


ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Deze protocollen zullen gebruikers helpen bij het onderzoeken van mitochondriaal energiemetabolisme in 3D-kankercellijn-afgeleide sferoïden met behulp van Seahorse extracellulaire fluxanalyse.
Abstract
Driedimensionale (3D) cellulaire aggregaten, sferoïden genoemd, zijn de afgelopen jaren de voorhoede geworden van in vitro celkweek. In tegenstelling tot het kweken van cellen als tweedimensionale, eencellige monolagen (2D-cultuur), bevordert, reguleert en ondersteunt sferoïde celcultuur fysiologische cellulaire architectuur en kenmerken die in vivo bestaan, waaronder de expressie van extracellulaire matrixeiwitten, celsignalering, genexpressie, eiwitproductie, differentiatie en proliferatie. Het belang van 3D-cultuur is erkend in vele onderzoeksgebieden, waaronder oncologie, diabetes, stamcelbiologie en tissue engineering. In het afgelopen decennium zijn verbeterde methoden ontwikkeld om sferoïden te produceren en hun metabole functie en lot te beoordelen.
Extracellulaire flux (XF) analyzers zijn gebruikt om de mitochondriale functie in 3D-microtissues zoals sferoïden te onderzoeken met behulp van een XF24 eilandje capture plate of een XFe96 spheroid microplaat. Verschillende protocollen en de optimalisatie van het onderzoeken van mitochondriaal energiemetabolisme in sferoïden met behulp van XF-technologie zijn echter niet in detail beschreven. Dit artikel biedt gedetailleerde protocollen voor het onderzoeken van mitochondriaal energiemetabolisme in enkele 3D-sferoïden met behulp van sferoïde microplaten met de XFe96 XF-analyzer. Met behulp van verschillende kankercellijnen is aangetoond dat XF-technologie in staat is om onderscheid te maken tussen cellulaire ademhaling in 3D-sferoïden van niet alleen verschillende groottes, maar ook verschillende volumes, celnummers, DNA-inhoud en type.
De optimale mitochondriale effectorverbindingsconcentraties van oligomycine, BAM15, rotenon en antimycine A worden gebruikt om specifieke parameters van mitochondriaal energiemetabolisme in 3D-sferoïden te onderzoeken. Dit artikel bespreekt ook methoden om gegevens verkregen uit sferoïden te normaliseren en behandelt vele overwegingen die moeten worden overwogen bij het verkennen van sferoïdemetabolisme met behulp van XF-technologie. Dit protocol zal helpen bij het stimuleren van onderzoek in geavanceerde in vitro sferoïde modellen.
Introduction
Vooruitgang in in vitro modellen in biologisch onderzoek is de afgelopen 20 jaar snel vooruitgegaan. Dergelijke modellen omvatten nu organ-on-a-chip-modaliteiten, organoïden en 3D-microtissue-sferoïden, die allemaal een gemeenschappelijke focus zijn geworden om de translatie tussen in vitro en in vivo studies te verbeteren. Het gebruik van geavanceerde in vitro modellen, met name sferoïden, omvat verschillende onderzoeksgebieden, waaronder tissue engineering, stamcelonderzoek, kanker en ziektebiologie 1,2,3,4,5,6,7, en veiligheidstests, waaronder genetische toxicologie 8,9,10, nanomaterialen toxicologie11, 12,13,14, en drug veiligheid en werkzaamheid testen 8,15,16,17,18,19.
Normale celmorfologie is van cruciaal belang voor biologisch fenotype en activiteit. Het kweken van cellen tot 3D-microtissue-sferoïden stelt cellen in staat om een morfologie, fenotypische functie en architectuur aan te nemen, meer verwant aan die waargenomen in vivo , maar moeilijk te vangen met klassieke monolayer celkweektechnieken. Zowel in vivo als in vitro wordt de cellulaire functie direct beïnvloed door de cellulaire micro-omgeving, die niet beperkt is tot cellulaire communicatie en programmering (bijv. Cel-cel junctie formaties, mogelijkheden om celniches te vormen); celblootstelling aan hormonen en groeifactoren in de directe omgeving (bijv. cellulaire cytokineblootstelling als onderdeel van een ontstekingsreactie); samenstelling van fysische en chemische matrices (bijvoorbeeld of cellen worden gekweekt in stijf weefselkweekplastic of een elastische weefselomgeving); en vooral, hoe het cellulaire metabolisme wordt beïnvloed door voeding en toegang tot zuurstof, evenals de verwerking van metabole afvalproducten zoals melkzuur.
Metabole fluxanalyse is een krachtige manier om het cellulaire metabolisme binnen gedefinieerde in vitro systemen te onderzoeken. In het bijzonder maakt XF-technologie de analyse mogelijk van levende, real-time veranderingen in cellulaire bio-energetica van intacte cellen en weefsels. Gezien het feit dat veel intracellulaire metabole gebeurtenissen plaatsvinden binnen de orde van seconden tot minuten, zijn real-time functionele benaderingen van het grootste belang voor het begrijpen van real-time veranderingen in cellulaire metabole flux in intacte cellen en weefsels in vitro.
Dit artikel bevat protocollen voor het cultiveren van van kanker afgeleide cellijnen A549 (longadenocarcinoom), HepG2 / C3A (hepatocellulair carcinoom), MCF-7 (borstadenocarcinoom) en SK-OV-3 (ovarieel adenocarcinoom) als in vitro 3D-sferoïde modellen met behulp van gedwongen aggregatiebenaderingen (figuur 1). Het beschrijft ook in detail hoe mitochondriaal energiemetabolisme van enkele 3D-sferoïden kan worden onderzocht met behulp van de Agilent XFe96 XF-analyzer, (ii) wijst op manieren om XF-assays te optimaliseren met behulp van enkele 3D-sferoïden, en (iii) bespreekt belangrijke overwegingen en beperkingen van het onderzoeken van het 3D-sferoïdemetabolisme met behulp van deze aanpak. Het belangrijkste is dat dit artikel beschrijft hoe datasets worden verzameld die de berekening van zuurstofverbruik (OCR) mogelijk maken om oxidatieve fosforylering en dus mitochondriale functie in cellulaire sferoïden te bepalen. Hoewel niet geanalyseerd voor dit protocol, is extracellulaire verzuringssnelheid (ECAR) een andere parameter die wordt gemeten naast OCR-gegevens in XF-experimenten. ECAR wordt echter vaak slecht of onjuist geïnterpreteerd uit XF-datasets. We geven commentaar op de beperkingen van het berekenen van ECAR volgens basisbenaderingen van de technologiefabrikant.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

Figuur 1: Grafische workflow voor de generatie van cellulaire sferoïden, extracellulaire fluxanalyse en downstream assays. Vier kankercellijnen werden selectief gekweekt als monolagen (A), losgemaakt van weefselkweekkolven en gezaaid in ultralage 96-well microplaten om sferoïden (B) te vormen. A549 longcarcinoom, HepG2/C3A levercarcinoom, SK-OV-3 ovarieel adenocarcinoom en MCF-7 borstcarcinoomcellen werden gezaaid op 1 × 103-8 × 103 cellen/put en groeiden tot 7 dagen om enkele sferoïden te vormen en de sferoïde zaaidichtheid en teelttijd te optimaliseren door continue observatie en planimetrische metingen. Eenmaal gevormd, werden enkele sferoïden gewassen in een serumvrij XF-medium en zorgvuldig gezaaid in sferoïde assay-microplaten, voorgecoat met poly-D-lysine (C). Sferoïden werden onderworpen aan extracellulaire fluxanalyse met behulp van de XFe96-analyzer met behulp van verschillende protocollen om aan te pakken: (1) optimale sferoïde grootte voor basale mitochondriale ademhalingsrespons; (2) geoptimaliseerde titratie van mitochondriale respiratoire remmers; (3) optimalisatie van de plaatsing van sferoïden in microplaatputten. (D) Post XF-analyses, fasecontrastmicroscopie en sferoïde DNA-kwantificering werden gebruikt voor gegevensnormalisatie en andere downstream in vitro assays. Klik hier om een grotere versie van deze figuur te bekijken.
1. Teelt van kankercellijnen als 3D in vitro sferoïden
| Cellijn | Beschrijving | Cultuurmedium | Bron |
| A549 | Longcarcinoom cellijn | RPMI 1640 | Europese collectie van geauthenticeerde celculturen (ECACC) |
| Natriumpyruvaat (1 mM) | |||
| Penicilline- Streptomycine - (100 U / ml – 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| HepG2/C3A | Hepatisch carcinoom cellijn, een klonale afgeleide van de ouder HepG2 cellijn | Dmem | American Tissue Culture Collectie (ATCC) |
| Penicilline- Streptomycine - (100 U / ml – 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| Mcf7 | Borst adenocarcinoom cellijn | RPMI 1640 | Europese collectie van geauthenticeerde celculturen (ECACC) |
| Natriumpyruvaat (1 mM) | |||
| Penicilline- Streptomycine - (100 U / ml – 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| SK-OV-3 | Ovariële adenocarcinoom cellijn | RPMI 1640 | Europese collectie van geauthenticeerde celculturen (ECACC) |
| Natriumpyruvaat (1 mM) | |||
| Penicilline- Streptomycine - (100 U / ml – 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| Bestanddeel | RPMI-testmedium (50 ml eindvolume) | ||
| Basis medium | Agilent Seahorse XF RPMI, pH 7,4 | ||
| Glucose (1 M steriele voorraad) | 11 mM (0,55 ml voorraadoplossing) | ||
| L-glutamine (200 mM steriele voorraad) | 2 mM (0,5 ml voorraadoplossing) | ||
| Natriumpyruvaat (100 mM steriele bouillon) | 1 mM (0,5 ml voorraadoplossing) |
Tabel 1: Kanker cellijn media en XF media composities.
- Kweek alle cellijnen met behulp van de standaard aseptische weefselkweektechniek en bevestig dat ze vrij zijn van mycoplasma met behulp van een geschikte testkit.
- Kweek de cellijnen in T75 weefselkweekkolven of gelijkwaardig, met behulp van het aanbevolen medium (tabel 1). Kweek de cellijnen tot 65-80% samenvloeiing en passeer ze regelmatig tot een maximum van 25 passages.
- Spoel de celkweekkolven tweemaal af in dulbecco's gemodificeerde fosfaat gebufferde zoutoplossing (DBPS).
- Maak de cellen los van de kolven met 3 ml van het celdissociatiereagens (zie de materiaaltabel) gedurende 5 minuten bij 37 °C en bevestig het losmaken door microscopie.
- Aspirateer de losgemaakte celsuspensie voorzichtig om een eencellige suspensie te garanderen en deactiveer het celdissociatiereagens met 7 ml volledig weefselkweekmedium.
- Verzamel de cellen door centrifugeren op 300 × g gedurende 5 minuten, gooi het supernatant weg en resuspenseer de cellen in volledig medium.
- Tel de cellen met behulp van een hemocytometer of een geautomatiseerde celteller en titreer tot de gewenste celdichtheid die nodig is voor het zaaien.
OPMERKING: Om een volledige 96-well plaat te zaaien op 100 μL / put op 4 × 103 cellen / put, moeten cellen worden getitreerd tot 4 × 104 cellen / ml in een aanbevolen volume van 12 ml. - Decanteer de celsuspensie in een steriel reservoir en doseer 100 μL van de celsuspensie in elke put van een celafstotende microplaat met behulp van een meerkanaals pipettor.
