

Summary
Представлено и обсуждено формирование пучков актомиозина in vitro и измерение генерации силы ансамбля миозина с помощью оптического пинцета.
Abstract
Миозины являются моторными белками, которые гидролизуют АТФ, чтобы шагнуть по следам актиновой нити (ФП) и необходимы в клеточных процессах, таких как подвижность и сокращение мышц. Чтобы понять их силовые механизмы, миозин II был исследован как на уровне одной молекулы (SM), так и в виде команд двигателей in vitro с использованием биофизических методов, таких как оптический захват.
Эти исследования показали, что поведение миозина, генерирующее силу, может сильно отличаться при переходе от уровня одной молекулы в трехшарном расположении к группам двигателей, работающих вместе на жесткой бусине или поверхности покрова в скользящем расположении. Однако эти конструкции анализа не позволяют оценить групповую динамику миозина в вязкоупругой структурной иерархии, как это было бы в клетке. Мы разработали метод с использованием оптического пинцета для исследования механики генерации силы миозиновыми ансамблями, взаимодействующими с несколькими актиновыми нитями.
Эти пучки актомиозина облегчают исследование в иерархической и совместимой среде, которая захватывает двигательную связь и выход ансамблевой силы. Настраиваемый характер анализа позволяет изменять условия эксперимента, чтобы понять, как модификации ансамбля миозина, пучка актиновой нити или окружающей среды приводят к различным силовым выходам.
Introduction
Моторные белки необходимы для жизни, преобразуя химическую энергию в механическую работу 1,2,3. Миозиновые моторы взаимодействуют с актиновыми нитями, делая шаги вдоль нитей, похожие на дорожку, а динамика актин-миозиновых сетей осуществляет сокращение мышц, подвижность клеток, сократительное кольцо во время цитокинеза и движение груза внутри клетки, среди других важных задач 3,4,5,6,7,8 . Поскольку миозины играют так много важных ролей, сбой в функциональности миозин-актиновой сети может привести к развитию заболеваний, таких как мутации в тяжелой цепи миозина, которые вызывают гиперконтрактность сердца при гипертрофической кардиомиопатии (ГКМП)9,10,11,12,13,14 . При мышечном сокращении отдельные миозиновые моторы взаимодействуют друг с другом, работая как ансамбль, чтобы обеспечить необходимую механическую энергию, которая осуществляет относительное скольжение AFs 4,15,16,17,18. Миозиновые двигатели образуют поперечные мосты между АФ и используют конформационные изменения из-за его механохимического цикла, чтобы коллективно двигаться к колючему концу выровненных нитей 17,18,19,20,21.
Разработка количественных анализов подвижности in vitro на уровне SM с использованием таких методов, как оптический захват, способствовала сбору беспрецедентных деталей о том, как функционируют отдельные двигатели миозина, включая измерение генерации силы SM и размеров шагов 22,23,24,25,26,27,28,29,30 . Finer et al. разработали оптический анализ улавливания «три шарика» или «гантели» для исследования механики генерации силы одиночных двигателей миозина II23,31. Поскольку мышечный миозин II работает в командах для сокращения АФ, но не является процессивным на уровне SM, ориентация оптического улавливающего анализа должна быть перестроена из классического подхода32 с моторной бисерой. Для формирования анализа гантелей использовали две оптические ловушки для удержания AF над миозиновым двигателем, связанным с шариком, прикрепленным к крышке, и выходная сила одного двигателя измерялась через движения AF внутри ловушки23.
Однако силы СМ и использование ориентации анализа на один двигатель / одну нить не дают полного представления о генерации силы на системном уровне, поскольку многие моторные белки, включая миозин II, не работают изолированно и часто не функционируют как сумма их частей 15,16,17,32,33,34,35,36 . Более сложные структуры, включающие более одного мотора, взаимодействующего с более чем одной нитью, необходимы для лучшего понимания синергизма сетей миозиновых и актиновых нитей15,32. Ориентация анализа гантелей была использована для исследования генерации силы небольшого ансамбля путем прикрепления нескольких миозинов к бусину или использования нити толщиной миозина, прикрепленной к поверхности и позволяющей двигателям взаимодействовать с взвешенной AF 4,23,34,37,38,39,40.
Другие небольшие ансамблевые анализы включают в себя скользящий анализ нити in vitro, в котором двигатели миозина наносятся на поверхность покрова, а шарик, связанный с AF, используется для зондирования силы, генерируемой командой двигателей 4,35,36,38,39,40,41,42,43 . В обоих этих случаях миозины связаны с жесткой поверхностью — бусиной или крышкой — и используют одну ФП. В этих случаях двигатели не могут свободно двигаться или общаться друг с другом, и наличие жестко связанных миозинов не отражает соответствующую, иерархическую среду, в которой двигатели будут работать вместе в саркомере32. Предыдущие исследования показали, что миозин II может ощущать окружающую среду и соответствующим образом адаптироваться к изменяющимся условиям вязкоупругой или двигательной концентрации путем изменения таких характеристик, как генерация силы и коэффициент полезности 41,44,45. Таким образом, необходимо разработать оптический анализ улавливания, который способствует и фиксирует моторную связь и соответствие системы, чтобы нарисовать более реалистичную картину механистических основ генерации сил ансамбля миозина II.
Здесь мы разработали метод сопряжения иерархической структуры in vitro с оптическим захватом путем формирования пучков актимозина или сэндвичей, состоящих из нескольких миозиновых двигателей, взаимодействующих между двумя актиновыми нитями. Эта модульная геометрия анализа имеет возможность непосредственно исследовать, как молекулярные и экологические факторы влияют на генерацию ансамблевой силы миозина. Кроме того, исследование механизмов генерации силы через эти актин-миозиновые ансамбли может помочь в моделировании и понимании того, как крупномасштабные клеточные задачи, такие как сокращение мышц, распространяются вверх с молекулярного уровня 9,10,13.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Травление обложек
- Растворите 100 г KOH в 300 мл 100% этанола в стакане объемом 1000 мл. Перемешивайте с перемешиванием до тех пор, пока большая часть KOH не растворится.
