

Summary
이 문서는 박테리오파지 람다의 감염의 휘황 라벨이 지정된 버전을 준비하기위한 절차를 설명합니다
Abstract
박테리오파지 (파지) 람다와 박테리아 E.를 포함하는 시스템 대장균은 오래 세포의 운명 결정 1,2의 패러다임을 역임했다. phages의 숫자로 셀의 동시 감염에 이어 두 경로 중 하나가 선택되어 lytic (악성) 또는 lysogenic (휴지) 3,4. 우리는 최근 휘황 개인 phages 라벨을위한 방법을 개발, 개인 phages와 셀 (5)의 수준에서 현미경으로 실시간으로 후 감염 결정을 검토 할 수 있었다. 여기, 우리는 이전 일 5에 설명 된 감염 실험을 수행하기위한 전체 절차를 설명합니다. 이 형광 phages의 창조, 세포의 감염, 현미경 이미징 및 데이터 분석을 포함하고 있습니다. 형광 파지는 capsid gpD 단백질의 "하이브리드"공동 표현 야생 유형과 YFP 퓨전 버전입니다. 원유 파지의 lysate 먼저 gpD-EYFP (Enh의 lysogen을 유도함으로써 얻을 수 있습니다플라스미드 표현 야생 유형 gpD을 품고, 옐로우 형광 단백질) 파지를 anced. 정화 단계의 시리즈는 다음 현미경으로 DAPI-라벨 및 이미징 다음, 수행됩니다. 이것은 균일, DNA 포장 효율성, 형광 신호와 파지 주식의 구조 안정성을 확인하기 위해 수행됩니다. 박테리아에 phages의 초기 흡착는 ° C 바이러스 DNA 주입 6 실행하는 35 짧은 부화에 의해 다음 다음에 얼음이 수행됩니다. 파지 / 박테리아 혼합는 다음 얇은 영양 한천 판, coverslip으로 덮여 있으며 epifluorescence 현미경 이미징의 표면으로 이동됩니다. 후 감염 과정은 10 분 간격으로, 4 시간에옵니다. 여러 단계 순위는 ~ 100 셀 감염이 단일 실험에서 추적 할 수 있도록 추적합니다. 각 위치와 시간 시점에서, 이미지가 위상 대비 빨간색과 녹색 형광 채널에 인수되어 있습니다. 위상 대비 이미지가 자동 cel 나중에 사용됩니다형광 채널이 감염 결과를 특성화하는 데 사용하는 동안 난 인정 : 재개 세포의 성장과 분열에 이어 lysogeny 요소의 세포 용해, 또는 표현 (빨간색)에 이어 새로운 형광 phages (녹색)의 생산. 인수 시간 경과 영화는 수동 및 자동 방법의 조합을 사용하여 처리됩니다. 각 감염 이벤트 (phages을 감염 예 수와 위치)뿐만 아니라 감염 결과 (용해 / lysogeny)에 감염 매개 변수의 식별의 데이터 분석 결과. 원하는 경우 추가 매개 변수를 추출 할 수 있습니다.
Protocol
1. 원유 파지의 lysate의 창조 (그림 1)
- 50 ML 플라스크에 LE392의 새로운 식민지 (λ LZ1) [pPLate * D] (자세한 내용은 표 1 참조) 10 μg / ML의 Kanamycin과 100 μg / ML 암피실린과 보충 LB 매체 7 6 ML에의 예방. ° C 온화한 진동 (180 RPM)에 30에서 하루 아침에 성장.
- 에겐 IBM (LB 10 MM MgSO 4 보충)로 문화 1:100을 희석하고 부드러운 흔들림 (180 RPM)와 30 ° C에서 성장합니다. 파지 수율을 최적화하기 위해, 문화 볼륨이 더 이상 술병 볼륨 용량 둘은 확실히보다 수 없습니다 있는지 확인하십시오. 우리는 일반적으로 2 리터 또는 2.5 리터 용량의 두 플라스크를 준비하고, 각각의 플라스크에 250 ML의에겐 IBM 매체에 2.5 ML 밤 문화를 추가 할 수 있습니다.
- 셀 밀도는 OD 600에 도달하면 ≈ 0.6는 (~ 2.5-3 시간), incubat 후 가벼운 흔들림 (180 RPM)와 18 분에 42 ° C 물 목욕 셰이커에 문화를 이동하여 lysogen을 유도하고,37 번 E ° C 용해 될 때까지 약한 흔들림 (180 RPM)를 볼 수 있습니다 (문화 ~ 60에 분명해진다 - 90 분).
- 문화에 2 %의 클로로포름을 추가 실온에서 15 분에 품다 후 혼합 할 손으로 흔들어합니다. 주의 : 클로로포름을 처리하는 장갑을 착용하고, 숨을 피할 수 있습니다.
- 이 250 ML의 원심 분리기 병에 문화를 전송, 4 ° C.에 15 분에 10,000 RPM에서 Sorvall GSA 회에 문화를 원심 분리기 파지 입자를 포함하는 표면에 뜨는를 복구하고, 쓰레기의 펠릿을 폐기합니다. 보이는 파편을 제거해야 할 두 번째 원심 분리를 수행합니다.