OPMERKING: Alleen de binnenste 60 putten van een microplaat moeten worden gezaaid en de rest gevuld met DPBS. Dit vormt een verdampingsbarrière, zorgt voor sferoïde homogeniteit over de plaat en minimaliseert de effecten van de plaatrand. - Centrifugeer sferoïde microplaten op 300 × g gedurende 15 minuten om de cellen in losse aggregaten te dwingen.
- Incubeer de platen bij 37 °C, 5% CO2 gedurende minimaal 3 dagen om sferoïdevorming te garanderen.
- Voer fasecontrastmicroscopie uit met behulp van gestandaardiseerde laboratoriumpraktijken om de groei van sferoïden te volgen. Vul het celkweekmedium om de 3 dagen of twee keer per week aan door een halfvolume mediumuitwisseling uit te voeren.
2. Onderzoek van mitochondriaal energiemetabolisme van enkele sferoïden met behulp van extracellulaire flux (XF) -technologie
- Assay voorbereiding (een dag van tevoren)
- Controleer de levensvatbaarheid van sferoïden met behulp van een omgekeerde lichtmicroscoop met fasecontrast bij 4x vergroting om een intacte sferoïde structuur, morfologie en algehele uniformiteit tussen monsters te garanderen.
- Hydrateer de sensorcartridge.
- Aliquot ~20 ml van het kalibrant in een conische buis.
- Plaats de conische buis met het kalibrant een nacht in een incubator met uitzondering van37 °C die geen CO 2 is.
- Verwijder de inhoud uit de testkit.
- Verwijder de sensorcartridge van de nutsplaat en plaats deze ondersteboven op het werkblad naast de nutsplaat.
- Pipetteer 200 μL steriele ddH2O in elk putje van de sensorcartridge met behulp van een meerkanaals P300-pipet.
- Plaats de sensorcartridge bovenop de gebruiksplaat.
- Controleer of het waterniveau in elke put hoog genoeg is om de sensorsondes onder te dompelen.
- Breng de geassembleerde sensorcartridge over naar een niet-CO2-incubator van 37 °C en laat deze een nacht staan.
OPMERKING: Deze stap kan 12-72 uur voor aanvang van de test worden uitgevoerd.
- Coat sferoïde assay microplaat
- Voeg met behulp van aseptische technieken 30 μL/put steriele Poly-D-Lysine (0,1 mg/ml) oplossing toe aan de sferoïde microplaat en incubeer deze gedurende 30 minuten bij kamertemperatuur.
- Haal de oplossing uit elk putje van de sferoïde microplaat, keer de plaat om en tik deze stevig op tissuepapier om eventuele restoplossing te verwijderen.
- Was de plaat twee keer met 200 μL/putje steriel ddH2O.
- Na de laatste wasbeurt keert u de microplaat om en tikt u deze stevig op tissuepapier om eventueel restwater te verwijderen.
- Laat de plaat 30 minuten aan de lucht drogen voordat u deze gebruikt of bewaart bij 4 °C voor toekomstig gebruik.
OPMERKING: De sferoïde testmicroplaat moet worden bedekt met een moleculaire lijm om ervoor te zorgen dat de sferoïden aan de onderkant van de microplaat worden bevestigd. Zonder een moleculaire lijm kunnen sferoïden losraken en de testresultaten verstoren. Andere moleculaire lijmen kunnen ook worden gebruikt als alternatief voor Poly-D-Lysine voor het voorcoaten van platen. Voorgecoate platen kunnen bij 4 °C worden bewaard, maar moeten vóór het begin van de test op kamertemperatuur worden gebracht.
- Bereid XF Assay medium voor
- Bereid XF RPMI-medium, zoals beschreven in tabel 1, en steriel filter met een spuitfilter van 0,22 μm
- Testvoorbereiding (1 uur voorafgaand aan de test)
- Prewarm het aangevulde XF RPMI-testmedium voor tot 37 °C.
- Prewarm de gecoate sferoïde testmicroplaat in een niet-CO2 37 °C incubator of droog bad.
- Bereid de sensorcartridge voor.
- Haal de conische buis met het kalibrant en de sensorcartridge uit de luchtincubator.
- Verwijder de sensorcartridge van de gebruiksplaat en plaats deze ondersteboven op het werkoppervlak.
- Gebruik een P300 meerkanaals pipet om het water van de nutsplaat te zuigen en weg te gooien.
- Giet de kalibrantoplossing in een steriel reagensreservoir en voeg 200 μL/put van het voorgewarmde kalibrant toe aan de gebruiksplaat met behulp van een P300 meerkanaals pipet.
- Pak de sensorcartridge op en plaats deze terug op de nutsplaat, zodat de sensoren goed ondergedompeld zijn in het kalibrant.
- Breng de geassembleerde sensorcartridge terug in de niet-CO2 37 °C incubator totdat het klaar is om de poortinjectieoplossingen te laden.
- Was de sferoïden met het testmedium.
- Verwijder de sferoïde kweekplaat uit de 37 °C, 5% CO2 incubator en observeer de sferoïden onder de microscoop om hun integriteit te waarborgen voorafgaand aan de sferoïde transfer stappen.
- Laad alle putten van de sferoïde plaat met 180 μL/putje voorgewarmd testmedium, inclusief eventuele achtergrondcorrectieputten.
- Vul een petrischaaltje van 7 cm gedeeltelijk met 3 ml van het testmedium.
- Breng met behulp van een meerkanaals pipet geladen met brede opening pipetpunten de sferoïden over van de 96-well kweekplaat in petrischalen van 7 cm door de pipettor op een aspiratievolume van 10-50 μL te zetten.
- Zaadsferoïden in de voorgecoate sferoïde assay microplaat.
- Breng met behulp van een dissectiemicroscoop en een lichtbakapparaat de sferoïden over van de petrischaal naar de sferoïde testmicroplaat zoals hieronder beschreven.
- Stel het volume van een eenkanaals pipettor uitgerust met een brede pipetpunt van de opening in op 20 μL en aspirateer voorzichtig een enkele sferoïde. Plaats de punt direct in het midden van elke put van de sferoïde assay microplaat en laat de zwaartekracht een enkele sferoïde in het midden van elke put elueren, d.w.z. verdrijft geen medium uit de pipetpunt en laat capillaire actie de sferoïde uit de pipetpunt trekken. Om de elutie te bevestigen, kan de inhoud van de pipettor onder de microscoop worden teruggestort in de petrischaal van 7 cm.
OPMERKING: Zwaartekrachtelutie van een enkele sferoïde duurt meestal 15-30 s, afhankelijk van de grootte / dichtheid van de sferoïde. Gedurende deze tijd mag de pipettor niet worden verwijderd. Eventuele achtergrondcorrectieputten moeten vrij zijn van sferoïden en alleen testmedium bevatten. Bevestig onder de microscoop de positie van elke sferoïde. Elke sferoïde moet idealiter in het midden van elke put worden geplaatst. - Zodra alle sferoïden op de sferoïde assay-microplaat zijn overgebracht, brengt u de plaat over naar een niet-CO2-incubator bij 37 °C gedurende minimaal 1 uur voorafgaand aan de test.
- Stel het volume van een eenkanaals pipettor uitgerust met een brede pipetpunt van de opening in op 20 μL en aspirateer voorzichtig een enkele sferoïde. Plaats de punt direct in het midden van elke put van de sferoïde assay microplaat en laat de zwaartekracht een enkele sferoïde in het midden van elke put elueren, d.w.z. verdrijft geen medium uit de pipetpunt en laat capillaire actie de sferoïde uit de pipetpunt trekken. Om de elutie te bevestigen, kan de inhoud van de pipettor onder de microscoop worden teruggestort in de petrischaal van 7 cm.
- Breng met behulp van een dissectiemicroscoop en een lichtbakapparaat de sferoïden over van de petrischaal naar de sferoïde testmicroplaat zoals hieronder beschreven.
3. Bereiding en laden van verbindingen in de sensorcartridge voor XF-assays
| Injectie strategie | Compound (Poort) | XFe96 microwell startvolume (μL) | Gewenste eindputconcentratie | Poortvolume (μL) | Final XFe96 microwell volume na injectie (μL) | Concentratie van de werkvoorraad |
| 1 | Oligomycine (A) | 180 | 3 lug/ml | 20 | 200 | 30 μg/ml |
| Rotenon (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimycine A (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 2 | BAM15 (A) | 180 | 5 μM | 20 | 200 | 50 μM |
| Rotenon (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimycine A (B) | 200 | 2 μM | 20 | 220 | 22 μM |
Tabel 2: Mitochondriale samengestelde concentraties voor het onderzoeken van mitochondriaal energiemetabolisme van enkele 3D-sferoïden met behulp van de XFe96 Analyzer.
- Bereid de concentraties van de werkvoorraden van elke verbinding voor zoals vermeld in tabel 2 met behulp van volledig aangevuld, voorgewarmd XF RPMI-testmedium.
- Oriënteer de cartridgeplaat (gekoppeld aan de nutsplaat) kolomgewijs, 1-12 van links naar rechts.
- Als u een laadgeleider gebruikt, plaatst u deze bovenop de cartridgeplaat volgens de putlaadprocedure, bijvoorbeeld als poort A eerst wordt geladen, zorg er dan voor dat A zichtbaar is in de linkerbovenhoek van de geleider.
- Breng de werkoplossing van elke verbinding over in een geschikt reservoir en doseer met behulp van een gekalibreerde P100 meerkanaals pipet 20 μL in alle overeenkomstige poorten. Herhaal dit voor elke verbinding in de resterende poorten.
OPMERKING: Als er geen poorten op de sensorcartridgeplaat worden gebruikt, kunnen deze leeg worden gelaten of worden gevuld met een testmedium. Als alleen een selectie van een specifieke poortletter wordt gebruikt, zorg er dan voor dat de andere poorten die overeenkomen met die letter zijn geladen met een testmedium; anders wordt lucht in de put geïnjecteerd, waardoor de resultaten in die putten in gevaar komen. - Verwijder na het laden van de poort de plaatlaadgeleiders (indien gebruikt) en bereid de analysator voor op het laden van de sensorcartridge.
OPMERKING: Als de test niet onmiddellijk na het laden van de poorten wordt uitgevoerd, plaatst u het deksel terug op de sensorcartridge en plaatst u de plaat terug in de 37 °C-luchtincubator totdat deze klaar is om in de machine te worden geladen.
4. Testontwerp, injectiestrategieën en data-acquisitie
- Het uitvoeren van de test
- Schakel de analyzer in en sluit deze aan op de controller (computer).
OPMERKING: Dit kan worden geverifieerd door de verbindingsstatus van het instrument in het widgetpaneel van de Wave Controller-software. - Navigeer naar de sjablonenpagina in de WAVE-software, zoek het testsjabloonbestand voor het experiment en dubbelklik om het te openen.
OPMERKING: Als de testsjabloon niet wordt weergegeven in de weergave Sjablonen , importeert u het sjabloonbestand in de sjabloonmap vanaf een gedeeld netwerkstation of USB-flashstation. - Als u de test wilt starten, klikt u op het tabblad Assay uitvoeren .
