

Summary
Dannelse af actomyosinbundter in vitro og måling af myosinensemblekraftgenerering ved hjælp af optisk pincet præsenteres og diskuteres.
Abstract
Myosiner er motorproteiner, der hydrolyserer ATP for at træde langs actinfilament (AF) spor og er afgørende i cellulære processer såsom motilitet og muskelkontraktion. For at forstå deres kraftgenererende mekanismer er myosin II blevet undersøgt både på enkeltmolekyle (SM) niveau og som hold af motorer in vitro ved hjælp af biofysiske metoder såsom optisk fangst.
Disse undersøgelser viste, at myosinkraftgenererende adfærd kan variere meget, når man bevæger sig fra enkeltmolekyleniveauet i et tre-perlearrangement til grupper af motorer, der arbejder sammen på en stiv perle eller dækslipoverflade i et glidende arrangement. Imidlertid tillader disse analysekonstruktioner ikke at evaluere myosins gruppedynamik inden for viskoelastisk strukturelt hierarki, som de ville inden for en celle. Vi har udviklet en metode ved hjælp af optisk pincet til at undersøge mekanikken i kraftgenerering af myosinensembler, der interagerer med flere actinfilamenter.
Disse actomyosin-bundter letter efterforskningen i et hierarkisk og kompatibelt miljø, der fanger motorkommunikation og ensemblekraftoutput. Analysens tilpassede karakter gør det muligt at ændre eksperimentelle forhold for at forstå, hvordan ændringer af myosinensemblet, actinfilamentbundtet eller det omgivende miljø resulterer i forskellige kraftudgange.
Introduction
Motorproteiner er afgørende for livet og omdanner kemisk energi til mekanisk arbejde 1,2,3. Myosinmotorer interagerer med actinfilamenter ved at tage trin langs filamenterne svarende til et spor, og dynamikken i actin-myosin-netværk udfører muskelkontraktion, cellemotilitet, den kontraktile ring under cytokinese og bevægelse af last inde i cellen, blandt andre væsentlige opgaver 3,4,5,6,7,8 . Da myosiner har så mange væsentlige roller, kan svigt i funktionaliteten af myosin-actin-netværket føre til sygdomsudvikling, såsom mutationer i myosin-tungkæden, der forårsager hjertehyperkontraktilitet i hypertrofisk kardiomyopati (HCM)9,10,11,12,13,14 . I muskelkontraktion samarbejder individuelle myosinmotorer med hinanden ved at arbejde som et ensemble for at give den nødvendige mekaniske energi, der udfører den relative glidning af AFs 4,15,16,17,18. Myosinmotorer danner tværbroer mellem AF'er og bruger konformationsændringer på grund af dets mekanokemiske cyklus til kollektivt at bevæge sig mod den piggede ende af de justerede filamenter 17,18,19,20,21.
Udvikling af kvantitative in vitro-motilitetsassays på SM-niveau ved hjælp af teknikker som optisk fangst har gjort det lettere at indsamle hidtil usete detaljer om, hvordan individuelle myosinmotorer fungerer, herunder måling af SM-kraftgenerering og trinstørrelser 22,23,24,25,26,27,28,29,30 . Finer et al. udviklede "tre-perle" eller "håndvægt" optisk fangstanalyse for at undersøge kraftgenereringsmekanikken i enkelt myosin II-motorer23,31. Da muskelmyosin II arbejder i hold for at indgå AF'er, men er ikke-processiv på SM-niveau, måtte den optiske fangstassayorientering omarrangeres fra den klassiske motorbundne perletilgang32. For at danne håndvægtsanalysen blev to optiske fælder brugt til at holde en AF over en myosinmotor bundet til en dækslip-fastgjort perle, og kraftudgangen fra den enkelte motor blev målt gennem bevægelser af AF inden for fælden23.
SM-kræfter og brug af en enkelt motor / enkelt filament assay orientering giver imidlertid ikke et fuldt billede af kraftgenerering på systemniveau, da mange motorproteiner, herunder myosin II, ikke fungerer isoleret og ofte ikke fungerer som en sum af deres dele 15,16,17,32,33,34,35,36 . Mere komplekse strukturer, der omfatter mere end en motor, der interagerer med mere end et filament, er nødvendige for bedre at forstå synergien mellem myosin- og actinfilamenternes netværk 15,32. Håndvægtsanalyseorienteringen er blevet udnyttet til at undersøge generering af små ensemblekræfter ved at have flere myosiner fastgjort til en perle eller ved hjælp af et myosintykt filament fastgjort til en overflade og lade motorerne interagere med den suspenderede AF 4,23,34,37,38,39,40.
Andre små ensembleassays omfatter et in vitro-filament-glidende assay, hvor myosinmotorer er belagt på en dækslipoverflade, og en perle bundet til en AF bruges til at undersøge den kraft, der genereres af teamet af motorer4,35,36,38,39,40,41,42,43 . I begge disse tilfælde er myosinerne bundet til en stiv overflade - perle eller dækslip - og bruger en AF. I disse tilfælde er motorerne ikke i stand til at bevæge sig frit eller kommunikere med hinanden, og det afspejler heller ikke det at have myosiner stift bundet det kompatible, hierarkiske miljø, hvor motorerne ville arbejde sammen i sarkomeren32. Tidligere undersøgelser har antydet, at myosin II kan mærke sit miljø og tilpasse sig i overensstemmelse hermed til skiftende viskoelastiske eller motoriske koncentrationsforhold ved at ændre egenskaber som kraftgenerering og toldforhold41,44,45. Der er således behov for at udvikle et optisk fangstassay, der fremmer og fanger motorkommunikation og systemkompatibilitet for at tegne et mere realistisk billede af det mekaniske grundlag for myosin II ensemblekraftgenerering.
Her udviklede vi en metode til at koble hierarkisk struktur in vitro med optisk fangst ved at danne actomyosinbundter eller sandwich bestående af flere myosinmotorer, der interagerer mellem to actinfilamenter. Denne modulære assay geometri har evnen til direkte at undersøge, hvordan molekylære og miljømæssige faktorer påvirker ensemble myosin kraft generation. Endvidere har undersøgelse af kraftgenereringsmekanismer gennem disse actin-myosin-ensembler potentialet til at hjælpe med modellering og forståelse af, hvordan store cellulære opgaver, såsom muskelkontraktion, formerer sig fra det molekylære niveau 9,10,13.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Ætsning af dæksler
- 100 g KOH opløses i 300 ml 100 % ethanol i et 1.000 ml bægerglas. Rør med en rørestang, indtil størstedelen af KOH er opløst.
