

Summary
واستخدمت البروتينات تاو معدلة وhyperphosphorylated في اثنين من فحوصات في المختبر تجميع للكشف عن حركية تجميع سريع تعتمد على hyperphosphorylation. هذه المقايسات تمهد الطريق لشاشات المستقبلية للمركبات التي يمكن أن تعدل بميل تاو hyperphosphorylated لتشكيل الألياف التي تكمن وراء تطور مرض الزهايمر.
Introduction
مرض الزهايمر (AD) هو واحد من مجموعة كبيرة من الاضطرابات العصبية المعروفة باسم tauopathies. في الجوهر tauopathy علم الأمراض الكامنة هو التشابك الليفي العصبي، NFTS، في الخلايا العصبية، الخلايا النجمية والخلايا الدبقية الصغيرة 1-4. كثافة NFT يرتبط مع ضعف الادراك 3،5 والخلايا العصبية وفقدان 6. NFT يحتوي على بروتين تاو hyperphosphorylated في المقام الأول (المشار إليها باسم "P-تاو" من الآن فصاعدا) التي تشكل خيوط مستقيمة أو حلزونية تقرن (PHF) 7،8. تاو هو بروتين يرتبط أنيبيب يعتقد لتسهيل النقل المحاور التي لا غنى عنها للإشارات العصبية والاتجار 9،10. كل جزيء تاو يحتوي على 2 إلى 3 الفوسفات في الدماغ العادي، ولكن زيادة المحتوى فسفوريل عدة طيات في المرضى الذين tauopathy 11. من المحتمل أن تساهم في تاو hyperphosphorylation بما في ذلك GSK3β (الجليكوجين سينسيز كيناز 3β) وCDK5 (السيكلين دي تحركات متعددةكيناز معلقة 5) 12،13، ولكن على الزناد المباشر للالفسفرة المرضية لا يزال بعيد المنال 14. الفسفرة غير طبيعية في أو بالقرب من الزخارف ملزم أنيبيب تنأى تاو من أنيبيب 15، ويسبب تاو سوء الترجمة إلى المقصورة somatodendritic، حيث oligomerizes ف تاو إلى خيوط مستقيمة أو حلزونية تقرن التي يمكن أن تتبلمر في نهاية المطاف إلى الادراج NFT. أدى التعادل وثيق بين hyperphosphorylation تاو، وتشكيل NFT، والتنكس العصبي إلى فرضية السائدة التي التشابك ف تاو الحصول على ردود السامة للخلايا أفكارك وغيرها، وبالتالي هي السبب الكامن وراء tauopathy تنكس عصبي 16،17. تم إطلاق شاشات المخدرات والاختبارات السريرية في وقت مبكر على أساس هذا الافتراض 18. ومع ذلك، تواجه هذه الفرضية التحديات 19،20. على سبيل المثال، سانتا كروز وآخرون أظهرت أن الوظائف المعرفية من الفئران المعدلة وراثيا يمكن أن تتحسن عن طريق قمع التعبير عن المسختاو البشري، على الرغم من استمرار NFTS لتشكيل جزيئات من تاو القائمة 21. في نموذج ذبابة الفاكهة، فقد تبين NFT لعزل وعصاري خلوي تاو السامة لحماية الخلايا العصبية الكامنة 22،23. ومن الواضح أن دور التسبب في NFT، إن وجدت، سوف تؤثر بشكل كبير على اتجاه التنمية العلاجات tauopathy.
في تركيزات عالية، المؤتلف أو العادي بروتين تاو الدماغ بشكل عفوي ولكن ببطء يبلمر إلى بنية تشبه PHF في المختبر، كما يتبين من ربط عدة ورقة β فضل الأصباغ الفلورية، المجهر الإلكتروني، وتشتت الضوء الطيفي 24-27. مضيفا الهيبارين أو حمض الأراكيدونيك، وهو من الأحماض الدهنية وفرة في دماغ الإنسان، ويسرع بشكل كبير في تشكيل PHF isoform- تاو ومحفز تعتمد على التركيز الخلق 28-32. يثير الاهتمام والفضول، تنقية تاو hyperphosphorylated من أدمغة AD أو عن طريق شاملة أعدت في تفاعلات الفسفرة المختبر لggregates أسرع وأكثر كفاءة 26،33-35. هذه النتائج هي في الاتفاق ممتازة مع الأدوار المرضية للف تاو. وبالتالي يجوز للنظام في المختبر على أساس تجميع ف تاو بمثابة أداة قوية لفحص AD المخدرات.
ونظرا لارتباط وثيق بين تجميع تاو والتنكس العصبي التدريجي للم، وكذلك فشل الأخير في تطوير العقاقير التي تستهدف وحة Aβ، علامة أخرى النسيجية الرئيسية 36-38 م، والفائدة في اكتشاف العقاقير التي تتحكم تاو تجميع آخذ في الارتفاع. في الواقع، العديد من الجماعات قد بدأت بالفعل شاشات المخدرات في الإنتاجية المختلفة، وذلك باستخدام المختبر في ردود الفعل تاو التجميع كما الفحص الابتدائي. تم العثور على عدد من المواد الكيميائية لعرض الأنشطة المثبطة أو انعكاس على تاو التجميع في المختبر 39-42. ومع ذلك، كل الشاشات الحالية منظم تاو تاو تجميع تستخدم معدلة أن يفتقد العلامة المرضية الرئيسية من الفوسفورylation، ورفع مصدر قلق لخصوصية وفعالية استخدام هذه المركبات في علاج م.
