

Summary
Die zytotoxische Aktivität der Phenanthridin PJ-34 in Krebszellen Mitose wurde in Echtzeit Live-konfokale Bildgebung dokumentiert. PJ-34 ausgerottet menschlichen Brustkrebszellen MDA-MB-231-Zellen, die extra Centrosomen in Mitose. Im Gegensatz zu normalen bifokalen Mitose wurden die extra Zentrosome in den beiden Spindelpole in Gegenwart von PJ-34 gruppiert.
Abstract
Phenanthrenderivate als potente Inhibitoren PARP1 verhinderte die bi-fokale Clustering überzähliger Centrosomen in multi-zentrosomale menschliche Krebszellen in der Mitose. Die Phenanthridin PJ-34 war die stärkste Molekül. Declustering der extra-Zentrosome verursacht mitotischen Versagen und Zelltod in multi-zentrosomale Zellen. Die meisten soliden menschlichen Krebserkrankungen eine hohe Auftreten von extra Zentrosome. Die Aktivität der PJ-34 wurde in Echtzeit durch konfokale Bildgebung von lebenden menschlichen Brustkrebszellen MDA-MB-231-Zellen mit Vektoren, die für fluoreszierende γ-Tubulin, die sehr häufig in den Centrosomen ist und für Leuchtstofflampen Histone H2B in transfiziert dokumentiert die Chromosomen. Aberrante Chromosom Anordnungen und de-Cluster γ-Tubulin Brennpunkte darstellt Declustered Zentrosome wurden in den transfizierten MDA-MB-231-Zellen nach der Behandlung mit PJ-34 erfasst. Un-Cluster extra Centrosomen in den beiden vorangegangen Spindelpole ihren Zelltod. Diese Ergebnisse verknüpft for das erste Mal, das kürzlich entdeckt exklusiven zytotoxische Aktivität von PJ-34 in menschlichen Krebszellen mit extra Zentrosome de-Clustering in der Mitose und mitotischen Ausfall zum Zelltod führen. Nach den bisherigen Erkenntnissen durch konfokale Bildgebung von fixierten Zellen, PJ-34 ausschließlich ausgerottet Krebszellen mit Multi-Zentrosome ohne Beeinträchtigung normalen Zellen Mitose mit zwei Centrosomen und bi-fokale Spindeln beobachtet. Dieser zytotoxische Aktivität von PJ-34 wurde nicht durch andere potente Inhibitoren PARP1 geteilt, und wurde in PARP1 defizienten MEF beherbergen extracentrosomes beobachtet, was seine Unabhängigkeit von PARP1 Hemmung. Live-konfokale Bildgebung bot ein nützliches Werkzeug für die Identifizierung neuer Moleküle beseitigen Zellen während der Mitose.
Introduction
Phenanthrene abgeleitet PARP1 Hemmern, einschließlich PJ-34, wurden entwickelt, um ruhende Zellen von Apoptose durch die Energie verbrauchen PARP1 vermittelten DNA-Reparatur unter Stressbedingungen (Schlaganfall oder Myokardinfarkt) 1 induziert schützen. Doch vor kurzem entdeckten wir, dass PJ-34, bei zweimal höhere Konzentration als die induzierende PARP1 Hemmung kann ausschließlich Zelltod verursachen in menschlichen Krebszellen 2,3. Je schneller die Proliferation der Zelle war, desto effizienter ist die Tilgung der Zellen. Die zytotoxische Aktivität von PJ-34 wurde extra Zentrosome de-Clustering in Mitose 2 zugeschrieben. Viele menschliche Krebszellen Hafen multicentrosomes 4,5. Inkubation von menschlichen Brustkrebszellen MDA-MB-231, die überzähligen Centrosomen Hafen, mit 20 uM PJ-34 effizient beseitigt diese Zellen innerhalb 72-96 Stunden ohne Beeinträchtigung ruhenden Zellen oder einige gutartige proliferierende Zellen beherbergen zwei Centrosomen in der Mitose
Bipolar Zentrosoms Montage ist entscheidend für bipolare Spindel Bildung in der Mitose 4,5. Daher wurden Zellen mit mehr als zwei Centrosomen eine kaum verstanden molekularen Mechanismus entwickelt, Clustering ihre zusätzliche Zentrosome an zwei Pole 4-9. Ausfall der bipolaren Anordnung ihrer Zentrosome kann multipolare Spindeln und aberrante verzerrten Chromosomen Segregation, dass die Verhaftungen Zellzyklus in der G2 / M Verhaftung und zum Zelltod führt zu mitotischen Ausfall 4,5 zugeschrieben. Die molekularen Mechanismen, die extra Zentrosome de-Clustering werden intensiv untersucht <sup> 10. Das Verständnis dieses Mechanismus Tod ermöglichen exklusive Beseitigung von Krebszellen unter Schonung gesunden Gewebes 5,10.