ВНИМАНИЕ: Концентрированный раствор KOH может вызвать ожоги и повреждение одежды. Наденьте перчатки, защиту глаз и лабораторное пальто. - Поместите крышки по отдельности в стойки для чистки чехлов.
ПРИМЕЧАНИЕ: Стойки спроектированы с прорезями, которые удерживают отдельные крышки, расположенные друг от друга, чтобы обеспечить травление и промывку на каждой стороне крышки, дренажные отверстия в нижней части и изготовлены из материала, который может выдерживать суровые условия травления. Они могут быть изготовлены на заказ или приобретены на коммерческой основе. - Подготовьте и промаркируйте три стакана по 1000 мл: один с 300 мл этанола и два стакана с 300 мл воды обратного осмоса (RO).
ПРИМЕЧАНИЕ: Здесь вода RO была получена из лабораторного очистителя воды, но ее также можно было приобрести на коммерческой основе, если местный очиститель недоступен. - Поместите каждый из четырех стаканов в ультразвуковой аппарат для ванны, чтобы дегазировать в течение 5 минут.
- Погрузите стойку с крышками в стакан KOH и этанола и ультразвука на 5 мин.
- Перенесите стойку с крышками из стакана KOH/этанола в стакан только для этанола. Окуните стойку вверх и вниз в стакан до тех пор, пока не останется бисероплетения.
ПРИМЕЧАНИЕ: Позаботьтесь о том, чтобы не потревожить крышки и не уронить стойку в стакан. Это приведет к тому, что крышки выйдут из стойки или вызовут химическое разбрызгивание. - Аккуратно перенесите стойку с крышками с этанолового стакана на стакан с водой, опустив вверх и вниз до тех пор, пока не останется бисероплетения.
- Погрузите стойку с крышками в стакан с водой, которая еще не использовалась, и снова обработайте ультразвуком в течение 5 минут.
- Используйте бутылку, чтобы опрыскать стойку с крышками водой, пока она не сойдет с обложек гладко. Повторить с этанолом.
- Поместите стеллажи сушиться в духовку при температуре 90 °C в течение 20 мин. Храните стеллажи с травлеными крышками при комнатной температуре в закрытых контейнерах, чтобы предотвратить загрязнение перед использованием.
2. Полимеризация актиновой нити
- Сделать решение T
- В коническую пробирку объемом 50 мл добавить 3,94 г Tris-HCl и 0,147 г CaCl2. Добавьте воду обратного осмоса, чтобы получить общий объем 50 мл и хорошо перемешать.
ПРИМЕЧАНИЕ: Конечные концентрации раствора Т составляют соответственно 500 мМ Tris-HCl и 20 мМCaCl2 . - Маркируйте тюбик раствором T и храните его при температуре 4 °C.
- В коническую пробирку объемом 50 мл добавить 3,94 г Tris-HCl и 0,147 г CaCl2. Добавьте воду обратного осмоса, чтобы получить общий объем 50 мл и хорошо перемешать.
- Создать буфер TC
- Смешайте 40 мл воды обратного осмоса и 1,5 мл раствора Т в конической трубке объемом 50 мл. Измените рН до 8,0, добавив небольшое количество концентрированного КОН. Добавьте воду, чтобы получить 50 мл раствора, и проверьте рН. При необходимости отрегулируйте pH.
ПРИМЕЧАНИЕ: Конечный буфер TC содержит 5 мМ Tris-HCl и 0,2 мМCaCl2 при рН 8. - Нанесите маркировку на трубку TC и храните ее при температуре 4 °C.
- Смешайте 40 мл воды обратного осмоса и 1,5 мл раствора Т в конической трубке объемом 50 мл. Измените рН до 8,0, добавив небольшое количество концентрированного КОН. Добавьте воду, чтобы получить 50 мл раствора, и проверьте рН. При необходимости отрегулируйте pH.
- Создание буфера FC
- Добавьте 85 мл воды обратного осмоса, 10 мл раствора T, 3,73 г KCl и 0,041 г MgCl2 в буферную бутылку объемом 100 мл. Модифицируйте рН до 7,5 путем добавления небольших объемов концентрированного КОН. Добавьте воду, чтобы получить окончательный объем 100 мл и проверьте рН.
ПРИМЕЧАНИЕ: Конечный буфер FC содержит 500 мМ Tris-HCl, 500 мМ KCl, 2 мМ MgCl2 и 2 мМ CaCl2 при рН 7,5. - Маркируйте трубку FC и храните ее при температуре 4 °C.
- Добавьте 85 мл воды обратного осмоса, 10 мл раствора T, 3,73 г KCl и 0,041 г MgCl2 в буферную бутылку объемом 100 мл. Модифицируйте рН до 7,5 путем добавления небольших объемов концентрированного КОН. Добавьте воду, чтобы получить окончательный объем 100 мл и проверьте рН.
- Подготовьте общий буфер актина (GAB).
- Смешайте 485 мкл буфера TC, 10 мкл 10 мМ АТФ и 5 мкл 50 мМ DTT в микроцентрифужной трубке.
ПРИМЕЧАНИЕ: Конечные буферные условия составляют 5 мМ Tris-HCl, 0,2 мМ CaCl2, 0,5 мМ DTT и 0,2 мМ ATP. - Пометьте его как GAB и храните при температуре 4 °C.
- Смешайте 485 мкл буфера TC, 10 мкл 10 мМ АТФ и 5 мкл 50 мМ DTT в микроцентрифужной трубке.
- Приготовьте буфер полимеризации актина (APB).
- Смешайте 455 мкл fc-буфера, 25 мкл 100 мМ АТФ и 20 мкл 50 мМ DTT в микроцентрифужной трубке.
ПРИМЕЧАНИЕ: Конечные буферные условия составляют 50 мМ Tris-HCl, 500 мМ KCl, 2 мМ MgCl2, 2 мМ CaCl2 2 мМ DTT и 5 мМ ATP. - Пометьте трубку как APB и храните ее при температуре 4 °C.
- Смешайте 455 мкл fc-буфера, 25 мкл 100 мМ АТФ и 20 мкл 50 мМ DTT в микроцентрифужной трубке.