OPMERKING: Als de groepsdefinities correct zijn toegewezen binnen de plaatkaart, is de test klaar om te worden uitgevoerd zoals aangegeven door het groene vinkje aan de rechterkant van de pagina. In dit stadium kan eventuele aanvullende informatie worden ingevoerd op de pagina met de samenvatting van de test of op de pagina die leeg is gelaten; ga verder met de volgende stap. Vanwege de vertraagde penetratie van mitochondriale modulatoren in 3D-microtissue-sferoïden (figuur 2), gebruikt u de in tabel 3 beschreven meetprotocolinformatie.
- Schakel de analyzer in en sluit deze aan op de controller (computer).
| Meetperiode | Injectienummer en poort | Meetgegevens | Duur van de periode (h:min:s) |
| Calibratie | Niet van toepassing | XF-analysatoren voeren deze kalibratie altijd uit om ervoor te zorgen dat de metingen nauwkeurig zijn | 00:20:00 (dit is een gemiddelde en kan variëren tussen machines) |
| Equillibratie | Niet van toepassing | Equilibratie vindt plaats na kalibratie en wordt aanbevolen. | 00:10:00 |
| Basaal | Niet van toepassing | Cycli = 5 | 00:30:00 |
| Mix = 3:00 | |||
| Wachten = 0:00 | |||
| Meten = 3:00 | |||
| Oligomycine / BAM15 | Injectie 1 (poort A) | Cycli = 10 | 01:00:00 |
| Mix = 3:00 | |||
| Wachten = 0:00 | |||
| Meten = 3:00 | |||
| Rotenon + antimycine A | Injectie 2 (poort B) | Cycli = 10 | 01:00:00 |
| Mix = 3:00 | |||
| Wachten = 0:00 | |||
| Meten = 3:00 | |||
| Totale tijd: | 03:00:00 | ||
Tabel 3: Protocolopstelling voor het onderzoeken van mitochondriaal energiemetabolisme van enkele 3D-sferoïden met behulp van de XFe96 Analyzer.
- Klik op Uitvoeren starten om het dialoogvenster Locatie opslaan weer te geven.
- Voer de opslaglocatie voor het resultatenbestand in en plaats de geassembleerde sensorcartridge op de thermische lade die verschijnt vanuit de deur aan de zijkant van de analysator. Wacht tot de thermische lade automatisch wordt geopend en het scherm het bericht Load Calibrant Utility Plate weergeeft. Voordat u de aanwijzingen op het scherm volgt, moet u ervoor zorgen dat i) de sensorcartridge goed op de utility-plaat past, ii) het deksel van de sensorcartridge wordt verwijderd en iii) de sensorcartridge correct op de gebruiksplaat is gericht.
- Volg de opdrachten op het scherm om de kalibratie van de sensorcartridge te starten.
OPMERKING: De tijd die nodig is om de kalibratie te voltooien is ongeveer 10-20 minuten (voor testen bij 37 °C). - Na de kalibratie van de sensorcartridge laadt u de sferoïde microplaat in de analysator door de instructies op het scherm op de Wave Controller te volgen om de evenwichtsstap van 12 minuten te starten.
OPMERKING: Groene vakjes met witte vinkjes geven een 'goede' kalibratie voor die put aan. Als putten geen 'goede' kalibratie bieden, worden ze aangegeven met een rood vak en een wit kruis. Dergelijke putten moeten worden genoteerd en uitgesloten van elke analyse nadat de test is voltooid met behulp van het tabblad modificatietest . - Wacht tot de analysator automatisch begint met het verkrijgen van nulmetingen nadat de machine de equilibratiestap heeft voltooid (zoals beschreven in het instrumentprotocol).
- Volg de opdrachten op het scherm op de WAVE-controller om het experiment te voltooien.
OPMERKING: Zodra de sferoïde microplaat uit de analysator is verwijderd, gooit u de sensorcartridge weg en legt u de sferoïdeplaat opzij voor verdere analyse indien nodig (bijv. dubbelstrengs (ds) DNA-kwantificering). Als de microplaat niet nodig is voor verdere analyse, kan deze samen met de sensorcartridge worden weggegooid. - Wacht tot het testdialoogvenster wordt weergegeven en bekijk de resultaten of keer terug naar de sjablonenweergave .
5. Datanormalisatie- en analysestrategieën - post-assay normalisatie en downstream assays (optionele stappen)
- Datanormalisatie
- Om sferoïde gegevens te normaliseren, raadpleegt u de reeks protocollen die relevant zijn voor gegevensnormalisatiestrategieën voor het berekenen van de grootte en het volume van sferoïden en het kwantificeren van dsDNA in sferoïde assays. Deze zijn opgenomen als aanvullende bestanden; zie Aanvullend dossier 1 en Aanvullend dossier 2.
- Data-analyse
- Als u gegevens wilt exporteren naar een van de geautomatiseerde analysegeneratoren, volgt u de opdrachten voor gegevensexport op de WAVE-controller en selecteert u de exportgenerator die overeenkomt met het testtype. U kunt ook het gegevensbestand exporteren en uploaden naar Seahorse Analytics.
OPMERKING: Het nadeel van rapportgeneratoren en Seahorse-analyses is dat gegevensanalyse beperkt is tot hoe de XF-test is ontworpen en niet toestaat dat gemiddelden worden genomen over meetcycli. Handmatige export van datasets uit de instrumentsoftware maakt het mogelijk om de voorkeur van de gebruiker in dit opzicht te geven. Aangezien de injectiestrategie voor het beoordelen van mitochondriale ademhaling van 3D-sferoïden waarschijnlijk zal verschillen van die van een typische 'MitoStress'-test, is een reeks spreadsheetsjablonen ontwikkeld om deze datasets te helpen analyseren, specifiek voor 3D-celculturen en zal op verzoek worden verstrekt. Deze gegevenssjabloonbestanden bieden gegevens over de belangrijkste mitochondriale ademhalingsparameters die worden beschreven en uitgelegd in figuur 2. - Als u de gegevens wilt analyseren, exporteert u de gegevens als een spreadsheetrapport vanuit de WAVE-controllersoftware en gebruikt u een onafhankelijke spreadsheetsjabloon voor analyse.
- Als u gegevens wilt exporteren naar een van de geautomatiseerde analysegeneratoren, volgt u de opdrachten voor gegevensexport op de WAVE-controller en selecteert u de exportgenerator die overeenkomt met het testtype. U kunt ook het gegevensbestand exporteren en uploaden naar Seahorse Analytics.

Figuur 2: Schematische descriptoren voor parameters afgeleid van extracellulaire fluxgegevensanalyses. Afkorting: OCR = zuurstofverbruik. Klik hier om een grotere versie van deze figuur te bekijken.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Om goed gevormde, compacte sferoïden te verkrijgen, werd elke cellijn individueel geoptimaliseerd voor de zaaidichtheid en de duur van de teelt (figuur 3). A549, HepG2/C3A en SK-OV-3 cellijnen vormden aanvankelijk losse aggregaten die pas na 7 dagen in cultuur evolueerden naar ronde sferoïden met duidelijk gedefinieerde perimeters. Omgekeerd kunnen MCF-7-cellen binnen 3 dagen sferoïden vormen. Er was een duidelijke correlatie tussen de initiële celzaaidichtheid en het sferoïde volume na de kweekperiode voor alle sferoïde modellen. Sferoïde grootte en morfologie werden geoptimaliseerd voor zaaidichtheid. Morfologie en circulariteit begonnen af te nemen met een grotere sferoïdegrootte in alle modellen. Zaaistrategieën voor cellijnen werden geoptimaliseerd bij 4 × 103 cellen/put voor A549- en SK-OV-3-cellen; HepG2/C3A-cellen zijn eerder elders geoptimaliseerd tot 1 × 103 cellen/put, en MCF-7-cellen werden gebruikt bij 4 × 103 cellen/put in alle testen. Bij geoptimaliseerde zaaistrategieën lag het sferoïdevolume tussen 5,46 × 107 μm3 (SK-OV-3) en 1,45 × 108 μm3 (A549) (figuur 3B). Alle sferoïde typen hadden een lineaire correlatie tussen de initiële zaaidichtheid en het sferoïde volume, waarbij A549 en HepG2/C3A R2-waarden van respectievelijk 0,957 en 0,947 hadden. Mcf-7 en SK-OV-3 sferoïde volumes bleken beide een grotere correlatie te hebben met de initiële zaaidichtheid, R2 =0,977 (figuur 3A).
Sferoïde circulariteit werd berekend met behulp van beeldplanimetrie binnen FIJI-analysesoftware met behulp van de lange en korte sferoïde diameters. Perfecte sferoïde symmetrie had circulariteit = 1,0; afwijking van 1,0 duidde op een verlies van circulariteit (figuur 3C). Circulariteit was groter in MCF-7 sferoïden dan andere modellen waar circulariteit werd gehandhaafd tussen 0,83 en 0,9 bij alle zaaidichtheden. Ter vergelijking: de buitenste omtrek van SK-OV-3 sferoïden was niet zo duidelijk gedefinieerd en het sferoïdevolume was zelfs na 7 dagen in cultuur significant kleiner, wat sferoïden opleverde met een maximale circulariteit van 0,61 bij een zaaidichtheid van 4 × 103 / put. HepG2/C3A-cellen bleken ook strakke, goed gevormde sferoïden te vormen met een gelijkmatige morfologie over het oppervlak van alle sferoïden, waarbij de circulariteit op 0,79 bleef voor cellen gezaaid op 1 × 103 cellen / put. A549-cellen leken een trend te volgen waarbij sferoïde circulariteit en morfologie werden verbeterd met zaaidichtheid; de circulariteit was echter niet groter dan 0,63 bij de dichtheid die in deze experimenten werd gebruikt.
Basale mitochondriale ademhaling werd berekend als OCR gemeten uit sferoïden gezaaid op 1 × 103, 2 × 103, 4 × 103, of 8 × 103 cellen / put in ultralage sferoïde cultuurmicroplaten (figuur 3D). Voor alle sferoïdetypen nam de OCR toe met de sferoïdegrootte en was lineair gecorreleerd aan het sferoïdevolume met R2 hoogste in MCF-7 sferoïden op 0,988 en het laagst in SK-OV-3 sferoïden op 0,744 (figuur 3E). De gemeten OCR was statistisch verschillend tussen alle experimentele groepen. A549 had de laagste OCR en bereikte slechts 18 pmol/min/put bij de grootste sferoïde grootte (figuur 3D). Omgekeerd leverden MCF-7-sferoïden een vergelijkbare OCR op bij de kleinste sferoïdegrootte na slechts 3 dagen in cultuur, waarbij een maximale baseline OCR van 53 pmol / min / put werd bereikt voor de grootste sferoïdegrootte (figuur 3D). HepG2/C3A leverde OCR-gegevens op die zeer consistent zijn met de grootte en morfologie van de sferoïden. In HepG2/C3A-sferoïden die werden gezaaid uit 1 × 103 cellen/put, bereikte de baseline OCR een gemiddelde van 15 pmol/min/put, oplopend tot maximaal 52 pmol/min/put in de grootste sferoïden (figuur 3D). OCR in SK-OV-3 sferoïden was alleen significant tussen sferoïden gekweekt uit 4 × 103 cellen / put en 8 × 103 cellen / put, met weinig verschil gezien in sferoïden gekweekt tussen 1 × 103, 2 × 103, of 4 × 103 cellen. Ondanks grootteverschillen waren OCR-gegevens zeer vergelijkbaar tussen HepG2 / C3A en MCF-7 sferoïden op alle groottepunten. Ten opzichte van de grootte van de sferoïden (μm3) was baseline OCR door MCF-7 sferoïden vergelijkbaar met die van HepG2/C3A-sferoïden die gedurende 7 dagen werden gekweekt uit 1.000 cellen per put.