- 파지 농도를 측정하는 표준 파지 적정 프로토콜 팔을 사용합니다. 파지의 titer는 다음과 같아야합니다 ~ 5-10 X 10 9 PFU / ML. 등 때문에 형광 파지의 유전자형에 Sam7 변이의 지표 변형 등의 LE392로 supF 변형을 사용하여 (FI 큰 액자를 얻기 위해 다양한 NZYM으로 만든 상단 한천과 한천 플레이트를 사용하여gure 2).
2. 파지 정화 (그림 1)
- lysed 박테리아에서 해방 핵산을 소화하기 위해 lysate에 DNase I 및 RNase (1 μg / ML 각각)을 추가, 대형 (예 : 2 리터) 병에 lysate를 넣어하고, 실온에서 1 시간을 품다.
- lysate에 1M NaCl을 추가, 250 ML의 원심 분리기 병에 lysate를 전송하고, 얼음에 3 시간을 품다. 4에 15 분에 10,000 RPM에서 Sorvall GSA의 lysate를 원심 분리기 ° C. 표면에 뜨는을 복구 할 수 있습니다. 파지의 titer는 원유 lysate의 그것과 유사합니다 ~ 5-10 X 10 9 PFU / ML. NaCl의 첨가 세균의 잔해로부터 파지 입자의 분리를 촉진하고 PEG (8)에 의해 파지 입자를 효율적으로 강수량이 필요합니다.
- 2 리터 병 예를 들어, 큰 병에 lysate를 부어 lysate에 10 % (w / V) PEG8000을 추가 천천히 저어 또는 실온에서 PEG8000를 해산하기 위해 흔들. 250 ML의 퍼센트로 lysate를 전송rifuge 병은 다음 4 ° C.에서 (~ 16 시간) 밤을 품다 4에 15 분에 10,000 RPM에서 Sorvall GSA 회에 lysate을 원심 분리기 ° C. 표면에 뜨는 폐기하십시오.
- 파지 SM 버퍼 (초기 파지의 lysate 250 ML 당 4 ML SM 버퍼)와 펠렛을 (파지 입자가 PEG8000을 유발) 만끽 해보세요. 매우 흔들 온화한 나 더는 4 ° C. 16 시간에 대한 흔들림없는과 품다
- 부드럽게 50 ML Eppendorf 원심 분리기 튜브에 lysate를 (파지 입자와 SM 버퍼)을 받아 다음 0.5로 나머지 펠렛을 씻으 - SM 버퍼 1 ML 있습니다.
- lysate에 클로로포름의 동등한 볼륨을 추가합니다. 부드럽게 몇 번 반전 위쪽 및 아래쪽으로 클로로포름으로 lysate를 섞는다. 15 Eppendorf 5804R 4 ° C에서 분 또는 이와 유사한 벤치 톱 원심 분리기에 4,000 rpm으로 원심 분리기.
- 명확하게 lysate를 얻을 단계 2.6를 반복합니다. 파지의 titer는 ~ 1-2이어야 X 10 11 PFU / ML.
- 1.3 g / ML, 1의 세 가지 다른 밀도 (ρ)와 SM / CsCl 솔루션을 준비0.5 g / ML 및 1.7 g / ML. 보다 정확한 밀도 읽기를 얻기 위해 굴절률 (η)을 측정합니다. 밀도 변환 9 ρ입니다 = 10.8601 η - 25 13.4974 ° C. 자세한 내용은 표 3을 참조하십시오.
- 14 ML 초 명확한 Beckman 40Ti의 ultracentrifuge 관에 솔루션을로드하는 긴 바늘과 주사기를 사용하십시오. 혼합 방지하고 더 나은 밀도 기울기를 형성하기 위해 솔루션을 사용해야합니다 (다른 하나에 증가 밀도 즉, 레이어링 솔루션)는, 즉 부드럽게 1.3 g / ML, 1.5의 순서에 SM / CsCl 솔루션 2 ML을로드 underlaying g / ML 및 1.7 g / 튜브의 하단에 3 ML의 주사기와 바늘을 삽입하여 ML.
- 부드럽게 14 ML의 ultracentrifuge 관의 상단에서 오버레이하여 파지의 lysate 8 ML을로드합니다. 밸런스 튜브를 준비합니다. 4에서 4 시간에 24,000 rpm으로 Beckman SW40Ti 회에 원심 분리기 ° C.
- 부드럽게 AF를 사용하여 어두운 방에 튜브를 꺼내 검은 배경 튜브의 상단에서 조명lashlight. 파지 밴드 1.3 g / ML 및 1.5 g / ML SM / CsCl 레이어 (그림 3A) 사이의 인터페이스의 위치에서 명확하게 볼 수 있어야합니다. 약간 3 ML의 주사기와 21.5 게이지 바늘로 밴드 아래에있는 관의 측면을 통해 구멍. 부드럽게 파지의 정지 ~ 500 μl를 수집합니다. 파지의 titer는 ~ 5-10 X 10 11 PFU / ML해야합니다.