FORSIGTIG: Koncentreret KOH-opløsning kan forårsage forbrændinger og skader på tøj. Brug handsker, øjenbeskyttelse og en kittel. - Placer dæksler individuelt i dækslip-rengøringsstativer.
BEMÆRK: Racks er designet med slidser, der holder enkelte dækslips adskilt for at muliggøre ætsning og skylning på hver side af dækslip, drænhuller i bunden og lavet af materiale, der kan modstå de hårde ætsningsforhold. De kan specialfremstilles eller købes kommercielt. - Der tilberedes og mærkes tre 1.000 ml bægerglas: et med 300 ml ethanol og to bægre med 300 ml omvendt osmosevand (RO).
BEMÆRK: Her blev RO-vand hentet fra en laboratorievandrenser, men det kunne også købes kommercielt, hvis en lokal renser ikke er tilgængelig. - Læg hvert af de fire bægre i en badesonikator for at afgasse i 5 min.
- Nedsænk et stativ med dæksler i bægerglasset af KOH og ethanol og sonikat i 5 min.
- Racket med dæksler overføres fra KOH/ethanolbægerglasset til bægerglasset, der kun er ethanol. Dyp stativet op og ned i bægeret, indtil der ikke er perler.
BEMÆRK: Pas på ikke at forstyrre dækslerne eller smide stativet kraftigt i bægeret. Dette vil medføre, at dækslikkerne kommer ud af stativet eller forårsager kemisk stænk. - Overfør forsigtigt stativet med dæksler fra ethanolbægerglasset til et bægerglas med vand og dypp op og ned, indtil der ikke er nogen perler.
- Nedsænk stativet med dæksler i bægerglasset med vand, der endnu ikke er brugt, og soniker igen i 5 min.
- Brug en flaske til at sprøjte stativet med dæksler med vand, indtil det løber glat af dækslerne. Gentag med ethanolen.
- Sæt stativerne til tørre i en ovn ved 90 °C i 20 min. Opbevar stativerne med ætsede dæksler ved stuetemperatur i lukkede beholdere for at forhindre forurening før brug.
2. Polymerisation af actinfilamenter
- Lav løsning T
- I et 50 ml konisk rør tilsættes 3,94 g Tris-HCl og 0,147 g CaCl2. Tilsæt RO-vand for at lave et samlet volumen på 50 ml og bland godt.
BEMÆRK: De endelige koncentrationer af opløsning T er henholdsvis 500 mM Tris-HCl og 20 mM CaCl2 . - Mærk rørets opløsning T, og opbevar det ved 4 °C.
- I et 50 ml konisk rør tilsættes 3,94 g Tris-HCl og 0,147 g CaCl2. Tilsæt RO-vand for at lave et samlet volumen på 50 ml og bland godt.
- Lav TC-buffer
- Bland 40 ml RO-vand og 1,5 ml opløsning T i et 50 ml konisk rør. Skift pH til 8,0 ved at tilføje små mængder koncentreret KOH. Tilsæt vand for at gøre 50 ml af opløsningen, og kontroller pH. Juster pH-værdien, hvis det er nødvendigt.
BEMÆRK: Den endelige TC-buffer indeholder 5 mM Tris-HCl og 0,2 mM CaCl2 ved pH 8. - Mærk røret TC og opbevar det ved 4 °C.
- Bland 40 ml RO-vand og 1,5 ml opløsning T i et 50 ml konisk rør. Skift pH til 8,0 ved at tilføje små mængder koncentreret KOH. Tilsæt vand for at gøre 50 ml af opløsningen, og kontroller pH. Juster pH-værdien, hvis det er nødvendigt.
- Lav FC Buffer
- Tilsæt 85 ml RO-vand, 10 ml opløsning T, 3,73 g KCl og 0,041 g MgCl2 til en bufferflaske på 100 ml. PH-værdien ændres til 7,5 ved at tilføje små mængder koncentreret KOH. Tilsæt vand for at lave et endeligt volumen på 100 ml og kontroller pH.
BEMÆRK: Den endelige FC-buffer indeholder 500 mM Tris-HCI, 500 mM KCl, 2 mM MgCl 2 og 2 mM CaCl2 ved pH 7,5. - Mærk røret FC, og opbevar det ved 4 °C.
- Tilsæt 85 ml RO-vand, 10 ml opløsning T, 3,73 g KCl og 0,041 g MgCl2 til en bufferflaske på 100 ml. PH-værdien ændres til 7,5 ved at tilføje små mængder koncentreret KOH. Tilsæt vand for at lave et endeligt volumen på 100 ml og kontroller pH.
- Forbered generel Actin Buffer (GAB).
- Bland 485 μL TC-buffer, 10 μL 10 mM ATP og 5 μL 50 mM DTT i et mikrocentrifugerør.
BEMÆRK: Endelige bufferbetingelser er 5 mM Tris-HCI, 0,2 mM CaCl 2, 0,5 mM DTT og0,2 mM ATP. - Mærk det som GAB og opbevar det ved 4 °C.
- Bland 485 μL TC-buffer, 10 μL 10 mM ATP og 5 μL 50 mM DTT i et mikrocentrifugerør.
- Forbered Actin Polymerization Buffer (APB).
- Bland 455 μL FC-buffer, 25 μL 100 mM ATP og 20 μL 50 mM DTT i et mikrocentrifugerør.
BEMÆRK: De endelige bufferbetingelser er 50 mM Tris-HCI, 500 mM KCl, 2 mM MgCl 2, 2 mM CaCl2 2 mM DTT og 5 mM ATP. - Mærk røret som APB, og opbevar det ved 4 °C.
- Bland 455 μL FC-buffer, 25 μL 100 mM ATP og 20 μL 50 mM DTT i et mikrocentrifugerør.