So bieten Verbindungen, die mitotische Katastrophe Zelltod aktivieren einen neuen Modus eines selektiven Krebs-Therapie, die wirksam sein kann in einem breiten Spektrum von humanen soliden cancers.Our Ergebnisse legen nahe, dass konfokale Bildgebung verwendet werden, um Moleküle beeinflussen extra Zentrosome Clustering in identifizieren Mitose 2,3, wodurch diese Verbindungen Krebs Targeting Wirkstoffkandidaten.
Wir haben die zytotoxische Aktivität der Phenanthridin PJ-34 durch Abtasten festen und lebenden menschlichen Krebszellen (mit hohem Auftreten der extra-Zentrosome in Mitose) gegenüber normalen Zellen dokumentiert. Eine Schritt-für Schritt-Beschreibung der bildgebenden Verfahren verwendet, um die zytotoxische Aktivität der PJ-34 in menschlichen Krebszellen zu identifizieren sind unten aufgeführt.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Cell Culture Vorbereitung
MDA-MB-231-Zellen wurden von ATCC (American Type Culture Collection) erworben und in flüssigem Stickstoff gelagert.
- Seed 10 6 MDA-MB-231 Zellen in 92 mm Durchmesser Petrischale in 10 ml Vollmedium mit Dulbeco Modified Eagle Medium (DMEM), 10% Pferdeserum, 1% L-Glutamin und 1%-Penstrep Amphotericin B. Zellen zulassen bis zu etwa 80-100% Konfluenz vermehren.
- Entfernen Kulturmedium aus Schale und verwerfen.
- Waschen der Zellschicht kurz mit 0,25% (w / v) Trypsin-EDTA-Lösung, um alle Spuren von Serum zu entfernen.
- 2,0 ml Trypsin-EDTA-Lösung, austeilen und beobachten Zellen durch inverse Mikroskop bis Zellschicht verteilt ist (in der Regel innerhalb von 5 bis 15 min).
- In 18 ml komplettem Wachstumsmedium und sanft saugen die Zellen mit einer Pipette. Transfer zu einem Rohr.
- Zentrifuge die Zellsuspension bei 1.200 Umdrehungen pro Minute.
- Re-suspend das Zellpellet in 24 ml culture Medium.
- 2 ml Zellsuspension in 35 mm Petrischalen mit Glasboden (ca. 25% Konfluenz) und Platz im Inkubator (5% CO 2, 37 ° C).
- Lösungen:
- Komplette Medium für Zellen Zellproliferation: DMEM mit 10% FBS, 1% Antibiotika (100 Einheiten / ml Penicillin G, 100 ug / ml Streptomycin, Pen-Strep-Ampho Lösung) und 2 mM L-Glutamin.
- Trypsin-EDTA-Lösung, die 0,25% Trypsin-EDTA.
- Gerichte:
92 mm Durchmesser Petrischalen.
35 mm Durchmesser Poly-D-Lysin beschichteten Glasboden Kulturschalen.
2. Vorbereitung der Zellen für die Live-konfokale Imaging
- Seed 2 x 10 5 MDA-MB-231-Zellen in Glasbodenkulturschalen in 2 ml Vollmedium, wie in Kapitel 1 erwähnt. Wenn Zellkultur Zusammenfluss von 60-70% (ca. 3-4 x 10 5 Zellen pro Schale) erreicht, mit Transfektion gehen.
- Transfektion der Zellen mit zwei Plasmiden TonkanäleING die Fusionsproteine γ-Tubulin-GFP (für Fluoreszenzdetektion Zentrosome) und Histone-RED (H2b-RED, für Fluoreszenzdetektion der Chromosomen) mit dem liposomalen Transfektionsreagenz Jet-PI nach der Herstellung Protokoll. Kurz gesagt, mischen 2 ug von jedem Plasmid in einem Rohr mit 100 ul NaCl (150 mM). Mischen Sie das Transfektionsreagenz (100 ul) mit 100 ul NaCl (150 mM) in einem zweiten Rohr, und Inkubation 5 min bei Raumtemperatur (RT). Dann kombinieren Sie die beiden Lösungen, Mix (mit milden Vortex) und Spin-down. Inkubieren für 30 min bei RT.