- 4 ML 매우 명확 Beckman의 ultracentrifuge SW60Ti 회 관으로 파지 정지를 놓습니다. 1.5 g / ML SM / CsCl 솔루션 관을 입력합니다. 밸런스 튜브를 준비합니다. 4에서 24 시간에 35,000 rpm으로 Beckman SW60Ti 회에 원심 분리기 ° C.
- 보이는 밴드에서 파지를 수집하는 단계 2.11에서와 같이 동일한 절차를 반복합니다. 그림 3B에 도시 된 바와 같이, 밴드가 표시됩니다.
- 투석 막 카세트 (표 2)에 파지 현탁액을로드하고 3 시간, 3 시간 및 overnig의 기간에 대해 4에 SM 버퍼의 1000 배 볼륨 ° C에 대해 세 번 dialyzeHT (~ 16 시간). 투석의 목적은 파지 정지에 CsCl 존재를 제거하는 것입니다. 최종 파지의 titer는 ~ 5-10 X 10 11 PFU / ML해야합니다.
3. 한 아가로 오스 겔 판 (그림 4) 준비
- 70 % 에탄올로 6 현미경 슬라이드를 (75 X 50mm, 두께 1mm) 청소하십시오.
- 5 슬라이드를 정렬하고 그림 4와 같이 테이프로 고정하십시오.
- 집착 랩 (1.5 % 아가로 오스를 항복)으로 덮여 작은 비커 6 ML 매체에 0.09 g 아가로 오스를 섞는다. 솔루션은 분명집니다 때까지 핫 플레이트에 가열.
- 보안 슬라이드로 아가로 오스 솔루션을 넣어.
- 조심스럽게 기포를 피 상단에있는 마지막 슬라이드를 놓습니다. 장소 위에 무게와 ~ 30 분 동안 냉각 할 수 있습니다.
- 측면에 4 슬라이드를 제거하고, 집착 랩으로 상단과 하단 슬라이드와 함께 슬라브을 포장. 슬래브는 최대 3 일에 대해 4 ° C에 저장할 수 있습니다.
4. 정화 파지 주식 테스트
- 준비위에 설명 된대로 PBS-아가로 오스 겔 판 (제 3 항).
- DAPI로 정화 파지를 청바지. 10 파지의 μl (~ 1 개 10 10 PFU / ML) 10 μg / 4에서 30 분 동안 배양 ML DAPI (5 μg / ML의 최종 DAPI 농도), 10 μl로 ° C 또는 실온에서 10 분.을 혼합
- 제 1 24 미리 준비된 PBS-아가로 오스 판의 X 50mm coverslip, 오버레이 작은 조각 (~ 10 × 10 ㎜)의 중심에있는 파지 / DAPI 혼합물의 장소 1 μl. 아가로 오스 판의 작은 조각은 샌드위치 젤이 제거의 상단 슬라이드 후 면도날로 절단됩니다. YFP 및 DAPI 채널을 통해 epifluorescence 현미경 이미지 샘플을. 개인 phages는 두 채널 모두에서 회절 - 제한된 형광 "장소"(그림 5)와 같은 표시됩니다. 단계 아래 6.2에서와 같은 현미경과 카메라 설정을 사용합니다.
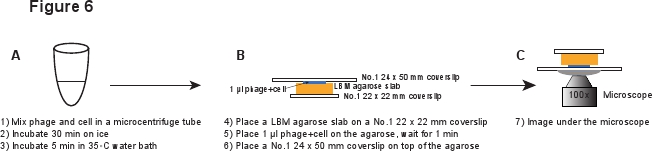
5. 감염 (그림 6)
- 14 ML 팔콘 튜브에 LE392의 새로운 식민지의 예방 [PP RE-mCherry] (자세한 내용은 표 1 참조) 100 μg / ML 암피실린, 10 MM MgSO 4 0.2 % 말토오스로 보충 LB 매체 2 ML에. 적당한 흔들림 (265 RPM)와 37 ° C에서 밤새 성장.
- LBMM (LB 10 MM MgSO 4 0.2 % 말토오스와 보충)로 문화 1:1000 희석, 즉 50 ML 플라스크의 5 ML LBMM 중간에 5 μl 밤 문화를 추가 할 수 있습니다. ° C 적당한 진동 (265 RPM)와 37 OD 600 ≈ 0.4로 성장합니다.
- 제 위의 3에 설명 된대로에겐 IBM - 아가로 오스 겔 판을 준비하는에겐 IBM 매체를 사용하십시오.
- 상온에서 2 분을위한 벤치 탑 microcentrifuge에 2,000그램에서 세포의 1 ML을 원심 분리기. 표면에 뜨는을 제거하고 부드럽게 OD 600 & 20에 도달하는 20 μl 얼음처럼 차가운 LBMM에 세포를 resuspend.
- 정화 파지 주식을 조작하면, 다양한 피펫 팁을 사용하거나 끝이 파지 입자 3 전단 방지하기 위해, 더 개방하기 위해 일반 피펫 팁을 잘라. 부드럽게 MIX 0.1의 범위에서 평균 phages - 투 - 셀의 비율에 도달하기 파지를 정화 20 μl와 세포의 20 μl - 5. 파지 흡착 할 수 있도록 30 분에 얼음을 품다 후 파지의 DNA 주입 6 실행하는 데 5 분에 35 ° C 물 목욕에 품다.