- Rekonstituere actin
- Rekonstituer kanin skeletmuskulatur actin ved at tilsætte 100 μL deioniseret vand til et 1 mg hætteglas med frysetørret actin. Bland godt ved forsigtigt at pipettere op og ned. Aliquot i 5 μL prøver, snap-frys, og opbevar 10 mg / ml actin aliquots ved -80 ° C.
- Rekonstituer biotinyleret kanin skeletmuskel actin ved tilsætning af 20 μL RO vand. Aliquot i 5 μL prøver, snap-frys, og opbevar 1 mg / ml biotinylerede actin aliquots ved -80 ° C.
- Ikke-mærket actinpolymerisation med rhodamin phalloidin stabilisering
- Optø et hætteglas med 10 mg/ml actin og hold det på is.
- Forbered frisk GAB-buffer, tilsæt 100 μL GAB til actin aliquot, og bland ved forsigtigt at pipettere op og ned. Inkuber opløsningen på is i 1 time.
- Forbered frisk APB under inkubationen. Efter inkubation polymeriseres actinen i filamenter ved at tilsætte 11 μL APB til actinopløsningen. Bland godt ved forsigtigt at pipettere op og ned. Placer på is i 20 min.
- Der tilsættes 5 μL rhodaminmærket phalloidin til den friskpolymeriserede actinfilamentopløsning. Lad stå på is i mørket i 1 time.
- Hætteglasset med rhodamin actin indpakket i aluminiumsfolie opbevares i mørke ved 4 °C.
BEMÆRK: Det anbefales at bruge disse filamenter i en maksimal periode på 1 uge. AF-kvaliteten kan bekræftes hver dag gennem en hurtig billeddannelse af en flowcelle, der kun indeholder AF'er, og visning af ensartede filamenter fra dag til dag.
- Biotinyleret actinpolymerisation med Alexa Fluor 488 phalloidinstabilisering
- Optø et hætteglas med 10 mg/ml actin og 1 hætteglas med 1 mg/ml biotinyleret actin, og hold dem på is.
- Lav frisk GAB-buffer.
- Kombiner de to hætteglas (trin 2.8.1) i et 10:1 actin:biotinyleret actinforhold. Tilsæt 100 μL GAB til actinblandingen og bland godt ved forsigtigt at pipettere op og ned. Inkuberes på is i 1 time.
- Lav frisk APB under inkubationen.
- Efter inkubationstrinnet polymeriseres actinen ved at tilsætte 11 μL APB til actinopløsningen. Bland godt ved at pipettere forsigtigt op og ned. Inkuberes på is i 20 min.
- Tilsæt 5 μL Alexa Fluor 488-mærket phalloidin og inkuber på is i mørke i 1 time.
- Opbevar det biotinylerede actinhætteglas indpakket i aluminiumsfolie i mørke ved 4 °C.
BEMÆRK: Disse filamenter kan bruges i en maksimal periode på 1 uge.
3. Myosin og perle forberedelse
- Rekonstituere Myosin II
- Spin kort ned (~ 5 s) frysetørret skeletmyosin II for at samle det i bunden af røret ved hjælp af en standard minicentrifuge.
- Myosinen rekonstitueres til 10 mg/ml ved tilsætning af 100 μL 1 mM DTT fremstillet i RO-vand.
- Fortynd stammyosinopløsningen 10x ved at tilsætte 10 μL 10 mg/ml myosin til 90 μL 1 mM DTT i RO-vand. Lav små volumen (1-5 μL) aliquoter, snap-frys og opbevar ved -80 ° C.
BEMÆRK: Myosinaktivitet kan bekræftes ved at udføre et standard glidefilamentassay som tidligere offentliggjort46,47. Se diskussionen for en kort beskrivelse.
- Rengøring af streptavidinbelagte perler
- Fortynd 20 μL 1 μm streptavidinperler i 80 μL RO-vand. Vask fire gange ved at dreje ned ved 9.600 × g og rekonstituere i 100 μL RO-vand.
- Sonicate i 2 min ved 40% amplitude og opbevare de vaskede perler på en rotator ved 4 °C.
4. Forberedelse af flowcelle
- Forbered en poly-l-lysinopløsning (PLL) ved at tilsætte 30 ml 100% ethanol til et 50 ml rør og tilsæt 200 μL 0,1% w / v poly-l-lysin i vand og bland godt.
- Tilsæt en ætset dæksel til PLL-opløsningen, og lad den trække i 15 minutter. Fjern dækslikket med en pincet, og sørg for kun at røre ved kanten af dækslip, når det trækkes op fra røret (se figur 1A-C). Grib dækslerne ved deres kanter med en behandsket hånd.
- Tør dækslippen med et filtreret flyselskab, indtil der ikke er ethanol tilbage og ingen rester på dækslip.
- Påfør to stykker dobbeltsidet klæbebånd på midten af et mikroskopglas, 3-4 mm fra hinanden. Riv eller skær det overskydende tape af, der hænger ud over kanten af diaset.
- Tilføj den PLL-belagte dæksel oven på båndet vinkelret på mikroskopets lange akse (danner et T) for at danne en kanal.
- Brug et lille rør til at komprimere dækslikket på båndet, og mikroskopet glider grundigt, indtil båndet er gennemsigtigt (figur 1A). Sørg for, at der ikke er bobler i båndet, da dette kan forårsage lækage fra flowkanalen.
BEMÆRK: Flowcellen kan rumme et volumen på 10-15 μL.
5. Forberedelse af Actomyosin-bundt
- I separate rør fortyndes hver type actinfilament (rhodamin- og biotinyleret 488-mærket) 600x ved at blande 0,5 μL af den respektive, mærkede actin med 300 μL APB. Tilsæt yderligere 5 μL af det tilsvarende mærkede phalloidin til hvert rør og inkuber på is i mørke i 15 minutter.
- Til den biotinylerede actinopløsning tilsættes et iltrensningssystem på 1 μL beta-D-glucose ved 500 mg / ml, 1 μL glucoseoxidase ved 25 mg / ml og 1 μL katalase ved 500 enheder / ml. Tilsæt 1 μL 100 mM ATP og 1 μL 100x fortyndede, rensede streptavidinperler. Rør forsigtigt med en pipettespids. Suspensionen sættes på en rotator ved 4 °C, mens resten af actomyosinbundtet samles.