- Während der Inkubation der Transfektionsgemisch, waschen Sie die Zellen einmal mit PBS und ersetzen Sie die Zell-Medium mit 2 ml warmem DMEM ohne Zusätze (37 ° C).
- Sanft fügen Sie die Transfektion Mischung zu den Zellen in DMEM und dann wieder die Zellen in den Inkubator (37 ° C, 5% CO 2) für 8 Stunden.
- Nach 8 h Inkubation, ersetzen Sie das DMEM mit 2 ml Vollmedium und Inkubation der Zellen im Brutschrank für 24hr.
- 24 Stunden nach der Transfektion, ersetzen Sie das Medium der Zellen mit 2 ml Vollmedium mit 20 uM PJ-34.
- Inkubieren Sie die Zellen für weitere 18 Stunden (37 ° C, 5% CO 2).
- Betrifft die Zellen konfokale Bildgebung für mindestens 16 Stunden zu leben in der Bildgebung Kammer halten die Zellen bei 5% CO 2 und 37 ° C.
- Parallel dazu untersuchen Transfektion Wirksamkeit 36 Stunden nach Transfektion unter Verwendung von Fluoreszenzmikroskopie wie folgt:
- Seed 2 x 10 5 MDA-MB-231-Zellen in 6-Well-Platte mit 1 Deckglas pro Vertiefung in 2 ml Vollmedium.
- Transfektion die Zellen wie in den Abschnitten 2.2-2.5 erwähnt.
- 36 Stunden nach Transfektion beheben die transfizierten Zellen auf einem Deckglas durch Inkubation montiert in kaltem Methanol: Aceton (1:1)-Lösung, 7 min, -20 ° C
- Saugen Sie die Fixierung Lösung und lassen das Deckglas mit den montierten Zellen in einer chemischen Haube trocknen.
- Bewerben ProLong Gold-antifade Reagenz mit DAPI und lassen the Deckglas im Dunkeln für 6 Stunden trocknen.
- Untersuchen Sie den Schieberegler unter dem Fluoreszenzmikroskop und berechnen den Anteil der transfizierten Zellen (rote und grüne Signale) von der gesamten Population von Zellen (DNA-Färbung durch DAPI). Die gewünschte Transfektion Prozentsatz etwa 20-40% bei 100-200 Zellen gezählt.
3. Technische Parameter der Live-konfokale Imaging Scanner-Einstellungen
- ScanMode xyzt; Pinhole [luftig] 1,00; Zoom 3,5; Auflösung 8 Bit; Laser DPSS 561 nm; Argon, sichtbaren Laser 488 nm; Laser He / Ne 633 nm sichtbar; Ziel HCX PL APO CS 63X 1.40 OIL UV; Numerische Apertur 1,4; Scan-Geschwindigkeit 700 Hz; Brechungsindex 1,52.
- Bild 3-D Präsentation wurden von Imaris Imaging-Software 7.0 vorbereitet.
4. Konfokalen Imaging der Mitose in fixierten Zellen
- Seed 2 x 10 5 MDA-MB-231 Brustkrebszellen (ATCC), normale embryonale Maus-Fibroblasten (MEF), oder PARP1 defizienten MEF (PARP - / -, hergestellt von Dr. Francoise Dantzer) auf Deckgläser in 6-Well-Platte in 2 ml Vollmedium. Die Deckgläser wurden mit 96% Ethanol gewaschen, gefolgt von Waschen mit sterilem DD Wasser gewaschen, 2 Stunden, und in jeder Vertiefung der 6-Well-Platte.
- In PJ-34 (10-30 pM) auf das Medium und Inkubation der Zellen für den gewünschten Zeitraum (in der Regel bis zu 96 Stunden).
- Waschen der Deckgläser einmal mit PBS (phosphate buffered saline), und fixieren die Zellen unter Verwendung von Inkubation in eiskaltem Methanol: Aceton (1:1)-Lösung, 7 min, 20 ° C
- Saugen Sie die Fixierung Lösung und lassen Sie die Deckgläser in chemischen Haube trocknen lassen (in diesem Stadium können die Deckgläser in -20 ° C für mehrere Wochen aufbewahrt werden).
- Waschen der Deckgläser einmal mit PBST (PBS mit 0,1% Tween-20 ergänzt), um die Zellmembranen durchlässig und blockieren die Zellen mit 10% NDS (Normale Eselserum) in PBST ("Blocking Solution") für 1 h bei RT.