واحدة من العقبات الرئيسية لتطوير المقايسات تجميع لتوصيف والكيمياء الحيوية وفحص AD المخدرات هو إنتاج كميات كافية من البروتين تاو hyperphosphorylated ذات الصلة pathophysiologically. باستخدام نظام الحفز سحابات بمساعدة الذي شكل الإسوي 1N4R من تاو وكيناز GSK-3β وشارك أعرب في E. القولونية كما يسين سستة البروتينات الانصهار، وتغلبنا على هذا التحدي (. سوي وآخرون، المقدمة؛ انظر الشكل 1 للمنتجات النهائية تاو وف تاو، انظر أيضا 43 لأولي الشامل توصيف الطيف من ص-تاو). من لجنة من تسعة أجسام مضادة محددة لمواقع الفسفرة مختلفة من تاو، وشوهدت إشارات إيجابية في ثمانية مواقع (لا تظهر البيانات). أدناه، وصفنا البروتوكولات وأدوات القياس التي يمكن أن تفرق تجميع د الحركيifferences بين تاو معدلة والأنواع ف تاو. تم تعديل هذه المقايسات من البروتوكولات نشرت أن تقاس زيادة مضان من thioflavin T (تى اتش تى) أو thioflavin S (تي إتش إس) عند اميلويد (المجاميع تاو) ملزمة 26. في أول "محطة"، نهج لا صبغ، يتم تجميع ردود الفعل تجميع وحضنت في غياب الصبغة اميلويد. في نقاط زمنية مختلفة، تتم إزالة قسامة من كل رد فعل ومختلطة مع حجم مساو من المخزن المؤقت تى اتش تى التي تحتوي على وقف التجميع والسماح تى اتش تى لربط المجاميع تاو. ويقاس مضان من قبل IAP FluoroMax-2 التألق. في المجموعة الثانية "مع صبغ" مراقبة الفحص المستمر، يتم تضمين تى اتش تى أو تي إتش إس في ردود الفعل التجميع. ويمكن قياس مضان باستمرار طوال التجربة بأكملها يدويا أو باستخدام قارئ لوحة متعددة. وبالإضافة إلى ذلك، نحن تصف مقايسة يستخدم تركيز شبه الفسيولوجية من تاو وف تاو للتجميع في مو قياس مستمردي. تأثير الفسفرة يبقى كشفها بسهولة. أدناه، فإننا سوف تصف الإجراءات العملية خطوة بخطوة، وتظهر نتائج ممثلة من هذه المقايسات. سوف مناقشة بعض من إيجابيات وسلبيات كل نهج، فضلا عن التطبيقات المحتملة متابعة فحص المخدرات.
بتركيز عال، تاو المجاميع في هياكل تشبه اميلويد من تلقاء أنفسهم. ومع ذلك، في المختبر، وعادة ما تسارع تاو fibrillization من هذه المحرضات كما الهيبارين (متوسط الوزن الجزيئي، 6،000 جم / مول) وحمض الأراكيدونيك. الأمثلة الموضحة في هذه الوثيقة تتضمن 30 ميكرومتر الهيبارين. ويتم رصد تشكيل تاو المجاميع اميلويد من قبل مضان الناتجة عن اميلويد ملزمة thioflavin T (تى اتش تى) أو thioflavin S (تي إتش إس). على ملزم لالمجاميع تاو، تى اتش تى يسلك التحول الأحمر في مضان (الإثارة: 450 نانومتر؛ الانبعاثات ذروتها: 485 نانومتر). تي إتش إس، من ناحية أخرى، لديها ضعف الانبعاثات في 510 نانومتر (الإثارة في 450 نانومتر) قبل اميلويد ملزمة، ولكن هذا fluorescencزيادات الإلكترونية بشكل كبير في وجود بروتين اميلويد مثل تاو مجمعة 44. كلا الأصباغ تعمل بشكل جيد في الكشف عن تاو وف تاو التجميع. بسبب ذروة الانبعاثات قوية واسعة نسبيا من تى اتش تى (انظر الشكل 2)، وهناك فقط 30٪ تخفيض في وحدة مضان في 510 نانومتر. للراحة، ونحن نستخدم نفس مزيج من موجات الإثارة / الانبعاثات (أي 450 نانومتر / 510 نانومتر) لمراقبة تاو تجميع عند استخدام إما صبغ.
ويمكن أن يتم تجميع تاو في وجود أو عدم وجود الصبغة، وهذا يتوقف على الغرض من الفحص وتوافر البروتين تاو. وتظهر كل من وسائط من ردود الفعل أدناه. وبالإضافة إلى ذلك، ونحن لشرح تشغيل اثنين من الأدوات المختلفة - مقياس التألق عينة واحدة (ISA-سبكس FluoroMax-2) وقارئ متعدد لوحة (SpectraMax M2). وينبغي أن تكون قادرة على التكيف مع هذه البروتوكولات لتناسب احتياجاتهم الخاصة وتوفر أداة القراء.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. إعداد الكواشف
- إعداد عازلة تجميع (20 ملي تريس، ودرجة الحموضة 7.4، 100 مم كلوريد الصوديوم، 1 ملم EDTA). تخزين في RT ومستقرة لعدة أشهر. الملحق 1 ملي dithiothreitol (DTT) قبل الاستخدام.
ملاحظة: منطقة عازلة على أساس HEPES (10 ملي HEPES، ودرجة الحموضة 7.5، 0.1 ملي EDTA، 5 ملي DTT) كما تنتج نتائج مماثلة في تاو التجميع. - إعداد thioflavin T أو thioflavin S حل الأوراق المالية (3 ملم، وحلت في المخزن التجميع)، وتصنف 0.22 ميكرون وحدة تصفية العقيمة. تخزينها في -20 درجة مئوية في أنبوب من رقائق الألومنيوم غطت ومستقرة لعدة أشهر.
- إعداد محلول المخزون الهيبارين (300 ميكرومتر، وحلت في المخزن التجميع). تخزينها في -20 ° C ومستقر لعدة أشهر.
- إعداد dithiothreitol (DTT) الأسهم (1 M، يذوب في الماء). قسامة إلى 1.5 مل أنابيب. تخزين في -20 ° C. قبل المقايسات التجميع، وذوبان الجليد الحل 1 M في RT. من هذا المخزون 1،000x، وإعداد قسامة من 100 ملي الأسهم العمل مع الماء منزوع الأيونات. ترك على الجليدحتى جاهزة.