- 모든 세포 응집체를 분리 할 수있는 몇 번 아래로 피펫합니다. 다시 phages을 전단 방지하기 위해 다양한 피펫 팁을 사용합니다. LBMM, 45 μl LBMM로 예를 들면 5 μl 혼합에 혼합 1시 10분을 희석.
- 제 1 22 X 22mm의 coverslip에에겐 IBM - 아가로 오스 판 (~ 10 × 10 ㎜)의 조각을 놓으십시오. 아가로 오스 판은 실내 온도에 도달하기 위해 사용하기 전에 미리 준비에겐 IBM - 아가로 오스 판은 최소한 1 시간 동안 실온에서 배치해야합니다. 장소 1 아가로 오스 판에있는 파지 / 세포 혼합물의 μl와 혼합는 아가로 오스 판으로 흡수 할 수 있도록 1 분 기다립니다. 부드럽게 아가로 오스 판의 상단에 1 위 24 X 50mm의 coverslip를 놓습니다. 이 절차는 INF에서 phages을 전단 방지하기위한 것입니다ected 셀 (그림 6).
6. 현미경으로 세포의 운명을 따라
- 현미경의 무대에 coverslip을 자세히 탑재합니다. 이미징를 들어, 높은 배율 (예 : 100x) 목표를 (아래 토론에서 현미경 시스템을 참조)을 사용합니다.
- 초기 기간에 대해 설정 한 이미지를 취득. 이 이미지 세트는 모든 감염 phages의 초기 번호와 위치를 특성화하는 데 사용됩니다. 200 nm의 Z-축 (세로) 간격으로 일련의 15 이미지보십시오. YFP 채널을 통해 이미지. 또한, 위상 대비, mCherry 채널을 통해 하나의 포커스에있는 이미지를 가져. (아래 토론에서 이미지 수집 참조) 표백 및 세포 손상을 최소화하면서 충분한 신호를 얻기 위해 빛의 세기 및 노출 시간을 최적화합니다.
- 후 감염 세포 운명의 시간 경과 영화를 취득. 위상 대비, YFP와 mCherry의 이미지 샘플을주변 4 시간 10 분의 시간 간격으로 채널. 시간 경과 영화 동안, 표백 및 광독성으로 이어질 수 샘플의 불필요한 노출을 방지하기 위해, 시점에 따라 채널 당 하나의 Z-위치 이미지를 사용합니다.
7. 이미지 분석
- 수동으로 phages하고 초기 시간 프레임에 기록 파지 위치와 세포 길이의 수를 계산합니다. 이것은 MetaMorph 또는 ImageJ와 같은 소프트웨어를 사용하여 수행 할 수 있습니다. 세포 운명 (lytic lysogenic 또는 감염되지 않은), 용해 시간 및 시간 경과 영화를 재생하여 원하는 다른 정보를 기록합니다. 다른 세포의 운명을 확인하려면, 아래의 대표 결과 섹션에서 시간 경과 영화를 참조하십시오.
- 위의 수동 분석뿐만 아니라,보다 양적 정보 (개별 셀에 시간이 지남에 따라 예를 들어 형광 레벨) 자동화 된 세포 인식 및 혈통 추적 알고리즘을 사용하여 추출 할 수 있습니다. 우리는 TR을위한 집 지어 MATLAB 프로그램을 사용하여셀 세분화에 대한 Schnitzcell MATLAB 코드 (칼 테크에서 Elowitz 그룹에 의해 작성된)와 함께 세포 계보 (Lineage) 및 형광 수준을 acing.
8. 대표 결과 :
파지 도금 :
휘황 표시 phages (단계 1.6 제 2)의 액자는 야생 타입 (그림 2)보다 훨씬 작습니다. 따라서 우리는 액자에 대한 ° C 배양기가 볼 수 있도록 37 판 할 최소한 12 시간을 품다.
파지의 Ultracentrifugation :
CsCl 단계 기울기 (단계 2.10)과 파지 샘플의 ultracentrifugation 후, 두 그룹은 (그림 3A) 표시됩니다. 상단 밴드는, 파지 정지 및 SM / CsCl 1.3 g / ML 층 사이의 인터페이스에서, 세포 파편과 빈 파지의 capsids가 포함되어 있습니다. 하단 밴드는, SM / CsCl 1.3 g / ML 및 1.5 g / ML 층 사이의 인터페이스에서 파지 밴드입니다. 티S 밴드는 형광 파지 λ LZ2을위한 녹색 나타납니다. 야생 유형 파지 λ IG2903의 밴드는 5 푸른 나타납니다. 단계 2.12에서 CsCl 평형 기울기의 ultracentrifugation 한, 파지 밴드는 관 (그림 3B)의 중간 부분에 표시됩니다. 형광 파지 λ LZ2는 gpD - EYFP 및 gpD capsids, 단백질 - 투 - DNA의 비율이 혼합되어 있기 때문에 야생 유형의보다 높은 것입니다. 따라서, 형광 파지 λ LZ2의 밴드는 야생 유형 λ IG290310보다 (튜브에서 더 높은 위치에있는 것 같습니다) 약간 가볍다.
