation of NHEJ repair. Genetics. 199 (2), 363-377 (2015).
- Farboud, B., Meyer, B. J. Dramatic enhancement of genome editing by CRISPR/Cas9 through improved guide RNA design. Genetics. 199 (4), 959-971 (2015).
- Wood, A. J., Lo, T. -W., et al. Targeted genome editing across species using ZFNs and TALENs. Science. 333 (6040), 307 (2011).
- Friedland, A. E., Tzur, Y. B., Esvelt, K. M., Colaiácovo, M. P., Church, G. M., Calarco, J. A. Heritable genome editing in C. elegans via a CRISPR-Cas9 system. Nature Methods. 10 (8), 741-743 (2013).
- Dickinson, D. J., Ward, J. D., Reiner, D. J., Goldstein, B. Engineering the Caenorhabditis elegans genome using Cas9-triggered homologous recombination. Nature Methods. 10 (10), 1028-1034 (2013).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. Injection of Parhyale hawaiensis blastomeres with fluorescently labeled tracers. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Gerberding, M., Browne, W. E., Patel, N. H. Cell lineage analysis of the amphipod crustacean Parhyale hawaiensis reveals an early restriction of cell fates. Development (Cambridge, England). 129 (24), 5789-5801 (2002).
- Browne, W. E., Price, A. L., Gerberding, M., Patel, N. H. Stages of embryonic development in the amphipod crustacean, Parhyale hawaiensis. Genesis. 42 (3), New York, NY. 124-149 (2005).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. Fixation and dissection of Parhyale hawaiensis embryos. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. In situ hybridization of labeled RNA probes to fixed Parhyale hawaiensis embryos. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Rehm, E. J., Hannibal, R. L., Chaw, R. C., Vargas-Vila, M. A., Patel, N. H. Antibody staining of Parhyale hawaiensis embryos. Cold Spring Harbor Protocols. 2009 (1), (2009).
- Kontarakis, Z., Pavlopoulos, A. Transgenesis in non-model organisms: the case of Parhyale. Methods in Molecular Biology. 1196, Clifton, NJ. 145-181 (2014).
- Kim, H. J., Lee, H. J., Kim, H., Cho, S. W., Kim, J. -S. Targeted genome editing in human cells with zinc finger nucleases constructed via modular assembly. Genome Research. 19 (7), 1279-1288 (2009).
- Qiu, P., Shandilya, H., D'Alessio, J. M., O'Connor, K., Durocher, J., Gerard, G. F. Mutation detection using Surveyor nuclease. BioTechniques. 36 (4), 702-707 (2004).
- Brinkman, E. K., Chen, T., Amendola, M., van Steensel, B. Easy quantitative assessment of genome editing by sequence trace decomposition. Nucleic Acids Research. 42 (22), 168 (2014).
- Tsai, S. Q., Zheng, Z., et al. GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases. Nature Biotechnology. 33 (2), 187-197 (2015).
- Frock, R. L., Hu, J., Meyers, R. M., Ho, Y. -J., Kii, E., Alt, F. W. Genome-wide detection of DNA double-stranded breaks induced by engineered nucleases. Nature Biotechnology. 33 (2), 179-186 (2015).
- Smith, C., Gore, A., et al. Whole-genome sequencing analysis reveals high specificity of CRISPR/Cas9 and TALEN-based genome editing in human iPSCs. Cell Stem Cell. 15 (1), 12-13 (2014).
- Veres, A., Gosis, B. S., et al. Low incidence of off-target mutations in individual CRISPR-Cas9 and TALEN targeted human stem cell clones detected by whole-genome sequencing. Cell Stem Cell. 15 (1), 27-30 (2014).
- Kim, D., Bae, S., et al. Digenome-seq: genome-wide profiling of CRISPR-Cas9 off-target effects in human cells. Nature Methods. 12 (3), 237-243 (2015).
- Hendel, A., Fine, E. J., Bao, G., Porteus, M. H. Quantifying on- and off-target genome editing. Trends in Biotechnology. 33 (2), 132-140 (2015).