- Inkubation der permeabilisierten fixierten Zellen mit primären Antibodies für 2 Stunden bei RT (für Spindeln und Zentrosome Färbung). Anti-α Tubulin (1:250 Verdünnung) und anti-γ Tubulin (1:200 Verdünnung): Die Antikörper werden in der Blocking-Lösung wie folgen verdünnt. Primäre Antikörper wie folgt angewendet: Anwendung 100 ul (in einem Tropfen) einer Mischung der Antikörper in Blocking-Lösung für jedes Deckglas auf 6-Well-Platte Abdeckung (der Deckel auf dem Kopf steht). Sanft legte das Deckglas auf dem Antikörper Tropfen, mit Blick auf die ausgesäten Zellen Drop. Inkubieren Deckgläser zugewandten Antikörper für 2 Stunden bei Raumtemperatur.
- Legen Sie die Deckgläschen wieder in den Brunnen und die Zellen 3-mal mit PBST. Dann nutzen Sie die gleiche Prozedur in 4.6 für die Markierung von Zellen auf den Deckgläsern mit den fluoreszierenden Sekundärantikörper beschrieben. Inkubieren der Zellen auf den Deckgläschen mit den sekundären Antikörpern für 1 Stunde, RT im Dunkeln. Die Antikörper werden in der Blocking-Lösung wie folgt verdünnt: und Alexa Fluor 568 (1:1000 Verdünnung Alexa Fluor 488 (grün 1:1000 Verdünnung);rot).
- Montieren Sie die Deckgläser mit ProLong Gold-antifade Reagenz mit DAPI (für die Chromosomen-Färbung) und Inkubation über Nacht bei RT in Dunkelheit, um zu trocknen.
- Untersuchen Sie die Deckgläser durch konfokale Mikroskopie.
5. Zellviabilitätsmessungen von ATP Produktion gemessen
ATP-Produktion wird durch eine leuchtende ATP Detektionsassay Kit gemessen.
- Seed die Zellen in 96-Well-Platte etwa 20.000 Zellen in 800 ul Medium in jeder Vertiefung. Drei leere Vertiefungen sollten für die Bestimmung der Hintergrundlumineszenz des Mediums verwendet werden.
- Bereiten ATP Standardverdünnungsreihen von etwa 10 um bis 100 um und halten auf dem Eis.
- In 50 ul Reinigungsmittel in jede Vertiefung und schütteln Sie die Platte für 5 min in Schüttler, 700 min.
- Den Inhalt jeder Flasche des "lyophilisierte Substrat 'mit 5 ml' Substratpuffers 'in dem Kit.
- In 50 ul des rekonstituierten Substrat-Lösung in die Vertiefungen, und schüttelndie Platte für 5 min auf Schüttler, 700 min.
- Halten Sie die Platte in Dunkelheit für 10 min.
- Messen Lumineszenz von jeder Vertiefung durch ELISA Mikroplatten-Reader.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34 ist eine stabile wasserlösliche Phenanthridin 1 (Abbildung 1). Unsere bisherigen Ergebnisse zeigen, Zelltod und extra Centrosomen in verschiedene Arten von festen Multi-zentrosomale Krebszellen, die mit PJ-34 behandelt wurden, de-Cluster. Im Gegensatz dazu wurden normalen proliferierenden Zellen nicht beeinträchtigt 2,3. Zentrosomen wurden durch doppelte Markierung mit Antikörpern gegen centrine1 und γ-Tubulin in den festen extra zentrosomale Zellen 2 gerichtet identifiziert.
Hier wurde die zytotoxische Aktivität der PJ-34 in diesen Live-extra zentrosomale Zellen in Echtzeit, indem Sie Live-konfokale Mikroskopie dokumentiert. Live-menschlichen Brustkrebszellen MDA-MB-231-Zellen, die ein hohes Vorkommen (> 50%) der Extra-Zentrosome 4,5 haben, wurden für mindestens 16 Stunden durch konfokale Bildgebung auf transfizierten Zellen mit γ-Tubulin-GFP konzentriert gescannt ( Fluoreszenzmarkierung von γ-Tubulin Brennpunkte 2) und Histone H2B-RED (fluoreszierend labeling der Chromosomen). Sechs bis zehn Live-transfizierten Zellen wurden parallel in jedem Experiment gescannt. Doppel Immunmarkierung der γ-Tubulin Brennpunkte in den transfizierten Zellen mit centrin1 war technisch unmöglich.