- Восстановление актина
- Восстановите актин скелетных мышц кролика, добавив 100 мкл деионизированной воды во флакон 1 мг лиофилизированного актина. Хорошо перемешать, аккуратно пипетировав вверх и вниз. Аликвота в образцы 5 мкл, заморозьте и храните актиновые аликвоты 10 мг/мл при -80 °C.
- Восстановите биотинилированный актин скелетных мышц кролика, добавив 20 мкл воды обратного осмоса. Аликвота в образцах 5 мкл, заморозьте и храните 1 мг/мл биотинилированных актиновых аликвот при -80 °C.
- Полимеризация немаркированного актина со стабилизацией родамина фаллоидина
- Разморозьте один флакон с актином 10 мг/мл и держите его на льду.
- Приготовьте свежий ГАМ-буфер, добавьте 100 мкл ГАБ в актин-аликвоту и перемешайте, аккуратно пипетируя вверх и вниз. Инкубировать раствор на льду в течение 1 ч.
- Приготовьте свежий APB во время инкубации. После инкубации полимеризуют актин в нити, добавляя 11 мкл APB к раствору актина. Хорошо перемешать, аккуратно пипетировав вверх и вниз. Выложить на лед на 20 мин.
- Добавьте 5 мкл меченого родамином фаллоидина в свежеполимеризованный раствор актиновой нити. Оставить на льду в темноте на 1 ч.
- Храните флакон с родамином актином, завернутый в алюминиевую фольгу, в темноте при температуре 4 °C.
ПРИМЕЧАНИЕ: Рекомендуется использовать эти нити в течение максимум 1 недели. Качество ФП может быть подтверждено каждый день с помощью быстрой визуализации проточной ячейки, содержащей только АФ, и просмотра последовательных нитей изо дня в день.
- Полимеризация биотинилированного актина со стабилизацией фаллоидина Alexa Fluor 488
- Разморозьте один флакон 10 мг/мл актина и 1 флакон 1 мг/мл биотинилированного актина и держите их на льду.
- Сделайте свежий GAB-буфер.
- Соедините два флакона (этап 2.8.1) в соотношении актин:биотинилированный актин 10:1. Добавьте 100 мкл GAB в смесь актинов и хорошо перемешайте, осторожно пипетируя вверх и вниз. Инкубировать на льду в течение 1 ч.
- Делайте свежий APB во время инкубации.
- После стадии инкубации полимеризуют актин, добавляя 11 мкл APB к раствору актина. Хорошо перемешайте, аккуратно пипетируя вверх и вниз. Инкубировать на льду в течение 20 мин.
- Добавьте 5 мкл меченого фаллоидина Alexa Fluor 488 и инкубируйте на льду в темноте в течение 1 ч.
- Храните биотинилированный флакон актина, завернутый в алюминиевую фольгу, в темноте при температуре 4 °C.
ПРИМЕЧАНИЕ: Эти нити можно использовать в течение максимум 1 недели.
3. Подготовка миозина и бисера
- Восстановление миозина II
- Ненадолго вращают вниз (~5 с) лиофилизированный скелетный миозин II, чтобы собрать его на дне трубки с помощью стандартной миницентрифуги.
- Восстановите миозин до 10 мг/мл, добавив 100 мкл 1 мМ DTT, приготовленного в воде обратного осмоса.
- Разбавить запас раствора миозина в 10 раз, добавив 10 мкл 10 мг/мл миозина к 90 мкл 1 мМ DTT в воде обратного осмоса. Сделайте небольшие (1-5 мкл) аликвоты, заморозьте и храните при -80 °C.
ПРИМЕЧАНИЕ: Активность миозина может быть подтверждена путем выполнения стандартного скользящего анализа нити, опубликованного ранее46,47. Краткое описание см. в обсуждении.
- Чистящие бусины со стрептавидиновым покрытием
- Разбавьте 20 мкл шариков стрептавидина 1 мкм в 80 мкл воды обратного осмоса. Промыть четыре раза, раскручивая вниз при 9 600 × г и восстанавливая в 100 мкл воды обратного осмоса.
- Хранить ультразвуком в течение 2 мин при амплитуде 40% и хранить промытые шарики на ротаторе при 4 °C.
4. Подготовка проточных клеток
- Приготовьте раствор поли-l-лизина (ФАПЧ), добавив 30 мл 100% этанола в пробирку объемом 50 мл и добавив 200 мкл 0,1% мас./об.поли-l-лизина в воду и хорошо перемешайте.
- Добавьте в раствор ФАПЧ травленую крышку и дайте ей впитаться в течение 15 минут. Снимите крышку пинцетом, позаботившись о том, чтобы коснуться только края крышки, когда она вытягивается из трубки (см. Рисунок 1A-C). Захватите чехлы по краям рукой в перчатке.
- Высушите крышку фильтрованной авиакомпанией до тех пор, пока не останется этанола и остатков на крышке.
- Нанесите два куска двусторонней липкой ленты на середину слайда микроскопа, на расстоянии 3-4 мм друг от друга. Оторвите или отрежьте лишнюю ленту, которая свисает с края горки.
- Добавьте крышку с фапч-покрытием поверх ленты перпендикулярно длинной оси слайда микроскопа (образуя Т), чтобы сформировать канал.
- Используйте небольшую трубку, чтобы плотно сжать крышку на ленту и микроскоп, пока лента не станет прозрачной (рисунок 1A). Убедитесь, что в ленте нет пузырьков, так как это может привести к утечке из канала потока.
ПРИМЕЧАНИЕ: Проточная ячейка может удерживать объем 10-15 мкл.
5. Препарат пучка актомиозина
- В отдельных пробирках разводят каждый тип актиновой нити (родамин- и биотинилированный 488-меченый) 600x путем смешивания 0,5 мкл соответствующего, меченного актина с 300 мкл APB. Добавьте дополнительно 5 мкл соответственно меченого фаллоидина в каждый тюбик и инкубируйте на льду в темноте в течение 15 минут.
- К биотинилированному раствору актина добавляют систему очистки кислорода из 1 мкл бета-D-глюкозы при 500 мг/мл, 1 мкл глюкозооксидазы при 25 мг/мл и 1 мкл каталазы при 500 ед/мл. Добавьте 1 мкл 100 мМ АТФ и 1 мкл 100 разбавленных, очищенных шариков стрептавидина. Аккуратно перемешайте кончиком пипетки. Поместите суспензию на ротатор при 4 °C, пока собирается остальная часть пучка актомиозина.