- O'Geen, H., Yu, A. S., Segal, D. J. How specific is CRISPR/Cas9 really. Current Opinion in Chemical Biology. 29, 72-78 (2015).
- Tsai, S. Q., Joung, J. K. Defining and improving the genome-wide specificities of CRISPR-Cas9 nucleases. Nature Reviews Genetics. 17 (5), 300-312 (2016).
- Hoban, M. D., Cost, G. J., et al. Correction of the sickle cell disease mutation in human hematopoietic stem/progenitor cells. Blood. 125 (17), 2597-2604 (2015).
- Simeonov, D. R., Gowen, B. G., et al. Discovery of stimulation-responsive immune enhancers with CRISPR activation. Nature. , (2017).
- Hultquist, J. F., Schumann, K., et al. A Cas9 ribonucleoprotein platform for functional genetic studies of HIV-host interactions in primary human T cells. Cell Reports. 17 (5), 1438-1452 (2016).
- Paix, A., Wang, Y., et al. Scalable and versatile genome editing using linear DNAs with microhomology to Cas9 sites in Caenorhabditis elegans. Genetics. 198 (4), 1347-1356 (2014).
- Lee, K., Mackley, V. A., et al. Synthetically modified guide RNA and donor DNA are a versatile platform for CRISPR-Cas9 engineering. eLife. 6, (2017).
- Minkenberg, B., Wheatley, M., Yang, Y. CRISPR/Cas9-enabled multiplex genome editing and its application. Progress in Molecular Biology and Translational Science. 149, 111-132 (2017).
- Doench, J. G., Fusi, N., et al. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nature Biotechnology. 34 (2), 184-191 (2016).
- Doench, J. G., Hartenian, E., et al. Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation. Nature Biotechnology. , 1-8 (2014).
- Liu, H., Wei, Z., Dominguez, A., Li, Y., Wang, X., Qi, L. S. CRISPR-ERA: a comprehensive design tool for CRISPR-mediated gene editing, repression and activation. Bioinformatics (Oxford, England). 31 (22), 3676-3678 (2015).
- Wu, X., Scott, D. A., et al. Genome-wide binding of the CRISPR endonuclease Cas9 in mammalian cells. Nature Biotechnology. 32 (7), 670-676 (2014).
- Bogenhagen, D. F., Brown, D. D. Nucleotide sequences in Xenopus 5S DNA required for transcription termination. Cell. 24 (1), 261-270 (1981).
- Cozzarelli, N. R., Gerrard, S. P., Schlissel, M., Brown, D. D., Bogenhagen, D. F. Purified RNA polymerase III accurately and efficiently terminates transcription of 5S RNA genes. Cell. 34 (3), 829-835 (1983).
- Chen, B., Gilbert, L. A., et al. Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas system. Cell. 155 (7), 1479-1491 (2013).
- Gagnon, J. A., Valen, E., et al. Efficient mutagenesis by Cas9 protein-mediated oligonucleotide insertion and large-scale assessment of single-guide RNAs. PLoS ONE. 9 (5), 98186 (2014).
- Ren, X., Yang, Z., et al. Enhanced specificity and efficiency of the CRISPR/Cas9 system with optimized sgRNA parameters in Drosophila. Cell Reports. 9 (3), 1151-1162 (2014).
- Ran, F. A., Hsu, P. D., Wright, J., Agarwala, V., Scott, D. A., Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nature Protocols. 8 (11), 2281-2308 (2013).
- Serano, J. M., Martin, A., et al. Comprehensive analysis of Hox gene expression in the amphipod crustacean Parhyale hawaiensis. Developmental Biology. 409 (1), 297-309 (2016).
- Sternberg, S. H., Redding, S., Jinek, M., Greene, E. C., Doudna, J. A. DNA interrogation by the CRISPR RNA-guided endonuclease Cas9. Nature. , 1-17 (2014).
- Lee, K., Conboy, M., et al. Nanoparticle delivery of Cas9 ribonucleoprotein and donor DNA in vivo induces homology-directed DNA repair. Nature Biomedical Engineering. 1 (11), 889-901 (2017).