Verstreute γ-Tubulin-Herde und aberranten Chromosomen Anordnung wurden selten in zufällig ausgewählten unbehandelten MDA-MB-231-Zellen in der Mitose nachgewiesen. Bifocal Clustering von γ-Tubulin-Herde, was extra Zentrosome Bifokal Clustering, in der Mehrzahl der Live unbehandelte MDA-MB-231-Zellen (Abbildung 2) wurde dokumentiert, wurden dagegen Zentrosome und abweichende Anordnung der Chromosomen un-Cluster in Live erkannt transfizierten MDA-MB-231-Zellen mit PJ-34 (20 um) und Mitose in diesen Zellen durch Zelltod (Abbildung 3) endete inkubiert. Diese Zellen wurden mit PJ-34 für 18 inkubiert - 24 Stunden vor dem Scannen und für weitere 16 Stunden während des Scannens (Abbildung 3). Die Echtzeit-Dokumentation der Zelle death während der Mitose ausdrücklich einen zuvor definierten positiven Korrelation zwischen der Anzahl von menschlichen malignen Zellen mit multipolaren Spindeln in der Mitose und der Anteil der toten Zellen in Zellen mit PJ-34 (20 uM) 2 inkubiert.
PJ-34 wirkt als potenter Inhibitor PARP1 1. Wir haben daher die Möglichkeit geprüft PARP1 Hemmung verursacht Zelltod mit mitotischen Versagen verbunden. Anders als der Bildgebung, die feste MDA-MB-231-Zellen Prüfung einer großen Population von Zellen in der Zellkultur aktiviert, wodurch die statistische Analyse der Auswirkungen von PARP1 Inhibitoren in einer Vielzahl von menschlichen Krebszelllinien. (Abbildung 4) Die Tätigkeit des PJ-34 wurde auf die Aktivität von anderen potenten, nicht Phenanthren PARP1 Inhibitoren in normaler oder PARP1 defizienten Zellen - (- /) embryonalen Maus-Fibroblasten (MEF), dh normal und PARP1 () verglichen. PARP1 defizienten MEF Hafen Multi-Zentrosome in der Mitose, aber sie sind keint Tumorzellen 11. Diese Zellen wurden von Dr. Francoise Dantzer, Straßburg, Frankreich hergestellt.
Feste normal und PARP1 (- / -) MEF wurden für α-und γ-Tubulin, die ihre Spindeln und Zentrosome beschriftet, jeweils als vor 2 berichtet immunmarkiert. Einige der untersuchten Zellkulturen wurden mit PJ-34 oder anderen starken, nicht-Phenanthren PARP1 Inhibitoren, einschließlich ABT-888 und AG01469, das die enzymatische Aktivität von PARP1 hemmen und BSI-201, eine Verbindung, die scheinbar dämpft PARP1 Bindung zu behandelnden geklaut DNA 12-14. Keiner der getesteten PARP1 Inhibitoren beeinträchtigt normalen MEF bei Konzentrationen hemmen PARP1 Aktivität (Abbildung 4). Im Gegensatz dazu PJ-34 dosisabhängig un-Clustering von γ-Tubulin-Herde, Verzerrung von Spindeln und Zelltod in PARP1 verursacht (- / -) MEF (4A und B). Dies war nicht in normalen MEF mit PJ-34 behandelten Patienten beobachtet (Figure 4B) oder in PARP1 (- / -) mit MEF behandelt nicht phenenthrene PARP1 Inhibitoren ABT-888 oder AG014699 (4C). Es sei darauf hingewiesen, dass PJ-34 in einer Konzentration von mehr als 20 uM hat normalen MEF beeinträchtigen, obwohl normalen MEF waren resistent gegen PJ-34-Aktivität als PARP1 werden (- / -) MEF.