- Der tilsættes 15 μL af det fortyndede rhodaminactin til PLL-flowcellen (figur 1D). Væg den overskydende opløsning gennem flowcellen, men lad ikke strømningskanalen blive tør. Inkuberes i 10 minutter i et fugtighedskammer.
BEMÆRK: Fugtighedskamre kan fremstilles af tomme pipettespidskasser med vand tilsat til bunden og låget dækket af aluminiumsfolie for at blokere lyset. - Der fremstilles en 1 mg/ml kaseinopløsning i APB.
- Der tilsættes 15 μL 1 mg/ml kasein for at forhindre uspecifik binding af de efterfølgende komponenter (figur 1E). Inkuberes i 5 minutter i et fugtighedskammer.
- Tilsæt den ønskede koncentration af myosin til det biotinylerede actin og perlesuspension fra trin 5.2. Rør forsigtigt med pipettespidsen, og tilsæt derefter straks 15 μL af trin 5.2-suspensionen + den ønskede myosinkoncentration til flowcellen (figur 1F, G). Inkuberes i 20 min. Forsegl de åbne ender af flowcellen med neglelak for at forhindre fordampning under billeddannelse og optiske fangsteksperimenter.
BEMÆRK: En myosinopløsningskoncentration på 1 μM giver robust bundling og kan bruges som udgangspunkt for den ønskede tilpasning af analysen (se figur 2).
6. Kraftmålinger ved hjælp af optisk fælde (NT2 Nanotracker2)
BEMÆRK: Mens protokollen nedenfor er specifikt til NT2-systemet, kan dette assay bruges med andre optiske fangstsystemer, herunder dem, der er specialbyggede, der også har fluorescensfunktioner. Den generelle arbejdsgang forbliver den samme som at få overfladen af diaset i fokus, udføre perlekalibreringer og erhverve data ved at finde fluorescerende actinbundter. For NT2-systemet indeholder supplerende figur S1, supplerende figur S2, supplerende figur S3, supplerende figur S4, supplerende figur S5, supplerende figur S6 og supplerende figur S7 nærmere oplysninger om det optiske fældesystem og softwaregrænsefladen.
- Tænd for kontrolboksen og laseren (supplerende figur S1).
- Start computersoftwaren til optisk fælde ved at klikke på JPK Nanotracker-ikonet på skrivebordet.
- Væk fjernbetjeningen ved at klikke på Logitech-knappen i midten (supplerende figur S2).
- Tænd fluorescensmodulet ved at trykke på tænd/sluk-knappen (supplerende figur S3).
- Drej filterkubetårnet til lysfeltbilleddannelse (supplerende figur S4).
- Når systemet er klar, skal du tænde laseren ved hjælp af lasertænd/sluk-knappen i nederste venstre hjørne af skærmen til 50 mW og lade den stabilisere sig i 30 minutter (supplerende figur S5).
- Klik sekventielt på knapperne Belysning, Kamera, Objektiv og Scenebevægelse i softwaren for at åbne disse vinduer til visning og manipulation under eksperimentet. Tænd for mikroskopbelysningen ved at klikke på tænd / sluk-knappen og indstille den til maksimal effekt ved at klikke og trække bjælken helt til højre (supplerende figur S5).
- Åbn prøveområdet, og fjern prøveholderen fra mikroskopstadiet. Tilføj flowcellen, fastgør den med metalprøveholderne, og sørg for, at diaset med dækslippet er i bunden.
- Der tilsættes 30 μL RO-vand til midten af bundmålet. Lad ikke pipettespidsen røre linsen. Indsæt prøvefasen igen.
BEMÆRK: Da NT2-systemet bruger et vandnedsænkningsmål som fangstmål, kan nedsænkningsmediet være forskelligt afhængigt af fangstmålet i brugerens opsætning. - Hæv det nederste mål ved hjælp af kontrolpilene på skærmen eller L2 på fjernbetjeningen, indtil vandperlen rører dækslippet (supplerende figur S5).
- Sænk det øverste mål, indtil ca. halvdelen af afstanden til flowcellen er nået ved hjælp af skærmens pile eller R2 på fjernbetjeningen. Tilsæt 170 μL RO-vand til toppen af flowcellen direkte under det øverste mål. Sænk det øverste mål, indtil det bryder vandets overfladespænding og danner en menisk.
- Flyt mikroskoptrinnet ved hjælp af pilepuden på fjernbetjeningen, indtil kanten af båndet ved siden af flowkanalen er nået. Luk prøvedøren.
BEMÆRK: Et "klik" ved lukning af prøvedøren indikerer, at laserlukkeren nu er åben. Dette er en sikkerhedsfunktion, der kun tillader lukkeren at åbne, hvis døren er lukket. - Brug vinduet Objektiv på skærmen til at bringe kanten af båndet i fokus ved at bringe det nederste mål med navnet Lasermål op ved at klikke på den øverste pil ved hjælp af kontrolelementerne på skærmen. Gør det samme for det øverste mål ved at klikke på den nederste pil (supplerende figur S5).
BEMÆRK: De dobbelte pile flytter målet eller trinnet hurtigere. Kanten af båndet bruges til fokusering, fordi det er et stort, let at finde objekt, der er tæt på dækslipoverfladen. Luftbobler i båndet er en anden mulighed. Dette er dog ikke nødvendigt, hvis brugeren har en automatiseret rutine for at finde overfladefokus eller en foretrukken intern metode. - Når båndet er i fokus, skal du delvist lukke iris øverst på den optiske fælde. Bring det øverste mål ned, indtil irisens polygonform er synlig. Bring disse kanter i fokus, åbn irisen igen, og par derefter målene ved hjælp af ved at klikke på hængelåsikonet (supplerende figur S5).
- Find en flydende perle, og fang den ved at klikke på knappen Trap Shutter , som åbner lukkeren og tillader fangstlaseren at ramme prøven. Klik på trapmarkøren på skærmen, og træk den for at flytte placeringen af diffuseringslaseren. Når den er fanget, skal du kalibrere perlen for at korrelere spændingsmålinger for at tvinge og forskyde.
- Klik på knappen Kalibrering . Juster kalibreringsrutinen baseret på effektspektralanalyse, og tilpas hjørnefrekvensen i softwaren til X-, Y- og Z-retningerne (supplerende figur S6).