- Добавьте 15 мкл разбавленного актаина родамина в проточную ячейку ФАПЧ (рисунок 1D). Пропустите лишний раствор через проточную ячейку, но не дайте каналу потока высохнуть. Инкубировать в течение 10 мин в камере влажности.
ПРИМЕЧАНИЕ: Камеры влажности могут быть изготовлены из пустых коробок с наконечниками пипеток с водой, добавленной на дно, и крышкой, покрытой алюминиевой фольгой, чтобы блокировать свет. - Приготовьте раствор казеина 1 мг/мл в APB.
- Добавьте 15 мкл 1 мг/мл казеина для предотвращения неспецифического связывания последующих компонентов (рисунок 1Е). Инкубировать в течение 5 мин в камере влажности.
- Добавьте желаемую концентрацию миозина к биотинилированному актину и суспензии шарика со стадии 5.2. Осторожно перемешайте кончиком пипетки, а затем сразу же добавьте 15 мкл суспензии стадии 5,2 + желаемую концентрацию миозина в проточную клетку (рисунок 1F,G). Инкубировать в течение 20 мин. Запечатайте открытые концы проточной ячейки лаком для ногтей, чтобы предотвратить испарение во время экспериментов по визуализации и оптическому улавливанию.
ПРИМЕЧАНИЕ: Концентрация раствора миозина 1 мкМ дает надежную комплектацию и может быть использована в качестве отправной точки для желаемой настройки анализа (см. Рисунок 2).
6. Измерение силы с помощью оптической ловушки (NT2 Nanotracker2)
ПРИМЕЧАНИЕ: Хотя приведенный ниже протокол предназначен специально для системы NT2, этот анализ может использоваться с другими оптическими системами улавливания, включая те, которые изготовлены специально, которые также имеют возможности флуоресценции. Общий рабочий процесс остается прежним: получение поверхности слайда в фокусе, выполнение калибровки шариков и получение данных путем поиска флуоресцентных пучков актина. Для системы NT2 дополнительный рисунок S1, дополнительный рисунок S2, дополнительный рисунок S3, дополнительный рисунок S4, дополнительный рисунок S5, дополнительный рисунок S6 и дополнительный рисунок S7 содержат подробную информацию о системе оптического захвата и программном интерфейсе.
- Включите блок управления и лазер (дополнительный рисунок S1).
- Запустите программное обеспечение компьютера с оптической ловушкой, щелкнув значок JPK Nanotracker на рабочем столе.
- Разбудите пульт дистанционного управления, нажав кнопку Logitech в центре (дополнительный рисунок S2).
- Включите флуоресцентный модуль, включив/выключив переключатель (дополнительный рисунок S3).
- Поверните башню куба фильтра для получения изображений яркого поля (дополнительный рисунок S4).
- Как только система будет готова, включите лазер с помощью кнопки Laser Power в левом нижнем углу экрана до 50 мВт и дайте ему стабилизироваться в течение 30 минут (дополнительный рисунок S5).
- Последовательно нажимайте кнопки «Освещение», «Камера», «Объектив» и «Сценическое движение» в программном обеспечении, чтобы открыть эти окна для просмотра и манипуляции во время эксперимента. Включите подсветку микроскопа, нажав на кнопку On/Off и установив ее на максимальную мощность, щелкнув и перетащив полосу вправо (дополнительный рисунок S5).
- Откройте область образца и извлеките держатель образца из сцены микроскопа. Добавьте проточную ячейку, закрепите ее металлическими держателями для образцов и убедитесь, что слайд с крышкой находится внизу.
- Добавьте 30 мкл воды обратного осмоса в центр нижнего объектива. Не позволяйте кончику пипетки касаться объектива. Повторно вставьте стадию выборки.
ПРИМЕЧАНИЕ: Поскольку система NT2 использует цель погружения в воду в качестве цели улавливания, иммерсионная среда может отличаться в зависимости от цели захвата в настройках пользователя. - Поднимите нижнюю цель с помощью экранных стрелок управления или L2 на пульте дистанционного управления до тех пор, пока шарик воды не коснется крышки (дополнительный рисунок S5).
- Опустите верхнюю цель до тех пор, пока не будет достигнуто около половины расстояния до проточной ячейки с помощью экранных стрелок или R2 на пульте дистанционного управления. Добавьте 170 мкл воды обратного осмоса в верхнюю часть проточной ячейки непосредственно под верхним объективом. Опустите верхнюю цель до тех пор, пока она не нарушит поверхностное натяжение воды и не образует мениск.
- Перемещайте ступень микроскопа с помощью стрелки на пульте дистанционного управления до тех пор, пока не будет достигнут край ленты, прилегающий к каналу потока. Закройте дверцу для образцов.
ПРИМЕЧАНИЕ: "Щелчок" при закрытии дверцы образца указывает на то, что лазерный затвор теперь открыт. Это функция безопасности, которая позволяет затвору открываться только в том случае, если дверь закрыта. - Используя окно «Объектив » на экране, переведите край ленты в фокус, подняв нижний объект с именем Laser Objective вверх, щелкнув верхнюю стрелку с помощью экранных элементов управления. Проделайте то же самое для верхней цели, щелкнув стрелку внизу (дополнительный рисунок S5).
ПРИМЕЧАНИЕ: Двойные стрелки перемещают цель или сцену быстрее. Край ленты используется для фокусировки, потому что это большой, легко обнаруживаемый объект, который находится близко к поверхности крышки. Пузырьки воздуха внутри ленты являются еще одним вариантом. Однако это не требуется, если у пользователя есть автоматизированная процедура для поиска фокуса поверхности или предпочтительный внутренний метод. - Как только лента окажется в фокусе, частично закройте радужную оболочку в верхней части оптической ловушки. Опустите верхнюю цель вниз до тех пор, пока не будет видна полигональная форма радужной оболочки. Поместите эти края в фокус, снова откройте радужную оболочку, а затем соедините цели, щелкнув значок замка (дополнительный рисунок S5).