- إزالة تاو من -80 ° C الفريزر. ذوبان الجليد على الجليد. ضبط تاو لتركيز محددة سلفا مع المخزن المؤقت التجميع. تدور في microcentrifuge في 20،800 x ج لمدة 10 دقيقة في 4 درجات مئوية لإزالة الركام كبيرة شكلت مسبقا. هذه الخطوة ما قبل الغزل يزيد الاتساق في قياس مضان لاحق من كل دفعة من الإعدادية البروتين. نقل طاف لأنبوب آخر؛ ترك على الجليد حتى جاهزة لتجميع رد فعل التجميع.
2. لا صبغ، الفحص الطرفية
ويتم تجميع رد فعل من هذا الاختبار في حالة عدم وجود صبغة الفلورسنت: ملاحظة. بعد خلط جميع المكونات، ولا يسمح للرد فعل للشروع في نقاط زمنية محددة مسبقا. ثم يتم أخذ مأخوذة من رد فعل تجميع ومختلطة مع تى اتش تى أو تي إتش إس للاميلويد ملزم قبل القراءة مضان. حجم الأولي من رد فعل تجميع يعتمد على عدد من النقاط الوقت اللازم. هذا النهج قد REQUغضب على كمية كبيرة من البروتين تاو، ولكن بشكل سريع، واضحة، ويمكن القيام به في التألق أو قارئ لوحة متعددة جيدا (انظر مناقشة). هو عملية خطوة بخطوة أدناه، وذلك باستخدام الطيفي المدمجة ISA سبكس FluoroMax-2 لمضان الكمي.
- إعداد خليط تجميع في 1.5 مل أنابيب إيبندورف كما في الجدول 1. ويمثل كل عمود المكونات اللازمة للتفاعل 100 ميكرولتر، وهو ما يكفي لأحد قياس الوقت نقطة. ضبط كمية من أجل مزيج تجميع كامل على أساس النقاط الوقت اللازم للتجربة معينة. إضافة إضافية بنسبة 10٪ لكل مكون لإعطاء مجال للخطأ pipetting ل. رد فعل تحتوي على الهيبارين نموذجي هو موضح هنا يمكن الاستعاضة عن حمض الأراكيدونيك أو عازلة التجميع. إضافة إلى DTT 1 ملم إلى خليط التفاعل. إذا يستمر رد الفعل كله أكثر من يوم واحد، تكملة جديدة DTT كل يوم (1 ملم) لضمان بيئة الحد.
- عكس الأنبوب بضع مرات لخلط. جيش التحرير الشعبى الصينىCE كل رد فعل في الحاضنة 37 درجة مئوية أو ماء الحمام. ليست هناك حاجة للالتحريض تاو التجميع.
- قبل قياس مضان، بدوره على الطيفي (مصباح أولا، ثم الكمبيوتر).
ملاحظة: مصباح قوس زينون التي يمكن استخدامها على الفور. ومع ذلك، لأفضل النتائج تسمح للآلة في عملية الاحماء لمدة 10 دقيقة قبل قراءة مضان. - بدء تشغيل البرنامج على جهاز الكمبيوتر.
- اختيار الوقت الحقيقي عرض الوضع في مركز التحكم صك، تعيين الطول الموجي الإثارة إلى 450 نانومتر (الشق إلى 2 نانومتر) والطول الموجي الانبعاثات إلى 510 نانومتر (شق إلى 5 نانومتر). وثيق في الوقت الحقيقي نافذة عرض الوضع للعودة إلى مركز التحكم الصك.
- اختر تحليل الطول الموجي، اضغط إضافة >> مفتاح المستمر في الإطار العلوي لإضافة موجات مجموعات. تعيين المعلمات الاستحواذ على خطأ قياسي ل1 والمحاكمات القصوى إلى 3، ثم انقر فوق إضافة. انقر فوق انتقال! لفتح بيانات العرضالنافذة.
- في نافذة العرض بيانات، انقر فوق ابدأ ACQ لفتح مربع الحوار نموذج جديد. اختيار "غير معروف" لنوع العينة.
- إلى كل 100 ميكرولتر خليط التجميع، إضافة 98 ميكرولتر عازلة تجميع و 2 ميكرولتر 3 ملي thioflavin T. الماصة عدة مرات لخلط.
- نقل الخليط بأكمله إلى كوفيت (FCA3، والبعد الخارجي، wxlxh = 12.5 مم × 12.5 مم × 45 مم). ضع كفيت في حامل العينة في عينة مقصورة وإغلاق الغطاء. انقر فوق تشغيل لجمع البيانات مضان. تسجيل البيانات.
- إزالة كفيت وصب الحل. شطف كفيت بواسطة الماء المقطر 3 مرات. الجافة التي تهب الهواء داخل وخارج كفيت.
3. مع صبغ، المستمر الوضع الفحص على لوحة قارئ SpectraMax M2
ملاحظة: هذا الاختبار يختلف عن سابقتها في أن يتم تضمين صبغة الفلورسنت أو تي إتش إس تى اتش تى في aggregaرد فعل نشوئها. وهذا يسمح القياس المستمر من نفس مجموعة من ردود الفعل. بسبب استخدام المتكرر من رد الفعل، هو أفضل فعلت هذه الطريقة مع التلقائي متعددة جيدا قارئ لوحة (كما هو موضح أدناه تشغيل SpectraMax M2). يعمل والتألق العادية أيضا ولكن وتيرة قياس ردود الفعل تجميع سريع محدودة نوعا ما، نظرا لطبيعة اليدوي للعملية.
- إعداد مزيج التجميع في لوحة 96-جيدا (96 جيدا لوحة سوداء صلبة، جيدا حجم 360 ميكرولتر، مسطحة القاع) كما في الجدول (2). ويمثل كل عمود المكونات اللازمة للتفاعل 200 ميكرولتر، وهو ما يكفي لواحد قياس الوقت نقطة. مزيج جيد من قبل pipetting عدة مرات. تكملة جديدة 1MM DTT كل يوم طوال فترة التجارب.