DAPI의 얼룩 :
그림 5는 DAPI (제 4 항)와 파지의 라벨을 한 후 얻은 전형적인 이미지를 보여줍니다. 성공적으로 정화 파지의 YFP와 DAPI 신호에 가까운 100 % 대응이 있어야합니다. 우리는 일반적으로 관찰 그 YFP 현장의 1 % 미만s은 (는) DAPI을 (바이러스 성 게놈없이 capsids을 대표하는)를 포함하지 않습니다. DAPI 명소의 1 % 미만이 YFP (비 형광 phages에 해당) 5를 포함하지 않습니다.
시간 경과 영화 :
Lytic 세포는 세포 용해 다음 세포 내부 YFP 형광 (녹색 채널)의 축적에 의해 인정 받고 있습니다. Lysogenic 셀은 셀 정상적인 세포의 성장과 분열의 재개 내부의 균일 한 mCherry 형광 (적색)의 축적에 의해 인정 받고 있습니다. 감염되지 않은 세포 (또는 감염이 실패 세포) 위의 phenotypes 중 하나를 표시하지 않으며 성장하고 정상적으로 나눕니다. 그림 7 단계 대비 몇 이미지 세트를 보여줍니다, YFP와 mCherry 채널, 이들의 해당 겹쳐 이미지 전형적인 시간 경과 영화 (제 6 항)에서 세 채널. 개인 phages (녹색 부분)은 초기 시간 프레임 (그림 7A)에서 명확하게 볼 수 있습니다. 일반적으로, 수다른 phages가 unadsorbed하는 동안의 phages가 그림 7B에 (왼쪽 패널)에 도시 된 바와 같이, 세포 표면 (아마도 이러한 세포를 감염)에 볼 수 있습니다. 감염 결과는 시간이 지남에 따라 구별된다. lytic주기는 세포 용해 (발표 녹색 phages으로 폭발 세포, 그림 7D)에 이어 새로운 phages의 세포 생산 (녹색, 7C 그림)로 표시됩니다. Lysogeny는 P의 RE 프로모터 (빨강, 그림 7C)과 세포 성장과 부문 (빨강, 그림 7D)의 재개에서 mCherry의 생산에 의해 표시됩니다.




1 그림. AB)를 품고, gpD-EYFP 파지의 lysogen을 유도함으로써 얻을 수 있습니다. 파지는 단계 (패널 CL)의 일련의 정화입니다.

그림 2. 파지의 액자. 형광 파지의 액자가 (왼쪽) 37에서 12 시간에 번호판을 잠복기 후 야생 유형 (오른쪽)의보다 작은 ° C. 아르

그림 3. 파지 밴드 ultracentrifugation 후. A) 두 밴드는 CsCl 단계 기울기에 ultracentrifugation 후 볼 수 있습니다. 상단 하나는 세포 파편과 빈 파지의 capsids에 해당, 하단 밴드는 원하는 파지가 포함되어 있습니다. 왼쪽 : 형광 파지, 오른쪽 :. 야생 타입 B ) A 단일 파지 밴드는 CsCl 평형 기울기에 ultracentrifugation 후 볼 수 있습니다. 형광 파지 밴드는 (왼쪽) 야생 타입 파지 (오른쪽)의 푸른 밴드에 비해 녹색입니다.

그림 4. 아가로 오스 겔 슬라브를 준비 절차.

그림 5. DAPI의 얼룩 후 phages의 형광 이미지. 개인 phages 쉽게 구별하고, YFP하고 DAPI 잘 공동 현지화 신호.

그림 6. 파지 감염 및 이미징 설정의 도식 설명. 이 이미지의 전체 크기 버전을 보려면 여기를 클릭하십시오.
gure 7 "SRC ="/ files/ftp_upload/3363/3363fig7.jpg "/>
그림 7. 파지 감염의 시간 경과 영화의 전형적인 이미지. 표시하면 단계 대비 있으며, YFP와 mCherry 채널뿐만 아니라,이 세 채널의 오버레이. 초기 시간 프레임에서 (A) YFP 채널 이미지. 왼쪽, 다른 Z-위치에서 YFP 이미지의 합계. 세 오른쪽 이미지는 다른 Z-위치, 세포 표면의 서로 다른 영역에 해당하는에 샘플 YFP 이미지입니다. (B), (C)와 (D) 상 대비 (중간 왼쪽), 다른 시간 프레임에서 YFP (가운데 오른쪽)과 mCherry (오른쪽) 채널의 중첩 이미지 (왼쪽). (B) t = 0에서 두 개의 셀이 볼 수 있으며, 하나의 파지 (녹색 부분), 그리고 한 셀에 감염 각각 3 phages에 감염되어 있습니다. 또한 일부 unadsorbed phages는 아르 관찰. (C) t = 80 분에서 한 phages에 감염 두 세포가 각 lytic 경로에 갔을 등 표시새로운 phages의 세포 생산 (녹색)에 의해 D. 3 phages에 감염 세포는 예약 발기인 (빨간색)에서 mCherry의 생산에 표시된 lysogenic 경로로왔다. lysogenic 셀 §을 나누어 반면 (D) t = 2 시간에서 lytic 경로는 세포 용해 (셀 폭발)이 발생했다.