- Найдите плавающую бусину и поймайте ее, нажав на кнопку Trap Shutter , которая откроет затвор и позволит лазеру захвата ударить по образцу. Нажмите на курсор Trap на экране и перетащите его, чтобы переместить местоположение лазера треппинга. После захвата откалибруйте шарик, чтобы соотнести измерения напряжения с силой и смещением.
- Нажмите на кнопку Калибровка . Отрегулируйте процедуру калибровки на основе анализа спектров мощности и установите угловую частоту в программном обеспечении для направлений X, Y и Z (дополнительный рисунок S6).
- Нажмите Настройки. Введите диаметр шарика (1000 нм) и введите температуру ступени, расположенную в левом нижнем углу окна программного обеспечения. (см. дополнительный рисунок S6).
- Нажмите на Ловушку 1. Нажмите на X Сигнал. Нажмите « Выполнить», чтобы выполнить угловую подгонку частоты. Щелкните и перетащите в окно, чтобы оптимизировать подгонку функции. Нажмите « Использовать его» для значений чувствительности и жесткости. Нажмите Принять значения. Повторите эти действия для сигналов Y и Z. Закройте окно. (см. дополнительный рисунок S6).
ПРИМЕЧАНИЕ: Процедуры калибровки шариков на других оптических системах улавливания или специально изготовленных системах, которые были надежно протестированы пользователем, таких как метод равнораспределения или метод силы сопротивления, также приемлемы57,58. - Найдите пучок актомиозина, выполнив поиск бусин, связанных с АФ на поверхности покровного листа.
- Когда обнаружена бусина, непереполненная другими плавающими бусинами, наблюдайте за АФ вокруг нее с помощью флуоресцентной визуализации, чтобы проверить наличие пучка.
- Убедитесь, что пучок присутствует, выполнив поиск обоих флуоресцентных АФ-колокализованных. Включите источник белого света и используйте соответствующий куб фильтра для изображения каждой актиновой нити накала, повернув башню (кубы фильтра возбуждения 488 нм и 532 нм для возбуждения Alexa Fluor 488 и родамина соответственно). См. дополнительный рисунок S4.
ПРИМЕЧАНИЕ: Контрольный эксперимент по проверке интенсивности флуоресценции одиночных АФ может быть полезен при идентификации пучков, состоящих из одной 488- и одной меченой родамином нитей, или применимых к любому набору флуорофоров, который пользователь решит использовать. - После проверки задерните бусину, прикрепленную к верхней нити накаливания пучка, нажав кнопку «Затвор ловушки ».
- Используйте экранные элементы управления для записи данных, нажав на кнопку Осциллограф (Дополнительный рисунок S7). Чтобы визуализировать измерения без записи данных, нажмите кнопку Пуск. Чтобы сохранить все данные, нажмите «Автосохранение». Чтобы записать измерения, нажмите кнопку Начать запись. Выберите, какие данные должны быть визуализированы в режиме реального времени (положение, сила, направление X, направление y), выбрав из раскрывающегося меню сигнал X или сигнал Y. Помните, что xdirection находится слева направо, а y-направление — вверх и вниз на экране. См. дополнительный рисунок S7.
ПРИМЕЧАНИЕ: Данные будут сохранены в виде файлов .out и включают время, напряжение, смещение и силу для каждого направления. Эти файлы могут быть экспортированы в другое программное обеспечение для визуализации и анализа.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Проточные клетки, содержащие системы пучков актомиозина, имеют стандартную конструкцию, состоящую из слайда микроскопа и травленого покровного листа, разделенного каналом, изготовленным из двусторонней липкой ленты (рисунок 1). Затем анализ строится из покрова вверх с использованием поэтапных введений, как описано в протоколе. Окончательный анализ состоит из шаблонных актиновых нитей, меченных родамином; желаемая концентрация миозина (для репрезентативных результатов на рисунке 2 и рисунке 3 использовались 1 мкМ); биотинилированные, меченые Alexa Fluor 488 актиновые нити; 1 мкм стрептавидиновых шариков; система очистки кислорода; СПС; и буфер APB. На проточную ячейку будет образовано несколько пучков, а описанные выше концентрации актина обеспечивают адекватное расстояние между пучками, чтобы обеспечить отсутствие нежелательных взаимодействий. Это также облегчает получение нескольких измерений силы на проточную ячейку для повышения эффективности сбора данных. Силовые профили должны быть воспроизводимыми внутри проточной ячейки и от проточной ячейки к проточной ячейке.
В то время как приведенный выше протокол ориентирован на использование коммерческой оптической установки улавливания, проточная ячейка и анализ, представленные здесь, могут быть легко использованы для другого коммерческого инструмента или специально разработанной оптической установки улавливания в сочетании с микроскопом или ступенью микроскопа и обладающими возможностями флуоресцентной визуализации. Как только все добавления проточных клеток завершены в соответствии с вышеуказанным протоколом, пучки актомиозина на слайде (рисунок 1) готовы к немедленному измерению. Проточная ячейка добавляется к стадии оптической ловушки микроскопа, получают несколько измерений калибровки шариков и идентифицируют пучки путем флуоресцентной колокализации пучковых нитей. Шарик, привязанный к пучку, попадает в ловушку, и начинается смещение и соответствующее измерение силы. Пользователь может наблюдать за получением данных в режиме реального времени на мониторе компьютера. В зависимости от концентрации миозина, используемого в проточной клетке, пучок может немедленно начать демонстрировать существенное движение, или может потребоваться 30 с-1 мин, чтобы эффективно увидеть увеличение смещения / силы.
Репрезентативный след силы показан на рисунке 3А , где моторы миозина демонстрируют устойчивую рампу в силе, за которой следует плато. Типично наблюдать, как эти типы следов развиваются в течение 2-5 минут. Однако также можно измерить пучки актомиозина, которые не генерируют никакой чистой силы (рисунок 3B). Эти следы проявляются как базовый шум или не демонстрируют существенного чистого увеличения силы более 90 с. Вероятно, это связано с низкой локальной концентрацией двигателя, которая не допускает продуктивного скольжения, или пучок находится в неблагоприятной параллельной ориентации, где плюсовой и минусовый концы нитей выровнены.