- Klik på Indstillinger. Indtast diameteren af perlen (1.000 nm), og indtast temperaturen på scenen, der findes nederst til venstre i softwarevinduet. (se supplerende figur S6).
- Klik på Trap 1. Klik på X Signal. Klik på Kør for at udføre hjørnefrekvenstilpasningen. Klik og træk i vinduet for at optimere funktionstilpasningen. Klik på Brug det for følsomhed og stivhedsværdier. Klik på Accepter værdier. Gentag for Y- og Z-signalerne. Luk vinduet. (se supplerende figur S6).
BEMÆRK: Perlekalibreringsrutiner på andre optiske fældesystemer eller specialbyggede systemer, der er blevet robust testet af brugeren, såsom ækvipartitionsmetoden eller trækkraftmetoden, er også acceptable57,58. - Find et actomyosin-bundt ved at søge efter perler, der er bundet til AF'er på overfladen af dækslippet.
- Når en perle, der ikke er fyldt med andre flydende perler, opdages, skal du observere AF'erne omkring den ved fluorescensbilleddannelse for at verificere tilstedeværelsen af et bundt.
- Kontroller, at et bundt er til stede ved at kigge efter begge fluorescerende AF'er colokaliseret. Tænd for den hvide lyskilde, og brug den relevante filterterning til at afbilde hvert actinfilament ved at dreje tårnet (henholdsvis 488 nm og 532 nm excitationsfilterterninger til Alexa Fluor 488 og rhodamin excitation). Se supplerende figur S4.
BEMÆRK: Et kontroleksperiment til verifikation af fluorescensintensiteten af enkelte AF'er kan være nyttigt til at identificere bundter, der består af en enkelt 488- og enkelt rhodaminmærkede filamenter eller gælder for det sæt fluorophorer, brugeren vælger at bruge. - Når den er verificeret, skal du fange perlen, der er fastgjort til bundtets øverste filament, ved at klikke på trapudløserknappen .
- Brug kontrolelementerne på skærmen til at registrere dataene ved at klikke på oscilloskopknappen (supplerende figur S7). For at visualisere målinger uden at registrere dataene skal du klikke på Start. For at gemme alle data skal du klikke på Autosave. For at registrere målinger skal du klikke på Start Record. Vælg, hvilke data der skal visualiseres i realtid (position, kraft, x-retning, y-retning) ved at vælge fra rullemenuen X-signal eller Y-signal. Husk, at xdirection er fra venstre mod højre, og y-retning er op og ned på skærmen. Se supplerende figur S7.
BEMÆRK: Data gemmes som .out-filer og inkluderer tid, spænding, forskydning og kraft for hver retning. Disse filer kan eksporteres til anden software til visualisering og analyse.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Flowceller, der indeholder actomyosinbundtsystemerne, er af standarddesign, der består af et mikroskopglas og en ætset dækslip adskilt af en kanal lavet af dobbeltsidet klæbebånd (figur 1). Analysen bygges derefter fra coverslip op ved hjælp af iscenesatte introduktioner som beskrevet i protokollen. Det endelige assay består af skabelon rhodamin-mærkede actinfilamenter; den ønskede myosinkoncentration (1 μM blev anvendt til de repræsentative resultater i figur 2 og figur 3); biotinyleret, Alexa Fluor 488-mærkede actinfilamenter; 1 μm streptavidin perler; iltrensningssystemet; ATP; og APB-buffer. Flere bundter vil blive dannet pr. flowcelle, og de ovenfor beskrevne actinkoncentrationer giver tilstrækkelig afstand mellem bundter for at sikre, at der ikke opstår uønskede interaktioner. Dette letter også opnåelse af flere kraftmålinger pr. flowcelle for at øge dataindsamlingseffektiviteten. Kraftprofiler skal kunne reproduceres i en flowcelle og fra flowcelle til flowcelle.
Mens protokollen ovenfor er rettet mod brugen af en kommerciel optisk fangstopsætning, kan flowcellen og analysen, der præsenteres her, let bruges til et andet kommercielt instrument eller specialbygget optisk fangstopsætning kombineret med et mikroskop eller mikroskoptrin og besidder fluorescensbilleddannelsesfunktioner. Når alle flowcelletilsætninger er fuldført i henhold til ovenstående protokol, er actomyosinbundterne på diaset (figur 1) klar til øjeblikkelig måling. Flowcellen føjes til det optiske fældemikroskoptrin, der opnås flere perlekalibreringsmålinger, og bundter identificeres gennem fluorescens-colokalisering af bundtfilamenterne. En perle bundet til et bundt er fanget, og forskydningen og den tilsvarende kraftmåling begynder. Brugeren kan observere erhvervelsen af data i realtid på computerskærmen. Afhængigt af koncentrationen af myosin, der anvendes i flowcellen, kan bundtet begynde at udvise betydelig bevægelse med det samme, eller det kan tage 30 s-1 minut at effektivt se en stigning i forskydning / kraft.
Et repræsentativt kraftspor er vist i figur 3A , hvor myosinmotorerne udviser en stabil rampe i kraft efterfulgt af et plateau. Det er typisk at se disse typer spor udvikle sig over 2-5 min. Det er dog også muligt at måle actomyosinbundter, der ikke genererer nogen nettokraft (figur 3B). Disse spor fremstår som basisstøj eller udviser ingen væsentlig nettostigning i kraft over 90 s. Dette skyldes sandsynligvis en lav lokal koncentration af motor, der ikke tillader produktiv glidning, eller bundtet er i en ugunstig parallel orientering, hvor plus- og minusenderne af filamenterne er justeret.
Da indholdet af flowcellen kan være modtageligt for nedbrydning fra den indfaldende belysnings- og fangstlaser, lokal opvarmning på rutsjebanen over tid og generering af radikale iltarter, anbefales det kraftigt ikke at bruge den samme flowcelle i mere end 1 time. For maksimal effektivitet foreslås det at have et andet assay, der inkuberer, mens der indsamles data. Forskydning/kraftsporing kan eksporteres fra den optiske fangstsoftware til Excel, Matlab, Igor eller andre datastyringsprogrammer til yderligere filtrering og analyse. Data, der kan udvindes fra sådanne optiske fangstensemble/bundteksperimenter, omfatter forskellige typer kraftgenereringsprofiler (baseline, rampe/plateau) under varierende analyseforhold, hastighed for kraftgenerering, maksimal kraftgenerering, ensemblekinetisk og trinadfærd gennem trinstørrelser og opholdstider mellem trin eller trin samt pligtforhold. Brugeren kan også ændre analysebetingelserne for at sammenligne, hvordan tilføjelse af forskellige typer myosinmotorer, tilføjelse af actinbindende proteiner eller ændring af bufferbetingelser påvirker disse ensemblekraftgenereringsegenskaber.