Figuur 3: Sferoïde groeiparameters bepalen de mitochondriale ademhaling bij aanvang. (A) A549, HepG2/C3A, MCF-7 en SK-OV-3 sferoïden werden geoptimaliseerd voor zaaidichtheid en hun groei werd gecontroleerd op 1 × 103, 2 × 103, 4 × 103 en 8 × 103 cellen/put in elk van de fotomicrografieën, respectievelijk linksboven naar rechtsonder; schaalstaven = 500 μm. (B) De grootte van de sferoïden werd berekend met behulp van planimetrische gegevens van verzamelde fotomicrografieën en vergeleken met behulp van de correlatiestatistiek van Pearson; stippellijnen vertegenwoordigen de verdeling van het 95% betrouwbaarheidsinterval. (C) De morfologie van de sferoïden werd vergeleken door berekening van de circulariteit. (D) OCR werd 5x gemeten, waarna rotenon-antimycine A werd toegevoegd om rekening te houden met de niet-tirochondriële ademhalingsfrequentie met behulp van de Agilent Seahorse XFe96-analyzer. OCRbasaal gemeten als OCR - OCRr/a werd vergeleken tussen seeding density (D) en spheroid volume (E). Gegevens zijn gemiddelden ± SEM van 5-8 put repliceert per sferoïde type en celzaaidichtheid. Afkortingen: OCR = zuurstofverbruik; OCRbasaal = Basale mitochondriale ademhaling; OCRr/a = OCR na toevoeging van rotenon-antimycine A. Klik hier om een grotere versie van dit cijfer te bekijken.
De concentratie en het tijdsverloop voor blootstelling aan respiratoire modulatoren in XF-analyse is een cruciale stap in testoptimalisatie. Respiratoire modulatorverbindingen-oligomycine, BAM15, een mengsel van rotenon-antimycine A- of een DMSO-voertuigcontrole werden sequentieel geïnjecteerd via de injectiepoorten van de sensorpatroon in microplaatputten met de MCF-7-sferoïden (figuur 4A). Vier meetcycli werden voltooid om de gemiddelde basale OCR van 30-40 pmol / min / put voor alle monstergroepen te bepalen. Voor de rest van de testduur werden respiratoire modulatoren sequentieel om de 5 meetcycli toegevoegd om de uiteindelijke putconcentraties van 0,5 μM (0,5 μg/ml oligomycine) na injectie te bereiken 1; 2,0 μM (2 μg/ml oligomycine) na injectie 2; 5 μM (5 μg/ml oligomycine) na injectie 3; en ten slotte een maximale putconcentratie van 11 μM (11 μg/ml oligomycine) na de vierde sequentiële toevoeging. MCF-7-sferoïden reageerden gedurende het hele experiment niet op de voertuigcontrole (figuur 4A). Basale OCR veranderde onmiddellijk na de eerste injectie van elke respectieve verbinding in de laagste concentratie van 0,5 μM of 0,5 μg/ml oligomycine (figuur 4B). OCR in MCF-7 sferoïden werd met oligomycine verlaagd van 41 pmol/min/put naar 23 pmol/min/put na 5 meetcycli na de eerste injectie van 0,5 μg/ml (figuur 4B).
In reactie op 0,5 μm BAM15 was de OCR verhoogd van 33 naar 41 pmol/min/put vóór de tweede injectie (figuur 4C). Relatief verlaagde de combinatie van rotenon plus antimycine A de OCR van 37 naar 13 pmol/min/ruim vóór de tweede injectie (figuur 4D). Kinetische sporen onthulden verder een gestage lineaire afname (oligomycine en rotenon-antimycine A) of toename (BAM15) in OCR. Voor alle samengestelde doseringsschema's werd een steady-state OCR bereikt binnen 10-12 volledige meetcycli (60-72 min) bij een totale putconcentratie van 2 μM BAM15, 2 μM rotenon, 2 μM antimycine A en 2 μg/ml oligomycine (figuur 4A). Het zuurstofverbruik bereikte een steady-state plateau bij ~ 19 pmol / min / put (oligomycine), 52 pmol / min / put (BAM15) en 10 pmol / min / put (rotenon-antimycine A) (figuur 4A). Het verhogen van de samengestelde concentratie van oligomycine, BAM15 of rotenon plus antimycine A had verder geen duidelijk effect op OCR, dat constant bleef gedurende de rest van de test. Deze gegevens tonen aan dat zowel de concentratie van verbindingen als het tijdsverloop van blootstelling aan respiratoire modulatorverbindingen in aanmerking moeten worden genomen voor testoptimalisatie bij het gebruik van 3D-sferoïden.

Figuur 4: Titratie van respiratoire modulatorverbindingen als een belangrijke stap voor het optimaliseren van de extracellulaire fluxanalyse. (A) MCF-7 sferoïden werden gezaaid op 4 × 103 cellen / put en gekweekt gedurende 3 dagen voordat ze in putten van een sferoïde assay-microplaat met XF RPMI werden geplaatst en onderzocht op OCR-± mitochondriale modulatoren met behulp van de XFe96-analyzer. OCR werd 5x gemeten, waarna titraties van ofwel vehicle control, oligomycine (B), BAM15 (C) of rotenon-antimycine A (D) werden toegevoegd om mitochondriale ATP-synthase te remmen, de maximale ademhalingscapaciteit te bepalen of de niet-tirochondriële ademhalingsfrequentie vast te stellen. De concentratie van elke mitochondriale modulator werd verhoogd over 4 individuele titratie-injectiestrategieën (0,5 μM, 1,5 μM, 3 μM en 6 μM; eenheden voor oligomycine zijn μg / ml) om maximale steady-state OCR te bepalen in reactie op optimale samengestelde concentratie. OCR werd gemeten voor 5 meetcycli tussen elke injectie. Gegevens zijn gemiddelden ± SEM van 5-8 individuele putreplicaties. Afkorting: OCR = zuurstofverbruik. Klik hier om een grotere versie van deze figuur te bekijken.
Een van de belangrijkste voordelen van XF-technologie is het vermogen om de mitochondriale functie in intacte cellen en weefsels te onderzoeken. Om specifieke aspecten van de mitochondriale functie in cellen en weefsels te onderzoeken, worden mitochondriale modulatoren achtereenvolgens toegevoegd aan putten van de monstermicroplaat via de 4 beschikbare injectiepoorten op de sensorcartridge. De typische sequentie van modulatoren die worden gebruikt om mitochondriale parameters in XF-testen te onderzoeken, zijn oligomycine, een protonofoor (bijv. FCCP of BAM15) en een combinatie van rotenon plus AntimycinE A, die sequentieel worden toegevoegd om de mitochondriale ATP-synthase te remmen, de maximale ademhalingscapaciteit te bepalen en te corrigeren voor respectievelijk niet-chiochondriële ademhalingsfrequentie. Deze typische reeks modulatortoevoegingen wordt door de fabrikant van de testtechnologie de MitoStress-test genoemd. Aangezien oligomycine de niet-ontkoppelaar gestimuleerde ademhaling in sommige celmonolagen kan remmen20, onderzochten we dit met van kanker afgeleide 3D-sferoïden door niet-ontkoppelde gestimuleerde OCR (OCRmax) te meten vóór (enkele) en na (sequentiële) oligomycine-injectie (figuur 5A-D). OCRmax werd niet significant beperkt door de toevoeging van oligomycine in sferoïden gevormd uit HEPG2/C3A of SK-OV-3 (figuur 5E en figuur 5G). De OCRmax werd echter significant verlaagd in A549 en MCF-7 sferoïden na een sequentiële injectie van BAM15 na oligomycine in vergelijking met OCRmax bereikt door een enkele injectie van BAM15 (figuur 5F en figuur 5H). Tenzij anders bekend, wordt daarom aanbevolen om afzonderlijke putten te gebruiken voor behandeling met oligomycine en ontkoppelaar, met een laatste toevoeging van rotenon en antimycine A bij het verkennen van mitochondriaal energiemetabolisme van 3D-sferoïden. Deze aanpak maakt het nog steeds mogelijk om alle mitochondriale parameters te berekenen, zoals bij een typische MitoStress-test waarbij verbindingen sequentieel worden toegevoegd.

Figuur 5: Enkelvoudige of sequentiële injectie van mitochondriale respiratoire verbindingen. Van kankercellen afgeleide sferoïden van MCF-7, HEPG2/C3A, SK-OV-3 en A549 werden in putten van een XFe96-sferoïde microplaat in XF RPMI geplaatst en onderzocht op OCR met behulp van de Agilent Seahorse XFe96-analyzer. OCR werd 5x gemeten, waarna 2 μg/ml oligomycine (injectie poort A: groen spoor) of 5 μM BAM15 (injectie poort A: blauw spoor of injectiepoort B: groen spoor) om respectievelijk de mitochondriale ATP-synthase te remmen en de maximale ademhalingscapaciteit te bepalen. Kinetische OCR-gegevens worden uitgedrukt als % basaal (A-D). Maximale respiratoire capaciteit (OCRmax) werd berekend als een factor van basale OCR door de vergelijking: OCRmax = OCRBAM15 / OCRbasaal. OCRmax werd verkregen uit OCR-gemiddelden over meetcycli 8-10 na BAM15-injectie met (groene balken) en zonder (blauwe balken) oligomycine. Gegevens zijn gemiddelden ± SEM van 3-8 individuele put repliceert over de sferoïde testmicroplaat. Afkortingen: OCR = zuurstofverbruik. Klik hier om een grotere versie van deze figuur te bekijken.
Met behulp van optimale celzaaidichtheden, samengestelde concentraties, injectiestrategie en meetcyclusperiode bepaald in deze optimalisatie-experimenten (tabel 3), hebben we een gedetailleerd protocol ontwikkeld voor het nauwkeurig onderzoeken van basale mitochondriale ademhaling: OCRbasaal (figuur 6A), ADP fosforyleringsademhaling: OCRADP (figuur 6B), lekademhaling: OCRomy (figuur 6C), koppelingsefficiëntie (figuur 6D ), maximale ademhalingscapaciteit: OCRmax (figuur 6E) en reserve ademhalingscapaciteit:OCR-reserve (figuur 6F) met behulp van van kanker afgeleide 3D-sferoïden.