Поскольку содержимое проточной ячейки может быть подвержено деградации от падающего освещения и улавливающего лазера, локального нагрева на слайде с течением времени и генерации радикальных форм кислорода, настоятельно рекомендуется не использовать одну и ту же проточную ячейку более 1 ч. Для максимальной эффективности предлагается провести еще один инкубационный анализ при получении данных. Трассировка смещения/силы может быть экспортирована из программного обеспечения оптического треппинга в Excel, Matlab, Igor или другие программы управления данными для дальнейшей фильтрации и анализа. Данные, которые могут быть извлечены из таких экспериментов с оптическим улавливанием ансамбля/связки, включают различные типы профилей генерации силы (базовая линия, рампа/плато) в различных условиях анализа, скорость генерации силы, генерация максимальной силы, кинетическое и пошаговое поведение ансамбля через размеры шагов и время ожидания между шагами или группами шагов, а также коэффициент нагрузки. Пользователь также может изменить условия анализа, чтобы сравнить, как добавление различных типов миозиновых двигателей, добавление актин-связывающих белков или изменение буферных условий влияют на эти характеристики генерации ансамблевой силы.
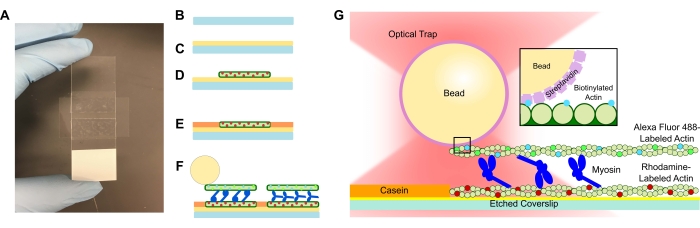
Рисунок 1: Схема анализа. (A-C) Травленые крышки покрыты поли-L-лизином и используются для формирования проточной ячейки с помощью двусторонней ленты и предметного стекла микроскопа. Синхронизированные введения и этапы инкубации, описанные в протоколе, приводят к тому, что в качестве шаблона или нижней нити (D) в качестве шаблона или нижней нити (D) следует блокировка казеина для предотвращения неспецифического связывания (E) и (F) стабилизированного фаллоидином актина Alexa Fluor 488 в качестве грузовой или верхней нити, а также команды миозина II, которые раздвигают нити и генерируют силу при введении АТФ. Геометрия двигателей и характер сшивки внутри пучка могут изменяться при различных условиях, таких как концентрация соли59. Предыдущие исследования показали, что хвостовой домен миозина обладает способностью взаимодействовать с актиновыми нитями и медленным ансамблемподвижности 46. Однако миозиновые головки в тяжелых экспериментах с меромиозином демонстрируют связывание каждой головки с соседними актиновыми нитями60. (G) Шарики стрептавидина используются в качестве оптической ручки для ловушки и связываются исключительно с грузовой биотинилированной актиновой нитью, что помогает подтвердить, что на слайде образуются надлежащие пучки. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.

Рисунок 2: Флуоресцентные пучки актомиозина. Четыре различных столкновения актиновых нитей и пучков в анализе пучка, представленном на рисунке 1. Верхняя грузовая биотинилированная актиновая нить с фаллоидиновым каналом Alexa Fluor 488 показана слева, а нижняя шаблонная актиновая нить с родаминовым фаллоидиновым каналом — справа. Внизу тот же рисунок показан с цветными линиями, наложенными, чтобы помочь направить глаз. (A) Верхняя актиновая нить находится рядом с нижней актиновой нитью, но имеет неполное наложение. Это не будет использоваться для пакетных экспериментов. (B) Верхняя и нижняя актиновые нити колокализованы, и интенсивность каждой нити накала подтверждает, что они представляют собой каждую отдельную нить внутри пучка. Это был бы хороший кандидат для пакетных экспериментов. (C) На дне находится большой пучок самосборных родаминовых нитей. Хотя существует соответствующая верхняя актиновая нить, которая колокализована, присутствует слишком много нижних нитей; таким образом, он не будет использоваться для экспериментов с пакетами. Это также пример того, как при соединении нескольких актиновых нитей одного типа интенсивность флуоресценции увеличивается. Пользователь может использовать его в качестве датчика для оценки одиночных нитей по сравнению с пучками одного и того же типа нити. D) Нижняя нить присутствует без соответствующей верхней нити накала, что также подтверждает отсутствие кровотечения. Это не будет использоваться для пакетных экспериментов. Мы отмечаем, что интенсивность нитей в канале Alexa Fluor 488 низкая, и считаем, что это связано с используемым набором фильтров (Filter Set 09 от Zeiss). Набор фильтров, используемых для канала родамина, — это набор фильтров 43 от Zeiss. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.