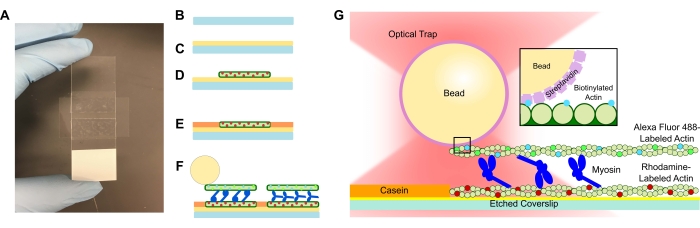
Figur 1: Analyseskema. (A-C) Ætsede dækslips er belagt med poly-L-lysin og bruges til at danne flowcellen ved hjælp af dobbeltsidet tape og et mikroskopglas. Tidsindstillede introduktioner og inkubationstrin beskrevet i protokollen resulterer i rhodamin-mærket phalloidin-stabiliseret actin som skabelon eller bundfilament (D) efterfulgt af kaseinblokering for at forhindre ikke-specifik binding (E) og (F) Alexa Fluor 488 phalloidin-stabiliseret biotinyleret actin som lasten eller topfilamentet og hold af myosin II, der skubber filamenterne fra hinanden og genererer kraft, når ATP introduceres. Motorernes geometri og arten af tværbinding i bundtet kan variere under forskellige forhold, såsom saltkoncentration59. Tidligere undersøgelser har vist, at myosinhaledomænet har evnen til at interagere med actinfilamenter og langsom ensemblemotilitet46. Imidlertid viser myosinhoveder i tunge meromyosineksperimenter binding af hvert hoved til tilstødende actinfilamenter60. (G) Streptavidinperler bruges som det optiske håndtag til fælden og binder udelukkende til lastbiotinyleret actinfilament, hvilket hjælper med at validere, at der dannes korrekte bundter på diaset. Klik her for at se en større version af denne figur.

Figur 2: Fluorescerende actomyosinbundter. Fire forskellige møder med actinfilamenter og bundter inden for bundtetsanalysen præsenteret i figur 1. Den øverste lastbiotinylerede actinfilament med Alexa Fluor 488 phalloidinkanalen vises til venstre, og den nederste skabelon actinfilament med rhodamin phalloidinkanalen er til højre. Nederst er den samme figur vist med farvede linjer overlejret for at hjælpe med at guide øjet. (A) En top actinfilament findes nær en bund actin filament, men har en ufuldstændig overlejring. Dette ville ikke blive brugt til bundteksperimenter. (B) Top- og bundaktinfilamenter er colokaliserede, og intensiteten af hvert filament bekræfter, at de hver især er enkelte filamenter i bundtet. Dette ville være en god kandidat til bundteksperimenter. (C) Et stort bundt selvsamlede rhodaminfilamenter findes på bunden. Mens der er et tilsvarende top actinfilament, der er colokaliseret, er der for mange bundfilamenter til stede; Således ville det ikke blive brugt til bundteksperimenter. Dette er også et eksempel på, hvordan fluorescensintensiteten øges, når flere actinfilamenter af samme type bundtes. Brugeren kan bruge dette som en måler til at bedømme enkeltfilamenter versus bundter af samme filamenttype. (D) Der findes et bundfilament uden tilsvarende topfilament, hvilket heller ikke bekræfter nogen gennemblødning. Dette ville ikke blive brugt til bundteksperimenter. Vi bemærker, at intensiteten af filamenterne i Alexa Fluor 488-kanalen er lav og mener, at det skyldes det filtersæt, der bruges (filtersæt 09 fra Zeiss). Filtersættet, der bruges til rhodaminkanalen, er filtersæt 43 fra Zeiss. Klik her for at se en større version af denne figur.

Figur 3: Myosin II ensemble kraft generation. Repræsentative spor af skeletmyosin II-motorer, der genererer kraft inden for det konstruerede in vitro-actinstrukturelle hierarki. Myosinmotorerne arbejder sammen for kollektivt og produktivt at generere kraft, indtil et plateau er nået, og kraften opretholdes (A) eller oplever antagonisering nær baseline (B). Klik her for at se en større version af denne figur.
Supplerende figur S1: Bruker / JPK Nanotracker2 optisk fælde. (A) Computerskærm. (B) Computertastatur. (C) Computertårn. (D) Controller boks. (E) Laser strømforsyning. (F) Optisk fælde optik boks. (G) Omvendt mikroskop. (H) Dør til mikroskop fase. (I) Polarizer-skyder til at skifte mellem lysfelt og differentiel interferenskontrastbilleddannelse. Klik her for at downloade denne fil.
Supplerende figur S2: Fjernbetjening til optisk fælde. (A) Tastatur til placering af det motoriserede trin. (B-C) Juster fældepositionen. (D) A, X og B tænder og slukker for henholdsvis hovedlukkeren, fælde 1-lukkeren og fælde 2-lukkeren. (E) Logitech-knappen bruges til at vække controlleren. (F) De op- og nedknapper, der bruges til at placere fangstmålet. (G) De op- og nedknapper, der bruges til at placere detektionsmålet. Bemærk, at fjernbetjeningen ikke er påkrævet, og alle disse manipulationer kan udføres i softwaren. Det er dog praktisk at være i stand til at kontrollere målene og scenepositionen, mens man ser på mikroskoptrinmiljøet. Klik her for at downloade denne fil.
Supplerende figur S3: Fluorescensmodul til optisk fælde. 89North PhotoFluor fluorescens hvid lyskilde er koblet til bagsiden af det omvendte mikroskop. Den tændes og slukkes med en vippekontakt (pil). Klik her for at downloade denne fil.