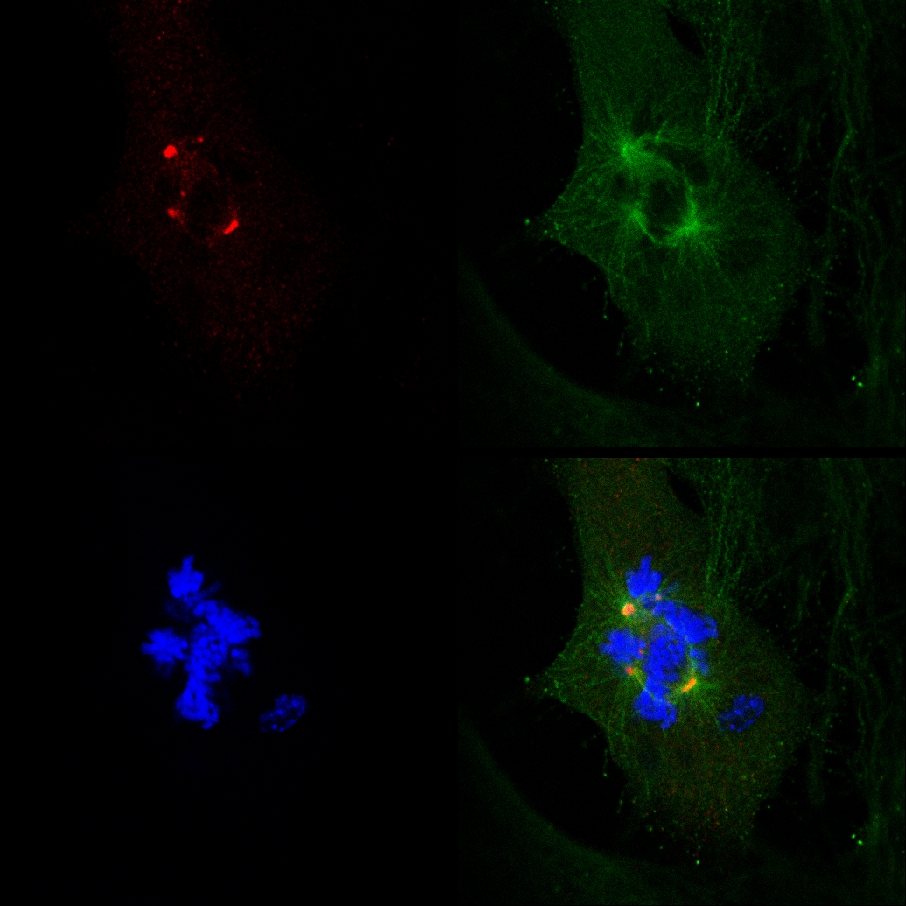
Die Tatsache, dass PJ-34 ausgerottet PARP1 (- / -) MEF trotz ihrer PARP1 Mangel, und die Korrelation zwischen der Bildung von multifokalen Spindeln und Zelle Tilgung in PARP1 (- / -) MEF mit PJ-34 bei Konzentrationen inkubiert höher als diejenigen für PARP1 Hemmung erforderlich, nicht im Einklang mit einer kausalen Verknüpfung zwischen extra Zentrosome de-Clustering in PARP1 (- / -) und MEF PARP1 Hemmung (Abbildung 4A). Die zytotoxische Aktivität von PJ-34 in PARP1 (- / -) MEF könnte besser durch seine Tätigkeit als ein extra-Zentrosome de-Clustering Agenten in Multi-zentrosomale Zellen 2 erläutert ( Ergänzende Informationen "/> Abbildung 3. Eine Live-konfokale Bildgebung Dokumentation des Zelltods in einem zufällig ausgewählten MDA-MB-231 Zellen in der Mitose mit un-Cluster γ-Tubulin markiert extra Zentrosome. MDA-MB-231-Zellen wurden mit PJ-34 für 24 Stunden vor dem Scannen und während der 16 Stunden von lebenden konfokale Abbildung inkubiert. PJ-34 wurde 24 h nach Transfektion mit Vektoren, γ-Tubulin-GFP (grün, γ-Tubulin-Herde und Zentrosome) und H2b-RED (Rot, Chromosomen) angewendet. 
Abbildung 1. Die phenanthridine PJ-34: N-(6-oxo-5 ,6-dihydro-phenanthridin-2-yl)-N, N-Dimethylacetamid. 
Abbildung 2. Bi-Clustering Schwerpunkte der Extra-Centrosomen in einer zufällig ausgewählten Live-MDA-MB-231 Zellen in der Mitose. A. Oberes Feld: Beschriftet Centrosomen in einer zufällig ausgewählten Live-MDA-MB-231 Zellen mit γ-Tubulin-GFP transfiziert Untere Tafel:. Chromosome Re-Arrangements während der Mitose in einer zufällig ausgewählten MDA-MB-231 Zellen mit Histone H2B-RED transfiziert. B. Bi-Schwerpunkt Mitose mit Clustered extra Centrosomen in einer zufällig ausgewählten Kult identifiziertgemessen MDA-MB-231-Zelle. Und Histone H2B-RED (Kennzeichnung Chromosomen; rot); Zellen wurden sowohl von γ-Tubulin-GFP (grün Kennzeichnung γ-Tubulin-Herde) transfiziert. 48 h nach der Transfektion wurden die Zellen zu einem Live-konfokale Abbildung für 16 Stunden ausgesetzt. Sechs Zellen wurden parallel in jedem Experiment gescannt. Vier verschiedene Experimente wurden durchgeführt. Siehe auch Ergänzende Informationen. Klicken Sie hier, um eine größere Abbildung anzuzeigen . 