- احتضان لوحة 96-جيدا عند 37 درجة مئوية.
- في كل نقطة زمنية قبل قياس مضان، بدوره على وضع متعددة قارئ صفيحة ميكروسكوبية والكمبيوتر. إتاحة الوقت الكافي للأماهعمود فقري لتحقيق الاستقرار، حوالي 10 دقيقة.
- بدء تشغيل البرنامج على جهاز الكمبيوتر. ضبط درجة الحرارة إلى 37 درجة مئوية، واختيار شدة الإسفار (الأعلى FI مقروءة) واسطة، تعيين الطول الموجي الإثارة في 450 نانومتر، والانبعاثات الطول الموجي في 510 نانومتر.
- إدراج لوحة 96-جيدا في درج واضغط على مفتاح READ لبدء القياس.
- بعد القراءة، وإزالة لوحة والعودة مرة أخرى إلى C حاضنة 37 درجة. نسخ البيانات ولصقها في جدول بيانات Excel لتحليل البيانات والتآمر.
4. مع صبغ، المستمر الوضع الفحص الطيفي على الاتفاق
- إعداد خليط تجميع في 1.5 مل أنابيب إيبندورف كما في الجدول 3. ويمثل كل عمود المكونات اللازمة للتفاعل 200 ميكرولتر، وهو ما يكفي لأحد قياس الوقت نقطة.
- عكس الأنبوب بضع مرات لخلط.
- بدوره على الطيفي ووضع البرنامج كما في الخطوات 2.3 و 2.4.
- نقل الخليط بأكمله رالزراعة العضوية كفيت. ضع كفيت في حامل العينة في عينة مقصورة وإغلاق الغطاء. انقر فوق تشغيل لجمع البيانات مضان. تسجيل البيانات.
- مواصلة القراءة على فترات مناسبة من خلال النقر تشغيل وتسجيل البيانات. إذا كان التجميع هو أن يتم رصدها في ذات تردد عال (على سبيل المثال، كل 30 أو 60 ثانية)، وترك رد الفعل في كفيت وفي الجهاز حتى إما الانتهاء من القياس، أو عندما يكون هناك وقت كاف لمبادلة ردود الفعل أو cuvettes.
- إزالة كفيت وصب الحل. شطف كفيت بواسطة الماء المقطر 3 مرات. الجافة التي تهب الهواء داخل وخارج كفيت.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
باستخدام تاو المؤتلف وف تاو (الشكل 1)، أنشأنا اثنين من بروتوكولات مختلفة للمقارنة بين حركية تجميع تاو وف تاو، والاستفادة من انبعاث مضان قوي من تى اتش تى وتي إتش إس عليه وملزم لamyloidal المجاميع البروتين، بما في ذلك تاو وف تاو (الشكل 2). مع أو بدون صبغة الفلورسنت في رد فعل التجميع، لاحظنا تعزيز ثابت من تاو التجميع التي كتبها hyperphosphorylation (الأرقام 3-5). هذا التحفيز هو مستقل من الهيبارين (لا تظهر البيانات). في رد فعل، وتاو تاو ف oligomerize نموذجية في معدلات سريعة خلال 30 دقيقة الأولى أمام تباطؤ ملحوظ (أرقام 3 و 5)، مع ف تاو واظهار وحدة مضان أعلى طوال فترة التجربة. بما في ذلك تى اتش تى في ردود الفعل تجميع يسبب التخلف الكبير في معدل التجميع (الشكل 4). كل من الأشكال الإسوية بلات اقتربقد مياه 160 ساعة بعد ردود الفعل التي. تي إتش إس، من ناحية أخرى، لا يسبب تباطؤ ملموس التجميع (الشكل 5).

الشكل 1. تنقية تاو تاو وhyperphosphorylated (ع-تاو) المستخدمة في هذه الدراسة. تم حل عينات بنسبة 10٪ هلام SDS-PAGE، وملطخة بواسطة Coomassie الأزرق R250 (يسار) أو سبر من قبل وحيدة النسيلة الأجسام المضادة لمكافحة تاو (يمين لوحة). حارة M، الوزن الجزيئي علامة. الممرات 1 و 3، تاو unphosphorylated. lane2 و 4، تاو hyperphosphorylated.

الشكل 2. الانبعاثات الأطياف لتى اتش تى (30 ميكرومتر) مع أو بدون ملزمة المجاميع تاو. وكان استحواذ الانبعاثات المعلبة من 460 نانومتر إلى 600 نانومتر (1 نانومتر زيادة؛ والتكامل 0.1 ثانية، 5 نانومتر عرض الشق)، الإثارة في 450 نانومتر. تم الحصول على المجاميع تاو خلال السماح 50 ميكرومتر تاو تجميع بالمضي قدما عند 37 درجة CO / N (انظر بروتوكول 2 لمزيد من التفاصيل).
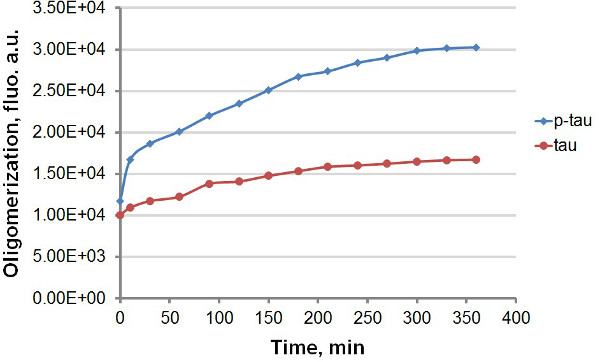
الشكل 3. منحنيات تجميع لتاو وف تاو في فحص المحطة. تجميع 50 ميكرومتر تم الانتهاء تاو وف تاو مع 30 ميكرومتر الهيبارين باعتبارها محفز. في أوقات مختلفة بعد بدء التفاعل، تمت إزالة 100 ميكرولتر من رد فعل ومختلطة مع نفس الحجم من 60 ميكرومتر تى اتش تى قبل قياس مضان. وقد تم قياس مضان في 450 نانومتر الإثارة، 510 الانبعاث نانومتر. "الاتحاد الافريقي"، وحدات التعسفي. لاحظ أن النطاق الزمني في غضون دقائق.