Рисунок 3: Генерация сил ансамбля Myosin II. Репрезентативные следы скелетных двигателей миозина II, генерирующих силу в построенной структурной иерархии актинов in vitro . Моторы миозина работают вместе, чтобы коллективно и продуктивно генерировать силу до тех пор, пока не будет достигнуто плато и сила не будет поддерживаться (A) или испытывать антагонизм вблизи исходного уровня (B). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Дополнительный рисунок S1: Оптическая ловушка Bruker/JPK Nanotracker2. (A) Компьютерный монитор. (B) Компьютерная клавиатура. (C) Компьютерная башня. (D) Блок контроллера. (E) Лазерный источник питания. (F) Оптический блок оптики ловушки. (G) Перевернутый микроскоп. (H) От двери до ступени микроскопа. (I) Ползунок поляризатора для переключения между ярким полем и дифференциальным интерференционным контрастным изображением. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S2: Пульт дистанционного управления для оптической ловушки. (A) Клавиатура для позиционирования моторизованной ступени. (Б-С) Отрегулируйте положение ловушки. (D) A, X и B включают и выключают основной затвор, захватывают 1 затвор и захватывают 2 затвора соответственно. (E) Кнопка Logitech используется для пробуждения геймпада. (F) Кнопки вверх и вниз, которые используются для позиционирования объекта захвата. (G) Кнопки вверх и вниз, которые используются для позиционирования объекта обнаружения. Обратите внимание, что пульт дистанционного управления не требуется, и все эти манипуляции могут быть выполнены в программном обеспечении. Тем не менее, удобно иметь возможность контролировать цели и положение сцены, глядя в сценическую среду микроскопа. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S3: Флуоресцентный модуль для оптической ловушки. Источник флуоресцентного белого света 89North PhotoFluor соединен с задней частью перевернутого микроскопа. Включается и выключается с помощью тумблера (стрелки). Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S4: Башня куба флуоресцентного фильтра. Башню (стрелку) можно повернуть, чтобы использовать фильтрующий куб, необходимый для визуализации в красителях DIC, rhodamine или Alexa Fluor 488. Обратите внимание, что кубы фильтров можно отключить, чтобы настроить настройку для использования различных флуорофоров. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S5: Программное обеспечение Nanotracker2. (A) Кнопка питания лазера и управление. (B) Окно объективного позиционирования. Стрелки направления используются для перемещения целей обнаружения (вверху) и захвата (внизу). Двойные стрелки перемещают цели с большей скоростью. Синяя и красная кнопки в левом нижнем углу разъединяют цели и втягивают их обратно в исходное положение. Это необходимо при взятии образцов в стадии микроскопа и вне ее. Третья кнопка слева с целями и значком замка «соединяет» цели так, что, когда они оба находятся в фокусе и достигают освещения Колера, пользователь может перемещать как захват, так и обнаружение целей вверх и вниз по оси Z. (C) Окно позиционирования образца, используемое для перемещения ступени микроскопа по осям X и Y. Двойные стрелки перемещают сцену с большей скоростью. Это окно активируется нажатием значка стрелки вверх/вниз и влево/вправо в верхнем меню. (D) Окно визуализации камеры. Значок гаечного ключа можно использовать для задания настраиваемых условий обработки изображений. Это окно активируется нажатием на значок камеры в верхнем меню. (E) Окно освещения микроскопа. Это окно активируется нажатием на значок лампочки в верхнем меню. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S6: Окно калибровки. (А) Это окно используется для калибровки шариков и активируется нажатием на значок Cal в верхнем меню. Для калибровки шарика наилучшее соответствие угловой частоты достигается в сигналах x, y и z. (B) Для каждого сигнала выберите соответствующую кнопку сигнала в левом верхнем углу. (C) Нажмите на запуск и оптимизируйте подгонку, щелкнув и перетащив в зеленое окно (D). (E) После того, как вы удовлетворены посадкой, нажмите «Использовать его » для чувствительности и жесткости. Это позволит регистрировать смещение в нанометрах и силу в пиконьютонах. (F) Затем нажмите « Принять значения» в левом нижнем углу. Повторите для направлений y и z. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок S7: Окно сбора данных. Это окно используется для получения данных о положении и силе и позволяет пользователю видеть измерения в режиме реального времени. (A) Это окно активируется нажатием значка x,t в верхнем меню. (B) Пользователь может переключаться между просмотром сигналов x и y. (C) Нажмите кнопку Пуск , чтобы начать визуализацию данных. Нажмите «Автосохранение», чтобы сохранить данные. Нажмите « Начать запись», чтобы начать запись и сохранение данных. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Исследование in vitro с использованием оптического пинцета в сочетании с флуоресцентной визуализацией было проведено для изучения динамики ансамблей миозина, взаимодействующих с актиновыми нитями. Актин-миозин-актиновые пучки собирали с использованием мышечного миозина II, родамин-актина в нижней части пучка и на поверхности покровного листа и 488 меченых биотинилированных актиновых нитей в верхней части пучка. Белок актина из мышц кролика полимеризовали и стабилизировали с использованием общих актиновых буферов (GAB) и актиновых полимеризующих буферов (APB). GAB и APB должны быть свежеприготовлены каждый день в лаборатории с использованием ATP, FC буфера и буфера TC. Мышечный миозин II использовался для образования актин-миозин-актиновых бутербродов. Фаллоидин применяли для флуоресцентного окрашивания актиновых нитей, а также стабилизации in vitro.
Активность миозина может быть подтверждена путем выполнения стандартного скользящего анализа нити, опубликованного ранее46,47. Миозин II и его субфрагменты могут связываться с поверхностью покрова в различных ориентациях, а наличие хвостового домена может замедлить скольжение нити по сравнению с анализами с использованием тяжелого меромиозина 46,48,49. Тем не менее, скольжение и движение поверхности все еще можно наблюдать. Более очевидной демонстрацией активности миозина является активное разрушение актиновой нити, которое можно наблюдать, когда более длинные актиновые нити разбиваются на более мелкие фрагменты, которые затем скользят в нескольких направлениях. Это происходит из-за высокой концентрации активных двигателей на поверхности, наблюдалось несколькими лабораториями и не происходит без активных миозиновых двигателей, присутствующих 42,50,51,52,53,54. Кроме того, анализ пучка, представленный здесь, помогает в облегчении проблем подвижности, которые в первую очередь были связаны с анализом скользящей нити, таких как разнообразие ориентаций моторного связывания на стеклянном крышке, потому что анализ пучка включает в себя блокировку казеина стеклянной поверхности таким образом, что двигатели связываются внутри пучка 47,55,56.
Первым шагом является добавление родаминовых актиновых нитей в качестве нижней или шаблонной нити к покрытому поли-L-лизином покрытому крышкой в проточной ячейке. Поли-L-лизин используется для содействия связыванию актина, поскольку полилизин заряжен положительно, в то время как актин имеет отрицательные заряды и использовался в предыдущих препаратах цитоскелетного анализа in vitro 61,62,63. Перед образованием пучка в проточную ячейку добавляли различные разведения актина для оптимизации концентрации актина. В этом случае 600x от запаса было оптимальным разбавлением, которое давало достаточное количество шаблонных нитей для формирования пучка, но с адекватным расстоянием, чтобы пучки были индивидуализированы. Разбавление проводили с использованием буфера APB. Добавление родамина актина сопровождалось слоем казеина, чтобы блокировать поверхность и избегать неспецифического связывания. Проточную клетку инкубировали в течение 30 мин и промывали после инкубации буфером для вымывания любых несвязанных актиновых нитей. Наконец, комбинацию миозина, 488/биотин актина и шариков, покрытых стрептавидином, добавляли в проточную клетку для облегчения образования актин-миозинового пучка. Концентрация шарика должна быть такой, чтобы было достаточно для связывания связанных с поверхностью пучков и достаточно в суспензии для облегчения калибровки. Однако слишком высокая концентрация бусин может вызвать трудности во время экспериментов по улавливанию из-за того, что соседние бусины попадают в лазерную ловушку и нарушают измерение. Миозиновые двигатели добавляются к комбинации непосредственно перед введением ее в затвор, так что миозиновые двигатели не превентивно агрегируются с грузом или верхней биотинилированной актиновой нитью и, таким образом, связывают нижний родамин с пучком биотинилированных актиновых нитей.