Figuur 6: Ocr onderzoeken met XF-technologie om mitochondriaal energiemetabolisme van van kanker afgeleide sferoïden vast te stellen. Van kankercellen afgeleide sferoïden van MCF-7, HEPG2/C3A, SK-OV-3 en A549 werden in putten van een sferoïde assay-microplaat in XF RPMI geplaatst en onderzocht op OCR met behulp van de Agilent Seahorse XFe96-analyzer. OCR werd 5x gemeten, waarna 2 μg/ml oligomycine, of 5 μM BAM15, en RA werd toegevoegd om respectievelijk de mitochondriale ATP-synthase te remmen, de maximale ademhalingscapaciteit te bepalen en de niet-chichondriële ademhalingsfrequentie te berekenen. (A) Basale mitochondriale ademhaling (OCRbasaal) werd berekend als het gemiddelde van OCR uit de 3 meetcycli vóór poort A-injectie. (B) De koppelingsefficiëntie van oxidatieve fosforylering werd benaderd door OCRADP (OCRbasaal- OCR-lek) tot expressie te brengen als een percentage van OCRbasaal. (C) ADP fosforyleringsinhalaming (OCRADP) werd gemeten als oligomycine-gevoelige OCR, berekend op basis van de gemiddelde OCR over meetcycli 11-13 voorafgaand aan BAM15-injectie. (D)Lek-omy-ademhaling (OCR-lek) werd gemeten als OCR-ongevoelig voor oligomycine, berekend op basis van de gemiddelde gemiddelde OCR over meetcycli 11-13. (E) De maximale ademhalingscapaciteit (OCRmax) werd gemeten als de gemiddelde maximale OCR gemeten na BAM15-injectie. (F) Reserve ademhalingscapaciteit werd berekend door OCRreserve (OCRmax - OCRbasaal) uit te drukken als een percentage van OCRbasaal. OCR na rotenon-antimycine A-injectie (OCRr/a) werd van alle tarieven afgetrokken om te corrigeren voor niet-chichondriële OCR. Gegevens zijn gemiddelden ± SEM van 3-8 individuele putreplicaties over de XFe96-sferoïdeplaat. Afkortingen: OCR = zuurstofverbruik; RA = 2 μM rotenon-2 μM antimycine A. Klik hier om een grotere versie van dit cijfer te bekijken.
MCF-7 sferoïden gekweekt uit 4 × 103 cellen / ruim 3 dagen werden gebruikt als een model om optimale overdracht, plaatsing en analyse binnen sferoïde assay microplaten te bepalen. Met behulp van de afmetingen die door de fabrikant voor de sferoïde microplaat werden verstrekt, werd het putoppervlak opgesplitst in drie zonegebieden voor een optimale plaatsing van de sferoïden (figuur 7A), waarbij zone 1 werd gemarkeerd als de optimale zone in het midden van de put. Met zorgvuldig pipetteren met behulp van pipetpunten met brede openingen werden sferoïden overgebracht naar de sferoïde platen en willekeurig verdeeld over de putoppervlakken door zwaartekrachtelution (figuur 7B). Waar sferoïden zorgvuldig werden overgebracht met behulp van zwaartekrachtelutie, konden de meeste sferoïden meestal worden gevonden in zones 1-2 van de microplaat, met behulp van de aanbevolen overdrachtstechnieken van de fabrikant. Waar sferoïden door aspiratie uit de pipetpunt werden gedwongen, werden sferoïden vaak buiten deze zones geplaatst en konden ze niet worden gezien met behulp van microscopie.
Om de plaatsingsposities van sferoïden te vergelijken, werden MCF-7-sferoïden overgebracht naar de sferoïde assay-microplaten in aangewezen zones 1-3 of buiten de zone (figuur 7A). Deze 4 putten werden gevolgd via een kinetisch experiment OCR bij baseline en na de toevoeging van oligomycine, BAM15 of rotenon-antimycine A (figuur 7C). OCR werd berekend op basis van het gemiddelde van drie cyclusmetingen vóór elke injectie (figuur 7B). OCR werd kinetisch gemeten gedurende 200 minuten in de 4 geselecteerde putten (figuur 7C) en baseline-gecorrigeerd (figuur 7D). Waar sferoïden in zone 3 of buiten de zone werden geplaatst, waren baseline OCR's significant lager dan sferoïden in zones 1 en 2 (figuur 7C). De effecten van respiratoire verbindingen oligomycine, BAM15 en rotenon-antimycine A verschilden ook dramatisch tussen sferoïden in zones 1 en 2 in vergelijking met zone 3 en out-of-zone regio's. Een toename van OCR werd waargenomen met oligomycine in sferoïden geplaatst in zone 3 of buiten zone (figuur 7E). Bovendien ondervonden sferoïden die in zone 3 of buiten zone werden geplaatst een buitensporig hoge respons op BAM15 met OCR hoger dan baseline na rotenon-antimycine A-injectie (figuur 7E). Ondanks een bijna tweevoudige toename van basale OCR (figuur 7C) met sferoïden geplaatst in zone 2 versus zone 1, waren de vouwveranderingen als reactie op alle ademhalingsverbindingen zeer vergelijkbaar (figuur 7E), wat suggereert dat verschillen in basale OCR tussen sferoïden geplaatst in zones 1 of 2 waarschijnlijk niet het gevolg zijn van plaatsing in de put.

Figuur 7: Plaatsing van sferoïden in de sferoïde assay microplaat dicteert basale OCR en mitochondriale modulatoreffecten met behulp van XF-technologie. MCF-7 sferoïden werden gezaaid op 4 × 103 cellen / put en gekweekt gedurende 3 dagen voordat ze in de putten van de sferoïde microplaat met XF RPMI werden geplaatst en onderzocht op OCR-± mitochondriale modulatoren met behulp van de Agilent Seahorse XFe96-analyzer. A) fotomicrografieën van sferoïde zoneposities in sferoïde assaymicroplaten na testduur; schaalbalk = 500 μm en OCR gevangen uit overeenkomstige putten in de loop van de tijd, uitgedrukt als pmol/min-1/put-1 (B) of % basaal (C). D) mitochondriale modulatoreffecten van MCF-7-sferoïden die in verschillende zones in de sferoïdetestmicroplaat zijn geplaatst; gegevens uitgedrukt als vouwverandering van basaal. (E) Voorbeeld kinetisch spoor dat aangeeft welke OCR-gegevensmetingen (rode cirkels) worden gebruikt om de respons van elke mitochondriale modulator voor gegevens in E te berekenen. De getoonde gegevens zijn afkomstig van individuele bronreacties. Afkorting: OCR = zuurstofverbruik. Klik hier om een grotere versie van deze figuur te bekijken.
De selectiecriteria voor de achtergrond zijn van groot belang; het gebruik van buitenste putten voor achtergrondcorrectie is niet representatief voor alle microplaatputten, wat kan leiden tot onjuiste gegevensaannames en onjuiste gegevensconclusies als gevolg van randeffecten op de sferoïde microplaat. Om deze waarneming te beoordelen, werden MCF-7-sferoïden gebruikt om de testcorrectieprocedures te vergelijken om OCR-waarden af te leiden als reactie op de toevoeging van een voertuigcontrole, oligomycine, BAM15 of rotenon-antimycine A (figuur 8). Alle respiratoire verbindingen leverden de verwachte kinetische OCR-profielen op voor de geselecteerde verbindingen, die een gemiddelde stabiele basale ademhaling van 20-30 pmol/min/put lieten zien (figuur 8A). Waar de testgegevens echter werden geanalyseerd met behulp van de buitenste putten voor correctie van de achtergrondtemperatuur (A1, A12, H1 en H12), waren de waarden die werden onthuld voor OCR na de toevoeging van ademhalingsverbindingen bijzonder laag; OCR leverde negatieve waarden op voor rotenon-antimycine A. Als reactie op deze waarnemingen werd alternatieve analyse uitgevoerd met behulp van een reeks lege putten, willekeurig verdeeld over de sferoïde microplaat, als achtergrondtemperatuurcorrectieputten (figuur 8B). Waar alternatieve achtergrondcorrectie werd toegepast, waren alle relatieve samengestelde effecten op OCR in beide analysesets hetzelfde; absolute OCR-waarden stegen echter met ongeveer 10 pmol/min/put (figuur 8). Deze gegevens benadrukken de kracht en het belang van achtergrondtemperatuurcorrectie op sferoïde assay microplaten en benadrukken het belang van gebruikersoptimalisatie voor XF-analyse.

Figuur 8: Willekeurige selectie van putten voor achtergrondcorrectie om de controle voor temperatuurgradiënten over de sferoïde testmicroplaat te verbeteren. OCR-gegevens geëxtrapoleerd uit figuur 2A met behulp van aanbevolen putten voor achtergrondcorrectie (A) versus willekeurig toegewezen putten voor achtergrondcorrectie (B). Afkorting: OCR = zuurstofverbruik. Klik hier om een grotere versie van deze figuur te bekijken.
In tegenstelling tot celmonolagen vertegenwoordigen sferoïden een heterogene aggregatie van cellen in een 3D-ruimte en vereisen daarom een grondige overweging met betrekking tot analyse, vooral bij het normaliseren van deze gegevens. Dit artikel presenteert drie benaderingen om XF-gegevens verkregen van MCF-7-sferoïden te normaliseren (figuur 9). Wanneer niet genormaliseerd, correleert OCR positief (R2 = 0,98) met de sferoïde grootte (bepaald door de initiële celzaaidichtheid) significant in vergelijking met Pearson correlatiecoëfficiënt, P = 0,0057 (figuur 9A). Deze lineaire relatie wordt verlaagd wanneer OCR wordt genormaliseerd naar de initiële celzaaidichtheid (R2 = 0,78) en correleert niet langer significant met de grootte van de sferoïden (P = 0,117, figuur 9B). Dit is ook het geval wanneer het wordt genormaliseerd tot sferoïdvolume (R2 = 0,77; Pearson correlatiecoëfficiënt P = 0,120, figuur 9C) en nucleair dsDNA-gehalte (R2 = 0,58; Pearson correlatiecoëfficiënt P= 0,233, figuur 9D). Deze gegevens benadrukken het belang van het normaliseren van XF-gegevens bij het onderzoeken van het mitochondriale metabolisme van sferoïden, vooral als ze van verschillende grootte zijn.

Figuur 9: Normalisatie van extracellulaire fluxgegevens verkregen van cellulaire sferoïden. (A) Ruwe OCR-gegevens werden verkregen uit MCF-7 gekweekt gedurende 3 dagen en uitgezet met behulp van pearson's model om een correlatiecoëfficiënt te verkrijgen tussen sferoïde zaaidichtheid en OCR; P-waarde ingesteld op 0,05. (B) Ruwe OCR-gegevens werden genormaliseerd tegen de initiële sferoïde zaaidichtheid; C) MCF-7 sferoïde volume verkregen door microscopie planimetrie; en (D) nucleair ds DNA-gehalte vergeleken met behulp van pearson's correlatiecoëfficiënt. Afkortingen: OCR = zuurstofverbruik; ds DNA = dubbelstrengs DNA. Klik hier om een grotere versie van deze figuur te bekijken.
| Cellijn | Zaaidichtheid (goed) | Sferoïde groei (dagen) | Eindvolume sferoïde (μM3) | Basale OCR (pmolO2/min/put) | Gevoeligheid voor basale OCR met (JA/NEE) |
| SKOV | 1000 | 5 | 9,52E+06 | 28 ± 3,5 | Ja |
| SKOV | 2000 | 5 | 2.38E+07 | 26 ± 1,4 | Ja |
| SKOV | 4000 | 5 | 4,92E+07 | 36 ± 3,1 | Ja |
| SKOV | 8000 | 5 | 1.11E+08 | 50 ± 7,9 | Ja |
| HepG2 | 1000 | 5 | 1.11E+07 | 15 ± 0,7 | Nee |
| HepG2 | 2000 | 5 | 2.88E+07 | 23 ± 1,8 | Ja |
| HepG2 | 4000 | 5 | 5.46E+07 | 31 ± 1,7 | Ja |
| HepG2 | 8000 | 5 | 1.21E+08 | 52 ± 2,8 | Ja |
| A549 | 1000 | 5 | 2.11E+07 | 30 ± 2,5 | Ja |
| A549 | 2000 | 5 | 3.57e+07 | 41 ± 1,6 | Ja |
| A549 | 4000 | 5 | 6,93E+07 | 53 ± 7,2 | Ja |
| A549 | 8000 | 5 | 1.44E+08 | 65 ± 8,4 | Ja |
| Mcf-7 | 1000 | 3 | 1,60E+07 | 29 ± 0,8 | Ja |
| Mcf-7 | 2000 | 3 | 2.52E+07 | 37 ± 1,7 | Ja |
| Mcf-7 | 4000 | 3 | 6,00E+07 | 46 ± 1,7 | Ja |
| Mcf-7 | 8000 | 3 | 1.06E+08 | 66 ± 2,9 | Ja |
Tabel 4: Geoptimaliseerde parameters voor het bepalen van basale OCR-metingen in enkele 3D-sferoïden. Afkorting: OCR = zuurstofverbruik.