Subcellular 수준에서 만들기 그림 7의 § 왼쪽 패널 (C)와 (D) 젱, 사무엘 O. 스키너, Chenghang 종, 장 Sippy, 마이클 Feiss, 그리고 이도 골딩을 Lanying, 세포, 141에서 재 있으며, 결정은 결과를 결정합니다 박테리오파지의 감염, 682-691, Elsevier의 허가와 저작권 (2010).
| 이름 스트레인 | 관련 유전자형 | 소스 / 참조 |
| 박테리아 변종 | ||
| LE392 | 저녁을 먹다에프 | 존 Cronan, 일리노이 대학교 |
| 파지의 긴장 | ||
| λ LZ1 | gpD - EYFP, cI857 Sam7 D-eyfp B :: kanR | 외. 5 젱 |
| λ LZ2 | λ LZ1로 gpD - 모자이크, 같은 유전자형 | 외. 5 젱 |
| Plasmids | ||
| PP RE - mCherry | P RE의 통제하에 mCherry, A R | 외. 5 젱 |
| pPLate * D | λ 후반 발기인의 통제하에 gpD, A R | 외. 5 젱 |
표 1. 세균 변종,이 작품에 사용 phages와 plasmids.
| 밀도 ρ (g / ML) | CsCl (g) | SM (ML) | 굴절률 η |
| 1.30 | 39 | 86 | 1.3625 |
| 1.50 | 67 | 82 | 1.3815 |
| 1.70 | 95 | 75 | 1.3990 |
표 3. CsCl 솔루션은 단계 기울기에 대한 SM 버퍼 (100 ML)에 준비.
Discussion
박테리아 변종, 파지 및 Plasmids :
스트레인 LE392은 supF입니다. 그것은 (자세한 내용은 표 1 참조) 파지의 게놈에 Sam7 돌연변이를 억제하기 위해 선정되었습니다. 따라서, 유도 lysogens 결국 lyse과 파지 입자를 공개, lytic 경로를 선택으로 의지 감염된 세포합니다. Lysogenic 세포가 30에서 재배 ° C 파지의 게놈에서 857 allele CI 민감한 온도의 존재로 인해. 열 유도 후, gpD - EYFP과 야생 유형 gpD는 λ LZ1의 게놈과 플라스미드 pPlate * D 각각에서 공동으로 표현됩니다. 그 결과, 새로 만든 파지 λ LZ2의 capsid는 gpD - EYFP 및 gpD 단백질의 혼합물이 포함되어 있습니다. 이 모자이크 파지는 개별 phages 5 감지를 허용 구조적으로 안정적이고 충분히 형광입니다. PP RE - mCherry는 lysogenic pathwa의 선택을 검색하는 데 사용 플라스미드 기자입니다Y. 발기인 P의 RE는 lysogeny 1,11의 설립시 CII에 의해 활성화됩니다. PP RE - mCherry 5 mCherry 12 GFP를 교체하여 PE-GFP 11에서 파생되었다. 자세한 내용은 이전 작품 5를 참조하십시오.
성장 조건 매개 변수 :
lysogen 유도 동안 (제 1), 180 rpm에서의 가벼운 흔들림은 좋은 바이러스 수율이 13을 제공합니다. 포도당 신진 대사로 피해야한다 성장 매체에 포도당의 이용은 산성 신진 대사 제품을 생성하고, 성숙 람다 입자는 산성 pH의 13 살 불안정합니다. MgSO 4의 추가는 파지 capsid 3 안정화 목적으로합니다. 야생 유형 CI를 (대신 CI 857의) 수행 phages를 들어, lysogen은 DNA-손상 에이전트 미토 마이신 C 3을 사용 유도 할 수 있습니다. 단계 1.3에서, 37의 부화는 ° C는 일반적으로 90 분 초과 할 수 없습니다. 이 usef입니다 매 30 분 OD 600까지 셀 밀도를 확인 UL. 좋은 lysate를 들어, OD 600 주변에 0.2 이하로 들어가게되고, 나머지 OD 600 세포 파편의 결과입니다. 새로 만든 파지는 세포 파편에 자신의 DNA를 삽입하기 시작 수 있으므로 너무 오래 잠복기는 낮은 파지 수율 될 수 있습니다. 단계 2.11과 2.13에 보이는 파지 밴드를 (적어도 1 개 10 11 파지 입자)을 획득하려면, 단계 1.2에서 적어도 500 ML 문화를 성장. 단계 5.1 및 5.2의 성장 매체에 0.2 % 말토오스의 추가는 양고기, 파지 람다 흡착 3,14에 대한 수용체의 표현을 유도 목적으로합니다. mCherry - 대신 단계 5.2의 100 배의 1,000 배 희석은 기자 플라스미드 PP의 RE에서 mCherry 배경 수준을 감소 목적으로합니다. 35를 실행 파지의 DNA 주입을위한 단계 5.5 ° C는 온도에 민감한 cI857 allele의 유도를 방지하기 위해 선택됩니다.