"الشكل 4" SRC = "/ الملفات / ftp_upload / 51537 / 51537fig4highres.jpg" />
الشكل 4. منحنيات تجميع لتى اتش تى وحده، تاو وف تاو في وضع القياس المستمر في وجود تى اتش تى. تألفت كل رد فعل من 0 أو 50 ميكرومتر تاو أو ف تاو، 30 ميكرومتر الهيبارين، و 30 ميكرومتر تى اتش تى في المخزن التجميع . وحضنت ردود الفعل عند 37 درجة مئوية في لوحة 96-جيدا. في نقاط زمنية مختلفة، تم إزالة لوحة من الحاضنة وتحميلها للقارئ لوحة لمضان القراءة (الإثارة 450 نانومتر، وانبعاث 510 نانومتر). بين القراءات، وكان يحتفظ لوحة في الحاضنة دون التحريض تحت غطاء. وجود تى اتش تى تباطأ بشكل كبير بانخفاض التجميع، ولكن، الأهم من ذلك أن تاو hyperphosphorylated لا يزال عرضت بمعدل أسرع من التجميع مما فعلت نظيرتها معدلة. لاحظ أن حجم الوقت في ساعات.
pload / 51537 / 51537fig5highres.jpg "/>
تم تقييم الشكل 5. على نطاق صغير المقايسات تاو تجميع بحضور تي إتش إس. في المختبر الناجم عن الهيبارين تجميع تاو 6 ميكرومتر وف تاو في وضع القياس المستمر مع thioflavin سنغافوري كما الصبغة المؤشر. وبالإضافة إلى البروتين، وتضمن كل رد فعل 30 ميكرومتر الهيبارين و 20 ميكرومتر تي إتش إس في HEPES تجميع عازلة (10 ملي HEPES الرقم الهيدروجيني 7.5، 5 ملي DTT، 0.1 ملي EDTA). جميع المكونات ما عدا الهيبارين كانت مختلطة والمتوازنة في RT. بعد إضافة الهيبارين، تم نقل رد فعل على كفيت ووضعها في حامل العينة. وسجلت مضان على الفور كما T 0، واستمر لمدة 2 ساعة أو حتى تباطأت الزيادة مضان وصولا الى ما يقارب الصفر. بسبب فترة رد فعل قصيرة نسبيا، تم تنفيذ رد فعل كامل بها في RT في نفس كفيت.
| تاو | ف تاو | |
| 60-100 ميكرومتر تاو | 50 ميكرولتر | 0 ميكرولتر |
| 60-100 ميكرومتر ف تاو | 0 ميكرولتر | 50 ميكرولتر |
| 300 ميكرومتر الهيبارين | 10 ميكرولتر | 10 ميكرولتر |
| عازلة التجميع | 39 ميكرولتر | 39 ميكرولتر |
| 100 ملي DTT | 1 ميكرولتر | 1 ميكرولتر |
الجدول 1. مكونات تجميع الخليط لمدة لا صبغ، فحص المحطة.
| تاو | ف تاو | صبغ وحدها | |
| 60-100 ميكرومتر تاو | 50 ميكرولتر | 0 ميكرولتر | 0 ميكرولتر |
| 60-100 ميكرومتر ف تاو | 0 ميكرولتر | 50 ميكرولتر | |
| 300 ميكرومتر الهيبارين | 20 ميكرولتر | 20 ميكرولتر | 20 ميكرولتر |
| 3 ملي thioflavin T | 2 ميكرولتر | 2 ميكرولتر | 2 ميكرولتر |
| عازلة التجميع | 126 ميكرولتر | 126 ميكرولتر | 176 ميكرولتر |
| 100 ملي DTT | 2 ميكرولتر | 2 ميكرولتر | 2 ميكرولتر |
الجدول 2. تجميع مكونات الخليط لمع صبغ، الفحص المستمر على قارئ لوحة.
| تاو | ف تاو | صبغ وحدها | |
| 60-100 ميكرومتر تاو | 5 ميكرولتر | 0 ميكرولتر | 0 ميكرولتر |
| 60-100 ميكرومتر ف تاو | 0 ميكرولتر | 5 ميكرولتر | 01؛ ل |
| 300 ميكرومتر الهيبارين | 20 ميكرولتر | 20 ميكرولتر | 20 ميكرولتر |
| 3 ملي thioflavin S | 1.5 ميكرولتر | 1.5 ميكرولتر | 1.5 ميكرولتر |
| عازلة التجميع | 171.5 ميكرولتر | 171.5 ميكرولتر | 176.5 ميكرولتر |
| 100 ملي DTT | 2 ميكرولتر | 2 ميكرولتر | 2 ميكرولتر |
الجدول 3. تجميع مكونات الخليط لمع صبغ، الفحص المستمر على الطيفي المضغوط.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
هذا البروتوكول يوضح شروط الفحص المختلفة والأدوات التي كشف تاو سريع حركية التجميع التي تعتمد على الفسفرة. في مقايسة المحطة، يتم إضافة الصبغة مضان تى اتش تى لجزء من رد فعل إزالتها من مزيج الرئيسي في كل نقطة زمنية. اميلويد يسببها ملزم ثم يقاس مضان 26. في المجموعة الثانية، مع صبغ واسطة، ويتم تجميع تاو في وجود تى اتش تى أو تي إتش إس، مما يجعل هذا النوع من رد الفعل المناسب في الوقت الحقيقي للتقييم التلقائي للنمو المجاميع تاو. كل من هذه الأساليب له ايجابيات وسلبيات.