Aanvullend bestand 1: Analyse van de grootte en het volume van de sferoïden. Klik hier om dit bestand te downloaden.
Aanvullend bestand 2: Kwantificering van dubbelstrengs DNA van sferoïden in de sferoïde microplaat. Klik hier om dit bestand te downloaden.
Aanvullend bestand 3: Aanbevelingen voor het aantal replicaties dat nodig is om betrouwbare XF-testgegevenssets te verkrijgen. Klik hier om dit bestand te downloaden.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Belangrijkste bevindingen en outputs
Dit artikel biedt een gedetailleerd protocol om het mitochondriale energiemetabolisme van enkele 3D-sferoïden te onderzoeken met behulp van een reeks van kanker afgeleide cellijnen met de XFe96 XF Analyzer. Een methode wordt ontwikkeld en beschreven voor de snelle teelt van A549, HepG2/ C3A, MCF7 en SK-OV-3 cellulaire sferoïden met behulp van celafstotende technologieën voor gedwongen aggregatie. Dit protocol behandelt veel overwegingen van het onderzoeken van het sferoïdemetabolisme met XF-technologie, waaronder (1) optimalisatie van sferoïde cultuurprotocollen en de behandeling en overdracht van sferoïden in specifieke sferoïde assay-microplaten van de technologiefabrikant vanuit hun oorspronkelijke kweekvaten; (2) de concentratie van te gebruiken ademhalingsverbindingen en de tijdsafhankelijkheid van de penetratie van verbindingen; (3) te gebruiken injectiestrategieën; en (4) manieren om gegevens tussen experimentele groepen te normaliseren. Al deze overwegingen zijn onderzocht in het huidige document en worden hieronder in meer detail besproken. Deze methoden worden gepresenteerd als vereenvoudigde benaderingen voor het genereren van consistente metabole zuurstoffluxgegevens met behulp van enkele 3D-sferoïden met de XFe96 Flux-analyzer. Deze experimentele aanpak kan worden gebruikt als uitgangspunt en rubriek voor gebruik in andere sferoïde modellen die gemakkelijk kunnen worden geïmplementeerd binnen een basislaboratoriumomgeving.
Overwegingen
Sferoïde groei, grootte en gevoeligheid van XF-technologie
Om reproduceerbare gegevens met XF-technologie vast te stellen, is het essentieel om de test voor het specifieke model te karakteriseren en te optimaliseren. Deze aanpak is relatief eenvoudig in een basale monolaag van cellen; dit brengt echter extra uitdagingen met zich mee bij het kweken van cellen als 3D-sferoïden. Tijdens de hier gepresenteerde experimenten werd het RPMI-medium van de fabrikant bij gebruik aangevuld. Hoewel wordt opgemerkt dat sommige cellijnen, namelijk HepG2 / C3A, werden gekweekt in DMEM-groeimedium, had substitutie met RPMI-DMEM-formuleringen tijdens deze relatief korte testen (~ 3-5 h) een beperkte invloed op XF-analyse. De formulering van de twee media lijken erg op elkaar en gebruikers kunnen Seahorse RPMI-media 'afstemmen' op de matrix van hun celkweekmedia door suppletie, bijvoorbeeld verhoogde glucose, verdere toevoeging van koolhydraatbronnen. Cruciaal voor de uiteindelijke formulering van alle XF-buffers en -media is de afwezigheid van fenol-rood, dat waarschijnlijk zal interfereren met de fluorescerende sondes in de XF-sondepatroonplaat, en natriumbicarbonaat, wat zal leiden tot alkaliteit vanwege het gebrek aan CO 2-buffering in celkweekincubators. Andere media en buffers kunnen in eigen beheer worden aangeschaft en/of gemaakt. Krebs Ringer HEPES-buffer is bijvoorbeeld een eenvoudige buffer die kan worden gebruikt om de ademhaling in veel verschillende cellen te beoordelen, inclusief sferoïde modellen. Gebruikers van XF-assays moeten er echter rekening mee houden dat een verandering in medium / buffer en de suppletie ervan de algehele buffercapaciteit kan veranderen. Dit is met name van belang wanneer gebruikers geïnteresseerd kunnen zijn in het meten van ECAR, waarbij de bufferfactor van het medium moet worden beoordeeld om ECAR-transformatie naar protoneffluxsnelheid (PER) mogelijk te maken.
Omdat cellulaire OCR gemeten door XF-technologie evenredig is met de celdichtheid wanneer het celnummer in de put binnen de gevoeligheid van het systeem ligt, was het belangrijk om deze relatie te onderzoeken met behulp van enkele 3D-sferoïden. Door OCR van enkele 3D-sferoïden gekweekt uit 4 verschillende kankercellijnen te onderzoeken die zijn gezaaid met dichtheden van 1.000, 2.000, 4.000 of 8.000 cellen per put, laten we zien dat de XFe96-analyzer gevoelig genoeg is om veranderingen op te pikken in de snelheid van mitochondriale ademhaling tussen 3D-sferoïden gekweekt uit verschillende celzaaidichtheden (figuur 3). We laten zien dat het optimale bereik van celzaaidichtheid, en dus sferoïde volume voor het vormen van 3D-sferoïden voor het onderzoeken van OCR, verschilt afhankelijk van het celtype. Dit blijkt uit de lineaire relatie tussen OCR en seeding density of spheroid volume (Figuur 3). Voor A549- en HepG2/C3A-cellen lag de optimale zaaidichtheid voor OCR-gevoeligheid tussen 1.000 en 8.000 cellen/put; het was 2.000-8.000 cellen/put voor MCF-7 en 4.000-8.000 cellen/put voor SK-OV-3 cellen. Deze gegevens tonen aan dat optimalisatie van de grootte van sferoïden van bijzonder belang is bij het beoordelen van OCR met behulp van XF-technologie.
Overwegingen over minimale en maximale sferoïde volumes en basale OCR
Over het algemeen zullen er altijd minimum- en maximumdrempels zijn voor meetbare OCR-parameters die door de fabrikant voor deze experimenten worden aanbevolen. Voor de XFe96-analysator zijn basale OCR tussen 20 pmol O2/min/put en 200 pmol O2/min/put respectievelijk de onder- en bovengrens. Dit is het geval met monolaagcellen en sferoïden, en waar het experimentele model zich binnen dit dynamische OCR-bereik bevindt, hangt af van de hoeveelheid biologisch materiaal die beschikbaar is, bijvoorbeeld het aantal cellen als monolaag of de grootte van sferoïden. Zie tabel 4 voor een voorbeeld van hoe OCR-drempels werden bereikt door de hier gebruikte sferoïde modellen. Het kan verstandig zijn om het zuurstofgehalte in de put waarvoor deze gegevens ook beschikbaar zijn uit deze metingen te controleren als de niveaugegevens. Dit moet routinematig worden bekeken vanuit elk experiment voor kwaliteitscontroledoeleinden. Als er zuurstoftekort in de put is, zal dit duidelijk worden gemaakt in de gegevens. Mocht dit het geval zijn, dan kan het nodig zijn om de meetcycli binnen het experiment aan te passen; bijvoorbeeld het verhogen van de mengstap, zodat het zuurstofgehalte in de put wordt teruggewonnen vóór de volgende meetperiode binnen de meetcyclus. Hoewel mogelijk, hebben we ontdekt dat dit zeer onwaarschijnlijk is voor experimenten met één sferoïde met behulp van de beschreven cellijnen.
Keuze van mitochondriale ontkoppelaars voor extracellulaire fluxtesten
Protonionoforen, zoals carbonylcyanide 4-(trifluoromethoxy) fenylhydrazon (FCCP)21, carbonylcyanide m-chloorfenylhydrazon (CCCP)22 of BAM1523, zijn krachtige chemicaliën met kleine moleculen die de elektrochemische protongradiënt over mitochondriale membranen kunnen verstoren, de productie van ATP kunnen remmen en uiteindelijk mitochondriale ademhaling kunnen ontkoppelen24 . Voor deze doeleinden worden nog steeds nieuwe kleine moleculen ontwikkeld, met name bij de behandeling van stofwisselingsziekten 25,26,27; verwijzen naar twee uitstekende recensies28,29. Omgekeerd is ontkoppeling van oxidatieve ademhaling in verband gebracht met ongewenste off-target toxiciteit30. Binnen in vitro cellulaire assays depolariseert het molecuul FCCP echter mitochondriaal membraanpotentieel en oefent het off-target effecten uit, zoals plasmamembraandepolarisatie, waardoor NA + ion flux31 wordt verstoord; interferentie met cellulaire eiwitverwerking32, en zelfs inducering van cellulaire senescentie33. BAM15 werd oorspronkelijk geïntroduceerd in 2013 als een mitochondriale ontkoppelaar met minimale invloed op plasmamembranen23, met protonofore activiteit in het micromolaire bereik in hele cellen en nanomolair bereik in geïsoleerde mitochondriën 23,34.
Gezien de potentie van FCCP op plasmamembraandepolarisatie, is BAM15 een betrouwbaarder protonofoor voor het ontkoppelen van ademhaling in intacte hele cellen in extracellulaire fluxtests. Hoewel FCCP en zijn tegenhanger, CCCP, al meer dan 50 jaar worden gebruikt om maximale ademhalingscapaciteiten te testen en nog steeds op grote schaal worden gebruikt in XF-studies, onderschat het gebruik van deze kleine moleculen vaak de mitochondriale en cellulaire metabole capaciteit. Dit is deels gekoppeld aan de reden waarom zoveel publicaties met XF-technologie in de val lopen van het rapporteren van negatieve reserve ademhalingscapaciteiten of het onderschatten van echte mitochondriale ademhalingscapaciteiten wanneer FCCP wordt gebruikt. De toegevoegde potentie van FCCP in intacte cellen en weefsels leidt vaak tot een gecompromitteerde mitochondriale functie, en cellen kunnen moeite hebben om op de juiste manier te werken om een maximale ademhalingscapaciteit te behouden gedurende meerdere meetcycli na hun toevoeging, zelfs bij zeer lage concentraties35. Daarom kan de respons van cellen op FCCP in veel studies worden gevonden om af te nemen na de initiële meetcyclusperiode. Hoewel FCCP routinematig is gebruikt voor XF-analyse, wordt BAM15 bij voorkeur gebruikt in gevallen waarbij hele cellen of sferoïde modellen betrokken zijn, aangezien het een maximale ademhalingscapaciteit kan behouden in volledig gedepolariseerde mitochondriën bij concentraties zo hoog als 10 μM 3. Bovendien induceert BAM15 effecten op extracellulaire verzuring, die samenvalt met die van nutriëntenoxidatie door de hydratatie van CO2 om HCO3- en H + in grotere mate te vormen dan FCCP3. Niettemin, in het geval van geïsoleerde mitochondriën en permeabiliseerde cellen, zou elk van deze ontkoppelaars net zo goed moeten presteren als BAM15 voor mitochondriale ontkoppeling als getitreerd in de juiste concentratie.