Abbildung 3. Extra Zentrosome de-Clustering Zelltod in lebenden MDA-MB-231-Zellen mit PJ-34 behandelt voraus. Eine zufällig ausgewählte Live-MDA-MB-231 Zellen in der Mitose mit verstreuten Zentrosome (1. Frame links) durch Zelltod beendet ( 2. und 3. Frames). Diese ceund Histone H2B-RED (; ll wurde zufällig in einer Zellkultur für 24 h mit PJ-34 (20 um) angewendet 24 Stunden nach Transfektion mit Vektoren, γ-Tubulin-GFP (grün Kennzeichnung γ-Tubulin-Herde einschließlich Zentrosome) inkubiert ausgewählt Kennzeichnung Chromosomen; rot). Die Zelle wurde für 16 Stunden bei Live-konfokale Bildgebung gescannt. Sechs Zellen wurden parallel in jedem Experiment gescannt. Drei verschiedene Experimente wurden durchgeführt. Siehe auch Ergänzende Informationen. Klicken Sie hier, um eine größere Abbildung anzuzeigen . 
Abbildung 4. Eine zytotoxische Aktivität von PJ-34 in PARP1 (- / -) Maus embryonale Fibroblasten. A. (links) Der Prozentsatz der multifokalen Spindeln in n berechnetormal (schwarze Linie) und PARP1 - / - (graue Linie) MEF, für 48 Stunden mit PJ-34 lang bei den angegebenen Konzentrationen. Der Anteil der multipolaren Spindeln wurde aus insgesamt 20 Spindeln in 3 verschiedenen Experimenten nachgewiesen berechnet. (Rechts) Reduzierte Überleben der Zelle in Zellkulturen für 72 h mit PJ-34 (20 &mgr; M) in Bezug auf das Überleben der Kontrolle unbehandelten Zellen inkubiert erkannt (normal (schwarze Linie) und PARP1 - / - (graue Linie) MEF). Überleben der Zellen wurde durch die Zellen "ATP-Produktion (Protokoll 5) getestet. Die Mittelwerte von 4 Messungen für jede Zelllinie in 3 verschiedenen Experimenten dargestellt B. Spindeln in zufällig ausgewählten befestigt normal und PARP1 -. / - MEF in der Mitose, unbehandelt (Kontrolle) oder mit inkubiert PJ-34 für 48 Stunden bei der angegebenen Konzentrationen. PJ-34 verursacht multipolare Spindeln. Die Zellen wurden fixiert, permeabilisiert und immunmarkiert für α-und γ-Tubulin (grün Kennzeichnung von Spindeln und rote Kennzeichnung Zentrosome, respectively). Die Chromosomen wurden mit DAPI-Reagenz (blau) markiert. . Repräsentative Ergebnisse aus 3 verschiedenen Experimenten C. Potent nicht Phenanthren PARP1 Inhibitoren keinen Einfluss Zentrosome Clustering in PARP1 (- / -) MEF. Spindeln von zufällig ausgewählten normalen und PARP1 - / - MEF dargestellt; unbehandelten MEF (Kontrolle) oder MEF 48 h mit nicht Phenanthren PARP-Inhibitoren, AG01469 (20 uM) oder ABT888 (20 uM) behandelt. Chromosomen sind mit DAPI-Reagenz (blau) markiert. Ähnliche Ergebnisse wurden in 3 verschiedenen Experimenten erhalten. Klicken Sie hier, um eine größere Abbildung anzuzeigen .
Klicken Sie hier, um eine größere Abbildung anzuzeigen . 
Klicken Sie hier, um eine größere Abbildung anzuzeigen .