وأجرى رد فعل محطة الوضع مع فقط تلك المكونات اللازمة لتجميع تاو. تمييع وخلط رد فعل مع thioflavin T يبطئ بشكل كبير بانخفاض معدل الزيادة مضان، يتوقف أساسا على رد فعل لمضان الكمي. وبالتالي هذا الأسلوب هو أيضا متوافقة مع التشغيل اليدوي. ومع ذلك، لأن reactioيتم إنهاء ن عمليا على تى اتش تى بالإضافة إلى ذلك، كمية كبيرة من تاو قد يكون مطلوبا بتهمة التآمر منحنى التجميع. آخر التحذير المحتملة لهذا الأسلوب هو أن الوصول المتكرر إلى خليط التفاعل قد يعرض جرثومية أو تلوث بروتين، أو بروتين الأكسدة. في المقابل، فإن الوضع مع صبغ يسمح توليد اميلويد في حضور تى اتش تى أو تي إتش إس. النهوض التجميع يمكن رصدها بشكل مستمر دون أن يزعج رد فعل. هذه الميزة هي جاذبية خاصة عند إنشاء منصة الفحص الآلي. ومع ذلك، قد الأصباغ المختلفة تثير استجابات محددة. في الواقع، تى اتش تى يؤخر تاو وف تاو التجميع بشكل كبير، ولكن تي إتش إس له تأثيرات قليلا (مقارنة أرقام 3 و 5). هناك العديد من الأصباغ مضان أخرى، بما في ذلك الكونغو thiazines الأحمر والتي تم استخدامها في النسيجية وبيولوجيا الخلية دراسات لتشكيل PHF. وذكر تقرير واحد على الأقل أن بعض هذه الأصباغ ربما تتسبب في تجميع تاو في الأنسجةخلايا الثقافة 45. ولذلك، عند اختيار هذه المركبات لحركية amyloidogenesis الدراسات، ويجب أن تمارس بحذر وأن الأصباغ المختلفة قد يكون من الضروري مقارنة.
وفيما يتعلق باختيار الصك، والتألق عينة واحدة المستخدمة في النهج الأول هو موثوق بها للغاية، ولكن قد تكون عملية شاقة عندما تكون أكثر من بضعة ردود الفعل على أن تقارن. باستخدام cuvettes متعددة يمكن أن تساعد في تجنب التلوث المتبادل بين ردود الفعل، على الرغم من أن تكلفة هذه cuvettes الكوارتز الهشة قد تكون باهظة للبعض. على العكس من ذلك، يمكن للقارئ صفيحة ميكروسكوبية متعددة جيدا بفحص ردود فعل متعددة في نفس الوقت. استخدام المتاح لوحات 96-جيدا هو مفيد أيضا. مع وجود عنصر التدفئة، ويمكن للقارئ صفيحة ميكروسكوبية يكون جهاز مخصص لرصد ردود الفعل متعددة في 37 ° C لفترة طويلة من الزمن. ومع ذلك، تبخر قد يكون مصدرا للقلق. DiNitto وآخرون. مضافين رد فعل مماثل مع الزيوت المعدنية لمنعالتبخر 46.
بعض الاحتياطات التي يجب اتخاذها للالبروتوكولات المذكورة أعلاه لضمان تحقيق نتائج متسقة والكمية. أولا، تاو وف تاو تشكل بشكل عفوي المجاميع اميلويد على مر الزمن، وخاصة عندما تكون في تركيز عال. وبالتالي فمن الضروري تجميد جميع aliquots من محضرات البروتين، وذوبان الجليد فقط حجم اللازمة قبل التجارب. وحتى مع ذلك، بعض المجاميع، بما في ذلك سيطة الحبيبية اكتشافها من قبل تى اتش تى 47 قد شكلت خلال إعداد البروتينات المؤتلف. إن القراءة الأولية مضان كبيرة من رد فعل تجميع نموذجية هي بالتالي المشتركة. ومع ذلك، مضيفا خطوة ما قبل الغزل ونقل طاف لأنبوب منفصل، حتى بدون وجود البروتين بيليه مرئية، يمكن أن تقلل، والحفاظ على ثابت، مضان الأولي من نفس دفعة من تاو وف تاو الإعدادية. ثانيا، حل الأسهم عمل تى اتش تى (أي 60 ميكرومتر) هو مستقر في RT لأقل من أسبوع قبل عشريقلل البريد مضان. وهكذا ينصح لإعادة جعل 60 ميكرومتر تى اتش تى كل عدة أيام.
أحد الأسباب الكامنة وراء بارز دراسات ف تاو تجميع هو تطوير وسائل التشخيص AD جديدة والعلاجات. وقد تم تحديد المركبات التي تمنع أو العودة المؤتلف تاو تجميع من على شاشات عالية الإنتاجية والاختبارات المستهدفة 18،40،41،48. تبقى فعالية هذه المركبات لف تاو التجميع إلى توضيح. ونفذت هذه الشاشات في وضع المحطة من قبل الاستغناء مزيج التجميع المشترك دون صبغ إلى الآبار متعدد الطبقات الفردية مع مركبات مختلفة. عادة بعد O / N الحضانة، ويضاف تى اتش تى أو تي إتش إس لقياس مضان، وكشف عن السلطة المثبطة للعديد من المركبات. النهج مع صبغ المذكورة أعلاه والتي رانكين وآخرون. 49 لم يتم بعد إدراجها إلى شاشات عالية الإنتاجية. الآن مع تاو hyperphosphorylated المتاحة للدراسات الحركية والصيدلية لتقرنتشكيل خيوط حلزونية، من المرجح أن تقدم أبعد اكتشاف المخدرات مرض الزهايمر.