Kinetiek van samengestelde penetratie en assaycyclus
De concentraties, penetratie en kinetische profielen van chemische verbindingen die worden gebruikt om een typische MitoStress-test uit te voeren met 3D-sferoïden met behulp van de XF-analyzer zijn complexer om aan te pakken. Gezien het feit dat sferoïden een 3D-structuur vertonen, is de penetratie van moleculen over de diameter van de sferoïde een oneindig veel complexer proces dan over celmonolagen. De kinetische penetratie en dus gevoeligheid voor de chemotherapeutische sorafenib werd bijvoorbeeld bepaald door de sferoïde leeftijd en dus de grootte in een HepG2 sferoïde model36. Het vermogen van chemicaliën met kleine moleculen (bijv. Geneesmiddelen, nanodeeltjes) om een biologisch doelwit te bereiken, hangt af van verschillende onderliggende factoren, waaronder de complexiteit van het systeem dat dynamisch moet worden doordrongen en verspreid door 37,38. Dit geldt met name voor geneesmiddelen die zich richten op tumorweefsel39. Vergelijkbaar met tumortargeting in de context van een 3D-sferoïde, kunnen grootte, compactheid en andere fenotypische reacties zoals de expressie van geneesmiddeltransportereiwitten de penetratietijd en concentratie van een verbinding regelen die nodig is om een biologische respons uit te lokken.
In dit protocol hebben we het probleem rond penetratietijd en concentratie van kleine moleculen aangepakt als reactie op de ATP-synthaseremmer oligomycine, de protonofore en mitochondriale ontkoppelaar BAM15 en de combinatie van de Complex I- en Complex III-remmers rotenon en antimycine A. Door de OCR van enkele MCF-7-sferoïden die worden blootgesteld aan meerdere titraties van deze gemeenschappelijke ademhalingsverbindingen te onderzoeken, tonen we aan dat de optimale concentratie van elke verbinding die nodig is om een steady-state ademhalingsfrequentie te induceren binnen een vergelijkbaar bereik valt als die van monolaagcellen (figuur 4). Belangrijk is, en in tegenstelling tot hun monolaagse tegenhangers, is aangetoond dat het verhogen van het aantal meetcycli tussen injecties de sleutel is tot het bereiken van een steady-state OCR in enkele 3D-sferoïden. Deze gegevens benadrukken het belang van samengestelde penetratie en hun respectieve kinetische profielen bij het verkennen van mitochondriale ademhalingsparameters van 3D-sferoïden met behulp van deze benaderingen. Met behulp van sferoïde optimalisatie-eigenschappen, concentraties van verbindingen en meetcyclustijden op basis van de gegevens in figuur 3, figuur 4 en figuur 5, werd een gevalideerde MitoStress-test vastgesteld voor het onderzoeken van specifieke parameters van mitochondriaal oxidatief metabolisme in een reeks van kanker afgeleide 3D-sferoïden (figuur 6). Van belang, en net als sommige monolayer kankercellijnen40, werd de maximale ademhalingscapaciteit (snelheid van ontkoppelde gestimuleerde ademhaling) van bepaalde van kanker afgeleide 3D-sferoïden geremd door oligomycine (figuur 5). In het bijzonder vertoonden 3D-sferoïden gekweekt uit A549- of MCF-7-cellen een significant lagere maximale ademhalingssnelheid wanneer ze werden ontkoppeld met BAM15 na oligomycine-injectie in vergelijking met ontkoppeld door BAM15 zonder oligomycine (figuur 5F en figuur 5H). Aangezien dit effect aanwezig kan zijn in andere 3D-sferoïdeculturen, stellen we voor dat, tenzij een eerder gevalideerd protocol wordt gebruikt, de maximale ademhalingscapaciteit in 3D-sferoïden moet worden geschat zonder oligomycine.
Gelijktijdige verzameling van ECAR-gegevens als een maat voor glycolytische flux in cellulaire sferoïden
Zoals meestal te zien is in de literatuur of informatie van de technologiefabrikant, is de glycolytische snelheid van sferoïden, gemeten als ECAR, een secundaire parameter die naast OCR kan worden vastgelegd. Het berekenen van ECAR alleen is geen nuttige of zinvolle parameter in een XF-experiment, omdat het niet wordt gecorrigeerd voor de buffercapaciteit van de XF-testbuffer of de toevoeging van mitochondriale verzuring, die ontstaat door de hydratatie van CO2 tot HCO3- en H +. ECAR is pas inzichtelijk als deze datacorrecties worden toegepast, waarna het mogelijk wordt om nauwkeuriger conclusies te trekken over glycolytische flux. Om te corrigeren voor de buffercapaciteit om meer betekenisvolle PER-gegevens te genereren, moet men het volume van de microkamer voor de sferoïde microplaat kennen. De fabrikant heeft hiervoor geen echt volume kunnen leveren met de sferoïde microplaat en daarom kunnen PER-gegevens niet gemakkelijk worden bepaald. Hoewel deze metingen empirisch konden worden bereikt, viel dit buiten het bestek van dit manuscript. Echter, met de juiste correcties en het kennen van het volume van de microkamer voor een bepaalde sferoïde grootte aanwezig (bijvoorbeeld het verkrijgen van een maat voor sferoïde dichtheid) in de put, zouden ECAR-gegevens zinvol worden en zouden berekeningen van glycolytische PER kunnen worden gemaakt. Vandaar dat XF-gegevens dan informatiever kunnen zijn voor het onderzoeken van glycolytisch en oxidatief metabolisme in sferoïden, maar alleen als deze parameters diepgaand werden overwogen.
Sferoïde vorming, hantering, overdracht en beweging
Sommige cellijnen zijn beter geschikt voor de vorming van sferoïden dan andere en vormen mogelijk helemaal geen sferoïden, bijvoorbeeld MCF-7 eierstokkankercellen41,42 vormen sterk cirkelvormige sferoïden in vergelijking met andere cellijnen (figuur 3). Als een ander voorbeeld is aangetoond dat Capan-1 alvleesklierkankercellen betere sferoïden vormen dan Panc-1 of BxPC343. Evenzo is bekend dat hepatisch carcinoomcellijnen variabele vermogens hebben om compacte sferoïden 5,44 te vormen, met een waargenomen verandering in fenotype zoals verbeterd medicijnmetabolisme of de productie van albumine, zoals het geval is voor HepG2 versus HepG2 / C3A 9,45,46 of HepaRG-sferoïden 17,47,48 . Daarom moeten gebruikers de sferoïde kweektechnieken dienovereenkomstig optimaliseren en titratie-experimenten uitvoeren om de optimale zaaidichtheid en het teelttijdverloop te bepalen. Bovendien is aangetoond dat de formulering en samenstelling van testmedia van invloed zijn op de sferoïde formulering, inclusief de toevoeging van methylcellulose, vaak toegevoegd aan media om de matrixviscositeit te verhogen 43,49,50. Daarom moet de optimale celmediumsamenstelling empirisch worden bepaald voor alle gebruikte cellijnen.
Het aantal mediumuitwisselingen gedurende de sferoïde cultuur wordt bepaald door de gebruikte cellijn. Meestal is het mediumuitwisseling van het halve volume om de 2-3 dagen echter in de meeste gevallen van toepassing om voedingsstoffen aan te vullen. We gebruikten de geforceerde aggregaatbenadering om 3D-sferoïden te genereren met behulp van celafstotende microplaten uit commercieel beschikbare bronnen voor snelle ontwikkeling en inzet van sferoïde modellen in XF-analyses. Alternatieve platforms kunnen echter beter geschikt zijn om sferoïden te genereren uit andere celtypen, bijvoorbeeld hanging-drop of matrix-embedded benaderingen. In laboratoria met beperkte middelen kunnen gebruikers kijken naar de agarose-vloeistofoverlay-techniek voor de vorming van celafstotende microplaatoppervlakken51,52 om de economische kosten van de eerste stappen voor de ontwikkeling van de sferoïdemethode aanzienlijk te verminderen. De beweging van sferoïden tussen kweekvaten is noodzakelijk om XF-analyse en andere downstream-assays uit te voeren. Het gemak van overdracht wordt meestal bepaald door de grootte van de sferoïden en de totale dichtheid. We raden aan om een pipetpunt met brede openingen van P200 of P1000 te gebruiken om de sferoïde-integriteit te behouden; Pipetpunten met een kleinere boring riskeren mechanische verstoring van de sferoïde, die commercieel kan worden gekocht of, met zorg, kan worden gemaakt door eenvoudig het uiteinde van de pipetpunt bij te snijden om de opening te vergroten. Deze aanpak kan echter leiden tot furring in het plastic rond het uiteinde van de tip, wat mechanische verstoring tijdens het hanteren kan veroorzaken. Het gebruik van een achtergrondverlichting of lightbox is ook nuttig voor het hanteren en observeren van sferoïden onder een dissectiemicroscoop als een essentiële stap om de succesvolle overdracht van sferoïden in de sferoïde assay-microplaat te garanderen. Bovendien is de sferoïde positie in de put van een sferoïde testmicroplaat van bijzonder belang en heeft deze een directe invloed op ocr en samengesteld effect tijdens een typische MitoStress-test (figuur 7), hoogstwaarschijnlijk vanwege de relatie tussen de sferoïde positie en de sensorsonde fluoroforen.
Achtergrondcorrectie en temperatuurregelingsputten
Het gebruik van op microplaten gebaseerde assays is een veel gebruikte aanpak in verschillende onderzoeksgebieden; het gebruik ervan brengt echter verschillende praktische uitdagingen met zich mee. Zoals het geval is in andere experimentele benaderingen, met name die welke het 96 (of meer) array-formaat gebruiken, kunnen microplaatgeometrie en positionering de temperatuur en gasuitwisselingsgradiënten over de plaat in de loop van de tijd beïnvloeden, vaak aangeduid als 'randeffecten'53,54. We ontdekten hetzelfde voor de sferoïde assay microplaat. Volgens de richtlijnen en protocollen van de fabrikant worden de buitenste hoekputten: A1, A12, H1 en H12 altijd aangewezen als achtergrondcorrectie- en temperatuurregelingsputten voor de XFe96-analysator. Omgekeerd, met het 24-well array-formaat, worden A1 en D6 aangeduid als controleputten, naast twee andere putten gelijkmatig verdeeld over het midden van de plaat bij B4 en C3. Bij het uitvoeren van XF-sferoïde-analyse vonden we een significante afwijking in gegevens die aanvankelijk werden verzameld met behulp van de richtlijnen van de fabrikant. Dit ondanks de opname van de nodige stappen om te zorgen voor een pre-equivalentie van de test voor temperatuur en CO2-gehalte voordat met de verwerving van gegevens wordt begonnen, wat vaak negatieve waarden voor OCR oplevert na de injectie van bepaalde ademhalingsremmers (figuur 8).