Supplerende figur S4: Fluorescensfilter terningtårn. Tårnet (pilen) kan drejes for at bruge den filterkube, der er nødvendig til billeddannelse i DIC-, rhodamin- eller Alexa Fluor 488-farvestoffer. Bemærk, at filterkuber kan skiftes ud for at tilpasse opsætningen til brug af forskellige fluorophorer. Klik her for at downloade denne fil.
Supplerende figur S5: Nanotracker2 software. (A) Laser tænd / sluk-knap og kontrol. (B) Objektivt positioneringsvindue. Retningspile bruges til at flytte detektionsmålene (øverst) og diffusering (nederst). Dobbeltpile flytter målene med en højere hastighed. Den blå og røde knap nederst til venstre afkobler målene og trækker dem tilbage til deres oprindelige position. Dette er nødvendigt, når der tages prøver ind og ud af mikroskopstadiet. Den tredje knap fra venstre med målene og hængelåsikonet "kobler" målene, så når de både er i fokus og opnår Kohler-belysning, kan brugeren flytte både fangst- og detektionsmålene op og ned i z-aksen. (C) Prøvepositioneringsvindue, der bruges til at flytte mikroskoptrinnet i x- og y-aksen. Dobbeltpile bevæger scenen med en højere hastighed. Dette vindue aktiveres ved at klikke på pil op/ned og venstre/højre pil i topmenuen. (D) Vindue til kameravisualisering. Skruenøgleikonet kan bruges til at indstille tilpassede billeddannelsesbetingelser. Dette vindue aktiveres ved at klikke på kameraikonet i topmenuen. (E) Mikroskop belysning vindue. Dette vindue aktiveres ved at klikke på pæreikonet i topmenuen. Klik her for at downloade denne fil.
Supplerende figur S6: Kalibreringsvindue. A) Dette vindue bruges til perlekalibrering og aktiveres ved at klikke på Cal-ikonet i topmenuen. For at kalibrere en perle opnås den bedste pasform af hjørnefrekvensen i x-, y- og z-signalerne. (B) For hvert signal skal du vælge den relevante signalknap øverst til venstre. (C) Klik på kør og optimer pasformen ved at klikke og trække inden for det grønne vindue (D). (E) Når du er tilfreds med pasformen, skal du klikke på Brug den for følsomhed og stivhed. Dette vil gøre det muligt at registrere forskydning i nanometer og kraft i piconewtons. (F) Klik derefter på Accepter værdier nederst til venstre. Gentag for y- og z-retningen. Klik her for at downloade denne fil.
Supplerende figur S7: Vindue til dataindsamling. Dette vindue bruges til at erhverve positions- og kraftdata og giver brugeren mulighed for at se målingerne i realtid. (A) Dette vindue aktiveres ved at klikke på x, t-ikonet i topmenuen. (B) Brugeren kan skifte mellem visning af x- og y-signalerne. (C) Klik på Start for at begynde at visualisere data. Klik på Autosave for at gemme dataene. Klik på Start Record for at begynde at optage og gemme data. Klik her for at downloade denne fil.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
En in vitro-undersøgelse ved hjælp af optisk pincet kombineret med fluorescensbilleddannelse blev udført for at undersøge dynamikken i myosinensembler, der interagerer med actinfilamenter. Actin-myosin-actin-bundter blev samlet ved hjælp af muskelmyosin II, rhodamin actin i bunden af bundtet og på dækslipoverfladen og 488-mærkede, biotinylerede actinfilamenter på toppen af bundtet. Actinprotein fra kaninmuskel blev polymeriseret og stabiliseret ved anvendelse af generelle actinbuffere (GAB) og actinpolymeriserende buffere (APB). GAB og APB skal være frisklavede hver dag i laboratoriet ved hjælp af ATP, FC buffer og TC buffer. Muskelmyosin II blev brugt til at danne actin-myosin-actin sandwich. Phalloidin blev brugt til fluorescerende farvning af actinfilamenterne samt stabilisering in vitro.
Myosinaktivitet kan bekræftes ved at udføre et standard glidefilamentassay som offentliggjort tidligere46,47. Myosin II og dets underfragmenter kan binde til dækslipoverfladen i forskellige retninger, og tilstedeværelsen af haledomænet kan bremse filamentglidning sammenlignet med assays ved hjælp af tung meromyosin46,48,49. Glidende og overfladebevægelse kan dog stadig observeres. En mere tydelig demonstration af myosinaktivitet er aktiv actinfilamentbrud, der kan observeres, hvor længere actinfilamenter brydes i mindre fragmenter, der derefter glider væk i flere retninger. Dette sker på grund af den høje koncentration af aktive motorer på overfladen, er blevet observeret af flere laboratorier og forekommer ikke uden aktive myosinmotorer til stede 42,50,51,52,53,54. Endvidere hjælper bundtanalysen, der præsenteres her, med at lindre bevægelighedsproblemer, der primært har været forbundet med glidefilamentanalysen, såsom de forskellige motorbindingsretninger på en glasdæksel, fordi bundtassayet involverer kaseinblokering af glasoverfladen, så motorer binder inden for bundtet 47,55,56.
Det første skridt er at tilføje rhodamin actin filamenter som bunden eller skabelonfilament til en poly-L-lysin belagt coverslip i en flowcelle. Poly-L-lysin bruges til at fremme actinbinding, da poly-lysin er positivt ladet, mens actin har negative ladninger og er blevet anvendt i tidligere cytoskeletale in vitro-assaypræparater61,62,63. Før bundtdannelse blev forskellige actinfortyndinger tilsat til en flowcelle for at optimere actinkoncentrationen. I dette tilfælde var 600x fra bestanden den optimale fortynding, der gav et tilstrækkeligt antal skabelonfilamenter til bundtdannelse, men med tilstrækkelig afstand, så bundter blev individualiseret. Fortynding blev udført ved hjælp af APB-bufferen. Tilsætning af rhodamin actin blev efterfulgt af et lag kasein for at blokere overfladen og undgå ikke-specifik binding. Flowcellen blev inkuberet i 30 minutter og vasket efter inkubation med buffer for at vaske eventuelle ubundne actinfilamenter ud. Endelig blev en kombination af myosin, 488 / biotin actin og streptavidin-belagte perler tilsat til flowcellen for at lette dannelsen af actin-myosin-bundt. Perlekoncentrationen skal være sådan, at der er nok til at binde overfladebundter og nok i suspension til at lette kalibrering. Imidlertid kan for høj perlekoncentration forårsage vanskeligheder under fangsteksperimenter på grund af naboperler, der falder ned i laserfælden og forstyrrer målingen. Myosinmotorer tilsættes til kombinationen lige før injektion til diaset, så myosinmotorerne ikke præventivt aggregeres med lasten eller det øverste biotinylerede actinfilament og dermed binder den nederste rhodamin til bundt biotinylerede actinfilamenter.