وأخيرا، تجدر الإشارة إلى أن دراسة ف تاو التجميع ذات أهمية حاسمة لtauopathies فحسب، بل يمكن أيضا أن تؤثر على السكان أوسع. على سبيل المثال، هناك تقارير تفيد بأن التشابك الليفي العصبي يتم كشفها في بعض المرضى من التهاب الدماغ الصدمة المزمنة، مثل لاعبي كرة القدم الأميركي المهنية والملاكمين 50-52. كما تم الإبلاغ عن علاقة مماثلة لصدمة مرضى إصابات الدماغ واحد أو المتكررة بينهم جنود 53. وبالتالي قد البروتوكولات المذكورة في هذا العمل يساعد على اكتشاف وتطوير علاجات جديدة تستهدف المجاميع ف تاو في الخلايا العصبية.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Trizma base | Sigma | T1503 | |
| NaCl | Macron Fine Chemicals | MAL-7581-06 | |
| Ethylenediaminetetraacetic Acid (EDTA) | Invitrogen | 15576-028 | |
| Thioflavin T | Sigma | T3516 | Stored in dark |
| Thioflavin S | Sigma | T1892 | Stored in dark |
| heparin | Sigma | H3393 | |
| DL-Dithiothreitol (DTT) | Sigma | D9779 | Stored at 4 °C |
| 96-well plate | Corning | 3917 | |
| ISA SPEX FluoroMax-2 | Horiba | ||
| SpectraMax M2 Multi-Mode Microlate Reader | Molecular Devices | ||
| Mouse Anti-Tau Monoclonal Antibody | R&D Systems | MAB3494 | Stored at –80 °C |
References
- Lee, V. M., Goedert, M., Trojanowski, J. Q.
- Ballatore, C., Lee, V. M., Trojanowski, J. Q. Tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nat Rev Neurosci. 8 (9), 663-672 (2007).
- Arriagada, P. V., Marzloff, K., Hyman, B. T. Distribution of Alzheimer-type pathologic changes in nondemented elderly individuals matches the pattern in Alzheimer's disease. Neurology. 42 (9), 1681-1688 (1992).
- Arriagada, P. V., Growdon, J. H., Hedley-Whyte, E. T., Hyman, B. T. Neurofibrillary tangles but not senile plaques parallel duration and severity of Alzheimer's disease. Neurology. 42 (3 Pt 1), 631-639 (1992).
- Bancher, C., Braak, H., Fischer, P., Jellinger, K. A. Neuropathological staging of Alzheimer lesions and intellectual status in Alzheimer's and Parkinson's disease patients. Neurosci Lett. 162 (1-2), 179-182 (1993).
- Guillozet, A. L., Weintraub, S., Mash, D. C., Mesulam, M. M. Neurofibrillary tangles, amyloid, and memory in aging and mild cognitive impairment. Arch Neurol. 60 (5), 729-736 (2003).
- Hasegawa, M., et al. Protein sequence and mass spectrometric analyses of tau in the Alzheimer's disease brain. J Biol Chem. 267 (24), 17047-17054 (1992).
- Matsuo, E. S., et al. Biopsy-derived adult human brain tau is phosphorylated at many of the same sites as Alzheimer's disease paired helical filament tau. Neuron. 13 (4), 989-1002 (1994).
- Bamburg, J. R., Bloom, G. S.
- Denk, F., Wade-Martins, R. Knock-out and transgenic mouse models of tauopathies. Neurobiol Aging. 30 (1), 1-13 (2009).
- Gong, C. X., Iqbal, K. Hyperphosphorylation of microtubule-associated protein tau: a promising therapeutic target for Alzheimer disease. Curr Med Chem. 15 (23), 2321-2328 (2008).
- Mazanetz, M. P., Fischer, P. M. Untangling tau hyperphosphorylation in drug design for neurodegenerative diseases. Nat Rev Drug Discov. 6 (6), 464-479 (2007).
- Brunden, K. R., Trojanowski, J. Q., Lee, V. M. Advances in tau-focused drug discovery for Alzheimer's disease and related tauopathies. Nat Rev Drug Discov. 8 (10), 783-793 (2009).
- Querfurth, H. W., LaFerla, F. M.
- Buee, L., Bussiere, T., Buee-Scherrer, V., Delacourte, A., Hof, P. R. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res Brain Res Rev. 33 (1), 95-130 (2000).
- Lee, V. M., Brunden, K. R., Hutton, M., Trojanowski, J. Q. Developing therapeutic approaches to tau, selected kinases, and related neuronal protein targets. Cold Spring Harb Perspect Med. 1 (1), 006437 (2011).
- Mandelkow, E. M., Mandelkow, E. Biochemistry and cell biology of tau protein in neurofibrillary degeneration. Cold Spring Harb Perspect Med. 2 (7), a006247 (2012).
- Bulic, B., Pickhardt, M., Mandelkow, E. Progress and Developments in Tau Aggregation Inhibitors for Alzheimer Disease. J Med Chem. 56 (11), 4135-4155 (2013).
- Cowan, C. M., Quraishe, S., Mudher, A. What is the pathological significance of tau oligomers. Biochem Soc Trans. 40 (4), 693-697 (2012).
- Spires-Jones, T. L., Kopeikina, K. J., Koffie, R. M., de Calignon, A., Hyman, B. T. Are tangles as toxic as they look. J Mol Neurosci. 45 (3), 438-444 (2011).
- SantaCruz, K., et al. Tau suppression in a neurodegenerative mouse model improves memory function. Science. 309 (5733), 476-481 (2005).
- Wittmann, C. W., et al. Tauopathy in Drosophila: neurodegeneration without neurofibrillary tangles. Science. 293 (5530), 711-714 (2001).
- Shulman, J. M., Feany, M. B.
- Wille, H., Drewes, G., Biernat, J., Mandelkow, E. M., Mandelkow, E. Alzheimer-like paired helical filaments and antiparallel dimers formed from microtubule-associated protein tau in vitro. J Cell Biol. 118 (3), 573-584 (1992).