파지 정화 :
jove_content "> 파지 정화 단계 (2.11 단계 2.1) 기타 정화 프로토콜 5 대체하지만, CsCl 평형 기울기 (단계 2.12 및 2.13)을 통해 최종 ultracentrifugation은 피할 수 있습니다. 스윙 버킷 로터는 단계 2.10과 2.12에 필요합니다 순수한 파지 주식을 취득하는 것은 쉽게 주일까지 소요될 수 있습니다. 날카로운 눈에 보이는 파지 밴드를 보장하기 때문에 확실히 아무 중간 단계에서 잘못하지 못하게 길을 따라 파지의 titer를 확인 할 필요가 있습니다.파지 처리 :
제 2의 모든 정화 절차 동안, 그것은 파지는 파지 헤드에서 파지의 꼬리를 전단 않도록 부드럽게 lysate 처리하는 것이 중요합니다. 제 5 (예를 들어, 5.7을 통해 5.5 단계)의 세포 감염 동안, 그것은 또한 감염된 세포에서 파지 입자의 전단을 방지하는 것이 중요합니다. 파지는 그 DNA를 주입 한 후 감염된 세포에서 끊어 경우, 결과는 "어둠"의 감염입니다 참고로 즉fection의 결과는 실험에서 관찰하지만 감염 파지는 안 될 것입니다. phages 또는 파지 / 세포 혼합물을 처리 할 때마다 이러한 문제를 최소화하기 위해, 우리는 다양한 피펫 팁을 사용합니다.
DAPI 테스트 :
DAPI (제 4 항)과 파지 주식을 염색하면 파지 주식의 순도를 확인하기 위해 신속하고 효율적인 방법입니다. 또한 시간이 지남에 따라 기존 파지 주식의 가능한 저하를 테스트하는 데 사용할 수 있습니다. 순수한 주식은 형광 현미경으로 YFP와 DAPI 신호의 공동 지방화 가까운 100 %이어야합니다. 우리는 일반적으로 YFP 명소의 1 % 미만이 입자가 성공적으로 바이러스 DNA를 포장하지 못했음을 나타냅니다하거나 이미 다른 곳에서 자신의 DNA를 주입 한 DAPI을 (바이러스 성 게놈없이 capsids을 대표하는) 포함하지 않는 것이 관찰합니다. DAPI 명소의 1 % 미만이 YFP를 (비 형광 phages에 해당)를 포함하지 않습니다. 이 경우가 아닌 경우, O에서 반복 할 필요가 2.14으로 2.12 단계를다시 정화 rder. 더 장기적으로 라이브 셀 이미징 여기 필요하지 않습니다 때문에 이미지 매개 변수로 돌아가서 단계 4.3 현미경 설정은 섹션 5에서처럼 중요하지 않습니다. 바라던 하나의 파지 입자의 형광 강도를 조정하는 경우 5 항에서와 같은 현미경 설정을 유지하는 데 유용합니다. PBS-아가로 오스 판은 매우 깨끗하고, 또는 DAPI 염색을 사용 너무 많이없는 경우, 파지의 DNA에 해당하는 일부 DAPI 장소는 "후광"로 둘러싸여 될 수 있습니다. 너무 작은 DAPI의 염료를 사용하는 경우, DAPI 채널의 신호는 매우 약한 수 있습니다.
현미경 시스템 :
제 6의 이미지를 들어, 우리는 100x 목표 (계획 Fluo, 수치 공 1.40, 기름 침수 세례) 및 표준 필터 세트 (니콘)와 상업 역 epifluorescence 현미경 (이클립스 TE2000-E, 니콘)을 사용합니다. 형광 광원은 빛 강도 제어 아크 램프입니다. x, y 및 z PO : 다음 기능은 컴퓨터 제어sition, 시야 및 형광 셔터, 그리고 형광 필터 선택하실 수 있습니다. 자동 초점 기능이 필요합니다. 그렇지 않으면, 초점은 쉽게 시간 경과 동영상 (일반적으로 4시간 길이) 동안 멀어지기 수 있습니다. 이 병렬에서 여러 감염 이벤트를 수행 할 수 있습니다 각 시점에서 (X, Y) 여러 위치를 취득 할 수있는 기능은 유용합니다. 우리는 일반적으로 100 감염 이벤트까지 따라 각 영화 8 단계 위치를 취득. 우리가 사용하는 카메라는 16 비트 (Cascade512, Photometrics)의 동적 범위 16x16 규격 μm 픽셀 카메라와 냉각 512x512 CCD입니다. 인수는 MetaMorph 소프트웨어 (분자 장치)를 사용하여 수행됩니다. 현미경은 온도 조절 객실에 배치해야한다 또는 현미경 단계는 온도 조절 챔버로 둘러싸인해야합니다.