Оптическая система улавливания NT2 представляет собой коммерческую оптическую ловушку с комбинированными методами яркого поля, дифференциального интерференционного контраста (DIC) и эпифлуоресцентной визуализации. Он соединен с инвертированным микроскопом Zeiss AxioObserver 3 с целями улавливания и обнаружения воды 100x/NA 1.46 и 63x/NA 1.0. Система оснащена функцией захвата щелчка и перетаскивания одной лазерной ловушки и может использоваться при визуализации в любом из перечисленных ранее способов. Образовавшиеся пучки обнаруживаются и подтверждаются с помощью флуоресцентной визуализации. Наличие источника белого света с соответствующими фильтрующими кубами (GFP/FITC и TRITC/CY3) позволяет быстро переключаться между изображениями нити накаливания. Колокализованные АФ были проверены путем визуализации АФ на различных длинах волн возбуждения перед измерением каждой силы с использованием оптического пинцета. Поскольку нити могут быстро отбеливаться даже с помощью реагента для поглощения кислорода, исследователи могут оптимизировать параметры визуализации, такие как интенсивность и время воздействия, прежде чем выполнять эксперименты с пучком.
Для измерения силы использовалось оптическое улавливание с использованием шариков стрептавидина в присутствии АТФ для связывания нити биотинилированного актина груза и активации генерации силы миозина в качестве преобразователя силы. Данные о перемещении и силе и времени, полученные с помощью оптического улавливания, были извлечены из программного обеспечения для улавливания для анализа. Тем не менее, коммерческое программное обеспечение для треппинга также предоставляет процедуры анализа, которые могут быть использованы, или пользовательские алгоритмы в других программах могут быть запрограммированы пользователем для визуализации и анализа данных треппинга. На пользовательских оптических системах улавливания пользователь может иметь лазеры возбуждения вместо источника белого света с фильтрами, которые также приемлемы для использования. Кроме того, флуоресцентные красители могут быть изменены в соответствии с существующим оборудованием, которое может иметь пользователь, если спектры излучения не перекрываются и вызывают кровотечение.
Мы отмечаем, что представленный анализ является базовым анализом, который может быть дополнительно настроен пользователем в зависимости от его исследовательского вопроса в области механики ансамбля актомиозина. Общий рабочий процесс также может быть применен к другим системам цитоскелетного ансамбля in vitro, которые могут представлять интерес, таким как анализ пучка микротрубочек, которые образуют минимальные модели митотического веретена 32,61,63,64,65,66. Модификации могут включать, но не ограничиваются этим, изменение меток флуорофоров, которые подходят для существующей установки пользователя; изменение концентрации, конструкции или изотипа миозина; и титрование буферных условий, среди прочих аспектов.
Потенциальные проблемы возможны при выполнении этого анализа. При образовании актин-миозиновых пучков концентрация миозина в актиновых пучках может быть неоднородной по всему слайду. Чтобы приспособиться к этому, будут измерены несколько пучков по всему слайду, чтобы обеспечить правильную выборку профилей распределения двигателя и генерации силы. Также сложно узнать ориентацию расслоения, если это требуется для интерпретации данных о силе. Таким образом, для каждого пакета должно быть проведено несколько испытаний. Можно также включить маркировку конца актиновой нити через флуоресцентный гельсолин или бусины с гелсолиновым покрытием меньшего размера, чем оптическая улавливающая ручка. Флуоресцентная визуализация также может быть использована для просмотра сил компонентов x и y для определения ориентации пучка. Кроме того, поскольку состояние агрегации миозина сильно зависит от ионной силы буфера с образованием толстых нитей, происходящих при быстром разбавлении KCl, концентрация буферных солей должна контролироваться соответствующим образом67,68.
Предыдущие исследования, в которых использовались другие методы in vitro, такие как скользящие анализы, были полезны для выявления роли миозиновых доменов и изучения конфигурации и взаимодействия между миозином и другими актин-связывающими белками. Однако эти методы имеют недостаток в том, что связывание миозина на жесткой поверхности ограничит потенциал для координации между миозиновыми двигателями и, таким образом, механозондирующей обратной связи, которая возникает, чтобы определить, находится ли двигательный ансамбль в режиме с высоким или низким коэффициентом полезности 33,35,41,69. Кроме того, оптический захват с помощью одномиозиновых двигательных сетей не дает четкого понимания того, как миозиновые двигатели взаимодействуют друг с другом и с актиновыми нитями. Разработанный здесь протокол позволяет исследовать динамику моторного ансамбля миозина в совместимой иерархической актиновой сети. Он также настраивается с точки зрения характеристик ансамбля моторной нити накаливания, таких как концентрация, изоформа и буферная среда, среди других аспектов, чтобы обеспечить систематическое исследование. Представленный протокол является платформой для будущих исследований более сложных сетей актомиозина и поддерживает точность измерений смещения и генерации силы, чему способствует оптическое улавливание, которое традиционно использовалось для одномолекулярных исследований.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
У авторов нет конфликта интересов, о которых можно было бы заявить.
Acknowledgments
Эта работа частично поддерживается Исследовательской стипендией Совета аспирантов Университета Миссисипи (OA), Университетом Миссисипи Салли Макдоннелл-Барксдейл Honors College (JCW, JER), Консорциумом космических грантов Миссисипи под номером гранта NNX15AH78H (JCW, DNR) и Американской кардиологической ассоциацией под номером гранта 848586 (DNR).
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