- Alonso, A., Zaidi, T., Novak, M., Grundke-Iqbal, I., Iqbal, K. Hyperphosphorylation induces self-assembly of tau into tangles of paired helical filaments/straight filaments. Proc Natl Acad Sci U S A. 98 (12), 6923-6928 (2001).
- Barghorn, S., Biernat, J., Mandelkow, E. Purification of recombinant tau protein and preparation of Alzheimer-paired helical filaments in vitro. Methods Mol Biol. 299, 35-51 (2005).
- Wilson, D. M., Binder, L. I. Polymerization of microtubule-associated protein tau under near-physiological conditions. J Biol Chem. 270 (41), 24306-24314 (1995).
- Wilson, D. M., Binder, L. I. Free fatty acids stimulate the polymerization of tau and amyloid beta peptides. In vitro evidence for a common effector of pathogenesis in Alzheimer's disease. Am J Pathol. 150 (6), 2181-2195 (1997).
- Perez, M., Valpuesta, J. M., Medina, M., Montejo de Garcini, E., Avila, J. Polymerization of tau into filaments in the presence of heparin: the minimal sequence required for tau-tau interaction. J Neurochem. 67 (3), 1183-1190 (1996).
- Carlson, S. W., et al. A complex mechanism for inducer mediated tau polymerization. Biochemistry. 46 (30), 8838-8849 (2007).
- Goedert, M., et al. Assembly of microtubule-associated protein tau into Alzheimer-like filaments induced by sulphated glycosaminoglycans. Nature. 383 (6600), 550-553 (1996).
- King, M. E., Gamblin, T. C., Kuret, J., Binder, L. I. Differential assembly of human tau isoforms in the presence of arachidonic acid. J Neurochem. 74 (4), 1749-1757 (2000).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pseudo-phosphorylation of tau at Ser202 and Thr205 affects tau filament formation. Brain Res Mol Brain Res. 138 (1), 84-93 (2005).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pre-assembled tau filaments phosphorylated by GSK-3b form large tangle-like structures. Neurobiol Dis. 31 (3), 368-377 (2008).
- Grundke-Iqbal, I., et al. Abnormal phosphorylation of the microtubule-associated protein tau (tau) in Alzheimer cytoskeletal pathology. Proc Natl Acad Sci U S A. 83 (13), 4913-4917 (1986).
- Castellani, R. J., Perry, G. Pathogenesis and disease-modifying therapy in Alzheimer's disease: the flat line of progress. Arch Med Res. 43 (8), 694-698 (2012).
- Green, R. C., et al. Effect of tarenflurbil on cognitive decline and activities of daily living in patients with mild Alzheimer disease: a randomized controlled trial. JAMA. 302 (23), 2557-2564 (2009).
- Gauthier, S., et al. Effect of tramiprosate in patients with mild-to-moderate Alzheimer's disease: exploratory analyses of the MRI sub-group of the Alphase study. J Nutr Health Aging. 13 (6), 550-557 (2009).
- Pickhardt, M., et al. Anthraquinones inhibit tau aggregation and dissolve Alzheimer's paired helical filaments in vitro and in cells. J Biol Chem. 280 (5), 3628-3635 (2005).
- Crowe, A., Ballatore, C., Hyde, E., Trojanowski, J. Q., Lee, V. M. High throughput screening for small molecule inhibitors of heparin-induced tau fibril formation. Biochem Biophys Res Commun. 358 (1), 1-6 (2007).
- Taniguchi, S., et al. Inhibition of heparin-induced tau filament formation by phenothiazines, polyphenols, and porphyrins. J Biol Chem. 280 (9), 7614-7623 (2005).
- Sigurdsson, E. M. Tau-focused immunotherapy for Alzheimer's disease and related tauopathies. Curr Alzheimer Res. 6 (5), 446-450 (2009).
- Tan, Y. J., et al. Phosphopeptide Enrichment with TiO-Modified Membranes and Investigation of Tau Protein Phosphorylation. Anal Chem. 85 (12), 5699-5706 (2013).
- Santa-Maria, I., Perez, M., Hernandez, F., Avila, J., Moreno, F. J. Characteristics of the binding of thioflavin S to tau paired helical filaments. J Alzheimers Dis. 9 (3), 279-285 (2006).
- Lira-De Leon, K. I., et al. Molecular mechanism of tau aggregation induced by anionic and cationic dyes. J Alzheimers Dis. 35 (2), 319-334 (2013).
- DiNitto, J. P., Wang, L., Wu, J. C. Continuous fluorescence-based method for assessing dicer cleavage efficiency reveals 3' overhang nucleotide preference. BioTechniques. 48, 303-311 (2010).
- Maeda, S., et al. Granular tau oligomers as intermediates of tau filaments. Biochemistry. 46 (12), 3856-3861 (2007).
- Pickhardt, M., et al. Phenylthiazolyl-hydrazide and its derivatives are potent inhibitors of tau aggregation and toxicity in vitro and in cells. Biochemistry. 46 (35), 10016-10023 (2007).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Tau phosphorylation by GSK-3beta promotes tangle-like filament morphology. Mol Neurodegener. 2, 12 (2007).
- McKee, A. C., et al. Chronic traumatic encephalopathy in athletes: progressive tauopathy after repetitive head injury. J Neuropathol Exp Neurol. 68 (7), 709-735 (2009).
- Herrup, K. Reimagining Alzheimer's disease--an age-based hypothesis. J Neurosci. 30 (50), 16755-16762 (2010).
- Gavett, B. E., Stern, R. A., McKee, A. C. Chronic traumatic encephalopathy: a potential late effect of sport-related concussive and subconcussive head trauma. Clin Sports Med. 30 (1), 179-188 (2011).
- Tsitsopoulos, P. P., Marklund, N. Amyloid-beta Peptides and Tau Protein as Biomarkers in Cerebrospinal and Interstitial Fluid Following Traumatic Brain Injury: A Review of Experimental and Clinical Studies. Front Neurol. 4, 79 (2013).