이미지 수집 :
라이브 셀 이미징를 들어, 표백하고 검사하고 사진을로 이어질 수 샘플의 불필요한 노출을 방지하기 위해 중요합니다totoxicity. 따라서 먼저 표백 과도하거나 억제 세포의 성장으로 이어지는되지 않고 형광 검출이 가능 최적의 빛 노출을 찾기 위해 시스템을 특징하는 것이 가장 좋습니다. 좋은 형광 이미지를 얻으려면 흥미 진진한 광도, 노출 시간 및 카메라 게인을 재생합니다. 단계 6.2-6.3에서 10 분 프레임 간격은 빛 노출을 최소화하기위한 목적으로 선택됩니다. 각 프레임에서 단지 하나의 포커스에 이미지가 위상 대비 (셀 인식) 및 형광 채널 (세포의 운명을 결정하는)에 필요합니다. 제 1 시점에서 YFP 채널을 통해하지만, 여러 Z-위치 이미지는 세포 표면에있는 모든 감염 phages를 캡처 할 필요가 있습니다. 초기 프레임에 YFP 노출 시간도 나중에 프레임에서 시간 경과 영화에 사용보다 더해야 할 수 있습니다.
이미지 분석 :
매우 신중하게 단계 7.1에서 세포 표면 주위에 파지 입자를 계산합니다. 으로상술 한 바와 같이, 우리는 단계 6.2에서 YFP 채널을 통해 Z-스택 일련의를. 그러나이 아직 계산을 도전 일부 형광 파지 입자 밖에서 초점을 남길 수 있습니다. 초기 시간 프레임에서 셀의 길이는 Metamorph 소프트웨어를 사용하여 측정됩니다. 셀의 길이는 ImageJ 또는 다른 소프트웨어 도구에 의해 측정 할 수 있습니다. 또한, 자동화 된 집 지어 MATLAB 프로그램은 세포 lineages 따라 시간이 지남에 형광 변화 등의 정보를 얻어 매우 유용 할 수 있습니다.
Disclosures
관심 없음 충돌이 선언 없습니다.
Acknowledgments
우리는 마이클 Feiss과 파지 생성 및 정화에 안내를위한 장 Sippy에 감사하고 있습니다. 우리는 세포 인식 소프트웨어, Schnitzcell를 제공하기 위해 마이클 Elowitz 감사드립니다. , 웰치 재단 (기금 Q-1759)과 인간 프론티어 과학 : 골딩 연구실에서 작품은 국립 보건원 (R01GM082837), 국립 과학 재단 (살아있는 세포의 물리 센터 082,265, PFC)의 보조금에 의해 지원됩니다 프로그램 (RGY 2,008분의 70).
Materials
| Name | Company | Catalog Number | Comments |
| Chloroform | Fisher Scientific | C298-500 | |
| NaCl | Fisher Scientific | S271-3 | |
| DNase I | Sigma-Aldrich | D4527-10KU | |
| RNase | Sigma-Aldrich | R4642-10MG | |
| PEG8000 | Fisher Scientific | BP233-1 | |
| SM buffer | TEKnova, Inc. | S0249 | |
| NZYM | TEKnova, Inc. | N2062 | |
| CsCl | Sigma-Aldrich | C3011-250G | |
| Syringe | BD Biosciences | 309585 | |
| Needle | BD Biosciences | 305176 | |
| Dialysis cassette | Thermo Fisher Scientific, Inc. | 66333 | |
| Microscope slide | Corning | 2947-75x50 | |
| Agarose | Fisher Scientific | BP160-100 | |
| SW40Ti ultra-clear tube | Beckman Coulter Inc. | 344060 | |
| SW60Ti ultra-clear tube | Beckman Coulter Inc. | 344062 | |
| SW40Ti rotor | Beckman Coulter Inc. | 331302 | |
| SW60Ti rotor | Beckman Coulter Inc. | 335649 | |
| Refractometer | Fisher Scientific | 13-947 | |
| Epifluorescence microscope | Nikon Instruments | Eclipse TE2000-E | |
| Table 2. Reagents and equipment. | |||
References
- Oppenheim, A. B., Kobiler, O., Stavans, J., Court, D. L., Adhya, S.
- Ptashne, M. A genetic switch : phage lambda revisited. , 3rd edn, Cold Spring Harbor Laboratory Press. (2004).
- Hendrix, R. W. Lambda II. , Cold Spring Harbor Laboratory. (1983).
- Hershey, A. D. The Bacteriophage lambda. , Cold Spring Harbor Laboratory. (1971).
- Zeng, L. Decision making at a subcellular level determines the outcome of bacteriophage infection. Cell. 141, 682-691 (2010).
- Edgar, R. Bacteriophage infection is targeted to cellular poles. Mol. Microbiol. , (2008).
- Ausubel, F. M. Current protocols in molecular biology. , John Wiley & Sons. (1994).
- Sambrook, J., Russell, D. W. Molecular cloning : a laboratory manual. , 3rd edn, Cold Spring Harbor Laboratory Press. (2001).
- Fasman, G. D. Practical handbook of biochemistry and molecular biology. , CRC press. (1989).
- Kaiser, A. D. On the internal structure of bacteriophage lambda. J. Gen. Physiol. 49, 171-178 (1966).
- Kobiler, O. Quantitative kinetic analysis of the bacteriophage lambda genetic network. Proc Natl Acad Sci. 102, 4470-4475 (2005).
- Shaner, N. C. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 22, 1567-1572 (2004).
- Personal communication with M. Feiss. , Forthcoming.
- Schwartz, M. The adsorption of coliphage lambda to its host: effect of variations in the surface density of receptor and in phage-receptor affinity. J. Mol. Biol. 103, 521-536 (1976).

{kind=link}