Det optiske optiske fældesystem NT2 er en kommerciel optisk fælde med kombineret lysfelt, differentiel interferenskontrast (DIC) og epifluorescensbilleddannelsesmetoder. Det er kombineret med et Zeiss AxioObserver 3 omvendt mikroskop med 100x / NA 1.46 og 63x / NA 1.0 vandnedsænkning og detektion mål. Systemet er udstyret med klik- og trækfangstfunktion for en laserfælde og kan bruges under billeddannelse i en hvilken som helst af de tidligere anførte modaliteter. De dannede bundter detekteres og bekræftes ved anvendelse af fluorescensbilleddannelse. At have en hvid lyskilde med passende filterkuber (GFP / FITC og TRITC / CY3) giver mulighed for hurtig skift mellem filamentbilleddannelse. Colokaliserede AF'er blev verificeret ved at visualisere AF'erne ved de forskellige excitationsbølgelængder, før de tog hver kraftmåling ved hjælp af optisk pincet. Da filamenterne hurtigt kan fotoblege, selv med et iltrensningsreagens, foreslås det, at forskere optimerer visualiseringsparametre som intensitet og eksponeringstid, før de udfører bundteksperimenterne.
Optisk fangst blev anvendt til at foretage kraftmålingerne ved hjælp af streptavitinperlerne i nærværelse af ATP til at binde det biotinylerede lastactinfilament og aktivere myosinkraftgenerering som en krafttransducer. Forskydnings- og kraft- versus tidsdata opnået ved optisk fangst blev ekstraheret fra fangstsoftwaren til analyse. Den kommercielle fangstsoftware giver dog også analyserutiner, der kan bruges, eller brugerdefinerede algoritmer i andre programmer kan programmeres af brugeren til at visualisere og analysere fangstdata. På brugerdefinerede optiske fangstsystemer kan brugeren have excitationslasere i stedet for en hvid lyskilde med filtre, som også er acceptable at bruge. Desuden kan fluorescerende farvestoffer ændres, så de passer til det eksisterende udstyr, som en bruger måtte have, hvis emissionsspektrene ikke overlapper hinanden og forårsager blødning.
Vi bemærker, at det præsenterede assay er et baseline-assay, der kan tilpasses yderligere af brugeren afhængigt af deres forskningsspørgsmål inden for actomyosin-ensemblemekanik. Den generelle arbejdsgang kan også anvendes på andre in vitro cytoskeletale ensemblesystemer, der kan være af interesse, såsom mikrotubulusbundtassays, der danner minimale modeller af mitotisk spindel 32,61,63,64,65,66. Ændringer kan omfatte, men er ikke begrænset til, ændring af fluorophoretiketter, der passer til brugerens eksisterende opsætning; ændring af myosinkoncentration, konstruktion eller isotype; og titrerende bufferbetingelser, blandt andre aspekter.
Potentielle udfordringer er mulige, når du udfører denne analyse. Ved dannelse af actin-myosin-bundterne er myosinkoncentrationen i actinbundterne muligvis ikke homogen på tværs af diaset. For at imødekomme dette måles flere bundter på tværs af hele diaset for at sikre, at motorfordelings- og kraftgenereringsprofiler er korrekt samplet. Det er også udfordrende at kende bundtorientering, hvis dette er nødvendigt for fortolkning af kraftdata. Således skal der tages flere forsøg for hvert bundt. Man kunne også inkorporere actinfilamentendemærkning gennem fluorescerende gelsolin eller gelsolinbelagte perler af mindre størrelse end det optiske fangsthåndtag. Fluorescensbilleddannelse kan også bruges til at se på x- og y-komponentkræfter til at udlede bundtorientering. Da myosinaggregeringstilstanden desuden er stærkt påvirket af bufferens ionstyrke med dannelse af tykke filamenter, der forekommer ved hurtig fortynding af KCl, bør buffersaltkoncentrationen overvåges på passendevis 67,68.
Tidligere undersøgelser, der brugte andre in vitro-metoder såsom glidende assays, var nyttige til at identificere myosindomænernes rolle og studere konfigurationen og interaktionerne mellem myosin og andre actinbindende proteiner. Disse metoder har imidlertid en ulempe, idet binding af myosin på en stiv overflade vil begrænse potentialet for koordinering mellem myosinmotorer og dermed mekanosenserende feedback, der opstår for at afgøre, om motorensemblet er i en høj eller lav driftsforholdstilstand 33,35,41,69. Endvidere giver optisk fangst med single-myosin-motornetværk ikke en klar forståelse af, hvordan myosinmotorer interagerer med hinanden og med actinfilamenter. Protokollen, der er udviklet her, giver mulighed for undersøgelse af myosinmotorensembledynamik inden for et kompatibelt, hierarkisk actin-netværk. Det kan også tilpasses med hensyn til motorfilamentensembleegenskaber såsom koncentration, isoform og buffermiljø, blandt andre aspekter, for at muliggøre systematisk undersøgelse. Den præsenterede protokol er en platform for fremtidige undersøgelser af mere komplekse actomyosinnetværk og opretholder præcisionen af forskydnings- og kraftgenereringsmålinger lettet af optisk fangst, der traditionelt er blevet brugt til enkeltmolekyleundersøgelser.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ingen interessekonflikter at erklære.
Acknowledgments
Dette arbejde støttes delvist af University of Mississippi Graduate Student Council Research Fellowship (OA), University of Mississippi Sally McDonnell-Barksdale Honors College (JCW, JER), Mississippi Space Grant Consortium under tilskudsnummer NNX15AH78H (JCW, DNR) og American Heart Association under tilskudsnummer 848586 (DNR).
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