We ontdekten dat deze waarnemingen waarschijnlijk te wijten zijn aan randeffecten op de sferoïde assay-microplaat. In figuur 8 vonden we dat bij het herverdelen van achtergrondcontroleputten over de microplaat, XF-gegevens ongeveer 2-voudig werden aangepast. Twee meest waarschijnlijke redenen zijn (1) als gevolg van verdampingseffecten aan de randputten, wat resulteert in een kleinere totale volumekamer voor de XFe96-sonde om uit te bemonsteren, en (2) van ontoereikende temperatuurevenwichtingen tussen die putten die zijn aangewezen voor achtergrondcorrectie en monsterputten, wat resulteert in datasets die OCR maskeren of over-opblazen. Om dergelijke uitkomsten te voorkomen, wordt daarom aanbevolen, vooral in de context van sferoïde-analyse, dat gebruikers putten die zijn aangewezen voor achtergrondcorrectie herverdelen over het geheel van de sferoïde assay-microplaat en de nodige stappen ondernemen om hun assay vooraf in evenwicht te brengen voordat ze XF-gegevens verkrijgen.
Normalisatie van gegevens
Naast het verstrekken van een gedetailleerd protocol voor het onderzoeken van mitochondriaal energiemetabolisme van enkele 3D-sferoïden met XF-technologie, presenteert dit artikel ook mogelijke manieren om mitochondriale ademhalingsfrequentiegegevens verkregen met 3D-sferoïden te normaliseren. Met behulp van respiratoire snelheidsgegevens verkregen met MCF-7-sferoïden gekweekt bij verschillende celzaaidichtheden (figuur 3), presenteren we basale mitochondriale ademhalingsfrequenties van MCF-7-sferoïden van toenemende grootte en diameter wanneer genormaliseerd tot initiële celzaaidichtheid, sferoïdevolume en dsDNA-gehalte (figuur 9). De juiste normalisatiemethode is van het grootste belang voor de nauwkeurige interpretatie van XF-datasets, met name bij het vergelijken van in vitro 3D-sferoïde modellen en verschillende celtypen. Slechte normalisatie kan leiden tot foutieve resultaten die eenvoudigweg niet kunnen worden vergeleken tussen datasets. Eiwitgehalte heeft niet de voorkeur voor normalisatie van sferoïde XF-gegevens, omdat voorbehandelingen van invloed kunnen zijn op de snelheid van eiwitsynthese zonder significant effect op de ademhalingsfrequentie. Bovendien kunnen significante, inconsistente hoeveelheden eiwit zich binden aan sferoïde microplaten bij cellyse, waardoor variatie in eiwitgehalte tussen putten wordt geïntroduceerd. Dit kan verder worden gecompliceerd in XF-analyses met behulp van sferoïden of niet-adhesieve cellen die biomoleculaire lijmen nodig hebben om aan te binden, die eiwit kunnen bevatten.
In tegenstelling tot het intracellulaire eiwitgehalte is het nucleaire DNA-gehalte onafhankelijk van het celtype en evenredig met het celgetal (figuur 9D) - een nauwkeurigere en minder tijdrovende benadering dan de uitsplitsing van sferoïden voor celnummerkwantificering. Omgekeerd vonden Yepéz et al.55, die XF-analyses uitvoerden in monolagen van fibroblastcellen, dat het normaliseren van XF-gegevens naar celaantal een grotere dispersie van gegevens introduceerde dan vóór normalisatie. Het gehalte aan nucleair DNA is onafhankelijk van de gedifferentieerde toestand of het fenotype en is daarom nauwkeuriger voor de normalisatie van sferoïde gegevens in XF-assays dan het eiwitgehalte. DNA-inhoud is ook een bewezen strategie voor de analyse van andere aan het metabolisme gekoppelde datasets56. Het is echter belangrijk op te merken dat het nucleaire DNA-gehalte wordt gekwantificeerd uit alle cellen die aanwezig zijn in de sferoïde; daarom wordt normalisatie van het DNA-gehalte niet aanbevolen voor XF-datasets waarin sferoïden behandelingen ondergaan die kunnen leiden tot aanzienlijk verlies van levensvatbaarheid van cellen. Voor dergelijke datasets, indien mogelijk, heeft normalisatie naar levensvatbaarheid van cellen de voorkeur, of de gegevens kunnen worden gecorrigeerd voor basale ademhaling.
Reserve ademhalingscapaciteit gebruiken als voorbeeld voor het belang van datanormalisatie
Reserve respiratoire capaciteit is een maat voor de snelheid van de maximale mitochondriale respiratoire capaciteit minus basale mitochondriale ademhalingsfrequentie (figuur 6). Het probleem met het rapporteren van gegevens van dit type als een snelheid, d.w.z. pmolO2 / min / goed binnen bepaalde experimenten, is echter dat de gegevens geen normalisatie bevatten. Zelfs als men sferoïde gegevens normaliseert naar celdichtheid / DNA-inhoud, sluit dit vaak de belangrijkste parameter uit die moet worden genormaliseerd voor mitochondriale dichtheid in de cellen. Aangezien een verandering in mitochondriale dichtheid zal leiden tot een proportionele verandering in basale en maximale ademhaling, zal ook de reservecapaciteit toenemen. Als bijvoorbeeld sferoïde OCRbasaal 200 is en OCRmax 400, wordt de reservecapaciteit gerapporteerd als 200; als OCRbasaal 100 is en OCRmax, dan is de reservecapaciteit ook 100; als percentage zijn ze echter allebei 50% van het maximum (of 100% van het basaal). Daarom wordt de reservecapaciteit niet gewijzigd tussen deze twee voorbeelden, ondanks verschillen in snelheden van 200 en 100 wanneer berekend als pmolen O2/min/put. Intern genormaliseerde waarden zijn betrouwbaarder en inzichtelijker om XF-gegevens beter vergelijkbaar te maken tussen studies en projecten. Om dit te doen voor de reserve ademhalingscapaciteit, hebben we ervoor gekozen om dit te presenteren als een percentage van maximale ademhaling in plaats van een absolute snelheid. Dit kan ook worden gepresenteerd als een percentage van de basale ademhaling. Dit zou het geval zijn als je met cellen of sferoïden werkt. Aangezien de locatie van de sferoïde in de microwellplaat echter de absolute OCR kan veranderen, maar niet de relatieve veranderingen met remmers of ontkoppelaars, is het belangrijker om te kijken naar intern genormaliseerde reacties in sferoïden als vouwverandering of percentages.
De sferoïde modellen die hier worden gegenereerd, presenteren een reeks celtypen en architectuur die niet kunnen worden vastgelegd in klassieke 2D-modellen. Deze omvatten heterogene, ruimtelijke rangschikking van cellen in drie dimensies, verbeterde cel-celcontacten (bijv. Vorming van gap junctions en extracellulaire matrices) en biochemische gradiënten over de sferoïde diameter (bijv. pH-gradiënten, zuurstofdiffusietoegang tot voedingsstoffen). Het gebruik van extracellulaire flux om in vitro sferoïde biologie te bestuderen, zou het mogelijk kunnen maken om optimale doelen voor medicamenteuze therapieën te identificeren door middel van metabole verstoringsobservaties. Deze kunnen worden geëxtrapoleerd van in vitro sferoïden naar in vivo tumoren en routes identificeren die zich kunnen richten op het sferoïde-tumormetabolisme, bijvoorbeeld koolhydraatgebruik tijdens sferoïde groei. Therapeutische modaliteiten kunnen effectief zijn bij het richten op sferoïden in vroege groeifasen, maar blijken minder effectief in de latere fasen van sferoïde groei naarmate de complexiteit van het metabole netwerk volwassen wordt. Tot slot zal de ontwikkeling van 3D-celkweekmodellen en geavanceerde analysetechnologieën in biologisch onderzoek een dynamisch en snel veranderend veld blijven met een onovertroffen potentieel. Extracellulaire fluxanalyse van in vitro celkweeksferoïden kan worden gebruikt als een geavanceerde onderzoeksmethode om onderzoeksresultaten te bevorderen die kunnen worden geëxtrapoleerd om de voor de mens relevante biologie beter te begrijpen, het gebruik van diermodellen in onderzoek te verminderen en patiëntgericht onderzoek te verbeteren.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs hebben geen belangenconflicten te melden.
Acknowledgments
N.J.C werd ondersteund door een BBSRC MIBTP CASE Award met Sygnature Discovery Ltd (BB/M01116X/1, 1940003)
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
Kankeronderzoek Nummer 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
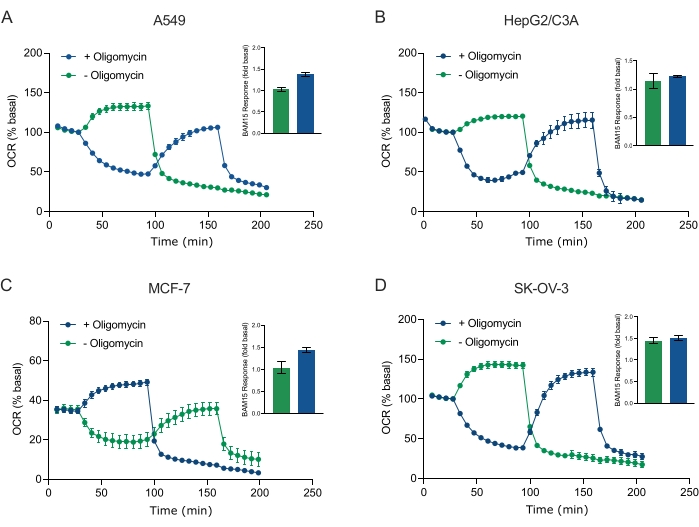
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
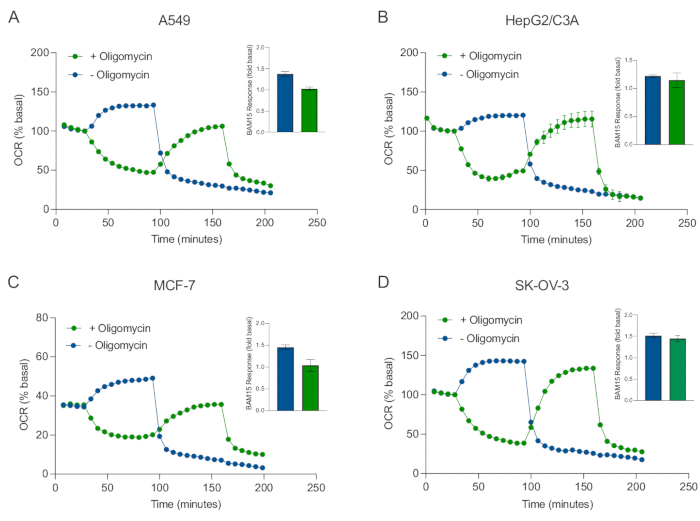
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