2B. Supplemental Abbildung. Ein zufällig ausgewähltes MDA-MB-231 Zellen in der Anaphase mit bi-Schwerpunkt gruppierten γ-Tubulin markiert extra Zentrosome wurde für 16 Stunden bei Live-konfokale Bildgebung, 48 Stunden, nachdem er mit Vektoren, γ-Tubulin-GFP transfiziert gescannt und H2b-RED (Etikettierung γ-Tubulin-Herde und Centrosomen in den fixierten Zellen (grün) und H2b Histon Kennzeichnung der Chromosomen (rot), respectively).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Live-konfokale Bildgebung bot eine Echtzeit-Dokumentation der zytotoxischen Wirkung der PJ-34 im Live-Multi-zentrosomale Zellen während der Mitose (Abbildung 3 und ergänzende Informationen). Dies war die erste Live-Dokumentation zuschreibt die Zytotoxizität von PJ-34 in menschlichen Krebszellen, um zusätzliche Zentrosome de-Clustering und Zelltod, was darauf hindeutet, Induktion von Mitotic Catastrophe Zelltod durch PJ-34 5-9. Im Gegensatz dazu wurde bi-fokale Clustering von super-Numerarier Centrosomen in Live unbehandelte MDA-MB-231 Zellen, die eine normale Mitose mit bi-Focali gruppierten zusätzliche Zentrosome (Abbildung 2, zusätzliche Informationen) beobachtet.
Nach diesen Ergebnissen leben konfokale Bildgebung von transfizierten Zellen nützlich sein könnte für Detektion der Translokation von Proteinen in extra Zentrosome Clustering in transfizierten Zellen während der Mitose 5,9,15,16 verwickelt. Identifizierung von Proteinen durch PJ-34 Multi-centros betroffenomal Zellen können einige Hinweise für das Verständnis der Mechanismen, durch Tod extra Zentrosome de-Clustering aktiviert bereitzustellen.
Der Vorteil von lebenden konfokale Abbildung der Bereitstellung der Informationen in Echtzeit während der Mitose in lebenden Zellen ist auch mit mehreren Einschränkungen gebunden. Die Anzahl der Zellen pro Experiment abgetastet wird begrenzt. Daher Chancen Mitosezellen erkennen sind niedrig, und mehrere sich wiederholende Versuche für eine Echtzeit-Dokumentation der Mitose in den gescannten Zellen erforderlich. Darüber hinaus ist der Erfolg stark von hoher Effizienz der Transfektion von Zellen mit Vektoren, markierten Proteinen. So, obwohl sie zuverlässig, ist Live-konfokale Bildgebung zeitaufwendig und erfordert sehr erfahrene harte Arbeiter.
Im Vergleich Immunzytochemie und konfokale Abbildung fixierten Zellen ermöglichen Untersuchung einer großen Anzahl von Zellen pro Experiment, das für eine zuverlässige statistische Analyse erforderlich ist. Wir nutzten diese Methode, um das zu vergleichenAuswirkungen der PJ-34 in der normalen Mitose gutartigen Zellen auf ihre Auswirkungen in Krebszellen Mitose mit überzähligen Centrosomen 2,3. In ähnlicher Weise wurde diese Methode für die Wirkungen von PJ-34 zu der nicht-Phenanthren PARP1 Inhibitoren in PARP verwendete (- / -) MEF beherbergen extra Zentrosome (Zellen mit hoher Auftreten von mehreren Zentrosome 11) (Abb. 4).
Zusammenfassend zeigen unsere Ergebnisse, den Vorteil der Kombination der wertvollsten Informationen in Echtzeit durch konfokale Bildgebung von lebenden Zellen in der Mitose (Abbildungen 2 und 3) mit Zytochemie und konfokale Analyse der fixierten Zellen zur Verfügung gestellt. Eine Kombination dieser Verfahren kann nützlich sein für die Identifizierung der cytotoxischen Aktivität von kleinen Molekülen, wie PJ-34, gezielt bestimmte Mechanismen von entscheidender Bedeutung für das Überleben der Zelle. Die einzigartige Abhängigkeit vieler menschlichen Krebszellen auf extra-Zentrosome bi-polar Clustering für ihre Proliferation und Hinterbliebenenal macht PJ-34 ein möglicher Kandidat für die Krebstherapie.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Die Autoren erklären, dass sie keine finanziellen Interessen haben.
Acknowledgments
Finanzierungsquellen dieser Forschung: ein gemeinsamer Fonds von der Universität Tel Aviv Technologietransfer Firma, RAMOT und dem Sheba Medical Center-(M. CA und SI.), ICRF - Israeli Cancer Research Foundation (M. CA.) Und Israel Science Foundation ( SI).
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S.
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}