

Summary
מאמר זה מתאר את התהליך להכנת גרסה מתויגת-fluorescently של bacteriophage מבדה, זיהום של
Abstract
המערכה המהווה bacteriophage (פאג) למבדה וחיידק E. coli שמש זמן רב כדוגמה לנחישות 1,2 תא גורל. בעקבות הזיהום בו הזמני של התא על ידי מספר פאגים, אחד משני מסלולים נבחר: 3,4 מוסס (ארסי) או lysogenic (רדום). לאחרונה פתחנו שיטה לתיוג פאגים בודדים fluorescently, והצליחו לבחון את ההחלטה לאחר פגיעה בזמן אמת מתחת למיקרוסקופ, ברמה של 5 פאגים ותאים בודדים. כאן, אנו מתארים את ההליך המלא לביצוע ניסויי הזיהום המתוארים בעבודתנו הקודמת 5. זה כולל יצירה של פאגים ניאון, זיהום של התאים, הדמיה מתחת למיקרוסקופ וניתוח נתונים. Phage הניאון הוא שיתוף מבטא-גרסות "היברידיות", סוג פרוע וYFP היתוך של חלבון GPD capsid. Lysate phage גולמי הושג לראשונה על ידי גרימת lysogen של GPD-EYFP (Enhanced חלבון) phage פלורסנט הצהוב, חסת סוג הבעת GPD פראי פלסמיד. סדרה של שלבים לטיהור הם בצעו אז, אחרי DAPI תיוג והדמיה מתחת למיקרוסקופ. הדבר נעשה במטרה לאמת את האחידות, יעילות אריזת ה-DNA, אות פלואורסצנטי ויציבות מבנית של מניית פאג. הספיחה הראשונית של פאגים לחיידקים מתבצעת על קרח, ואז אחרי דגירה קצרה ב 35 ° C כדי לעורר הזרקת DNA נגיפית 6. Phage / תערובת חיידקים ואז עבר למשטח של גוש דק תזונתי אגר, מכוסה coverslip וצלם תחת מיקרוסקופ epifluorescence. התהליך שלאחר הזיהום במעקב במשך 4 שעות, במרווח דקות 10. עמדות שלבים מרובות מעקב כזה ש~ 100 זיהומים סלולריים יכולים להיות נעוצים בניסוי אחד. בכל נקודת זמן ומקום, תמונות שנרכשו בשלב הניגוד והערוצים אדומים וירוקים זרחניים. תמונת שלב הניגוד משמשת מאוחר יותר לאוטומטי celהכרת l בעוד ערוצי הניאון משמשים לאפיין את תוצאת הזיהום: ייצור של פאגים ניאון חדשים (ירוק) ואחרי תמוגה תא, או ביטוי של גורמי lysogeny (אדום) ואחריו צמיחה וחלוקת תאים התחדש. את הסרטים שנרכשו זמן לשגות מעובדים באמצעות שילוב של שיטות ידניות ואוטומטיות. תוצאות ניתוח נתונים בזיהוי פרמטרי זיהום לכל אירוע זיהום (מספר דוגמה ועמדות של הדבקת פאגים), כמו גם תוצאת זיהום (תמוגה / lysogeny). פרמטרים נוספים ניתן לחלץ במקרה צורך.
Protocol
1. יצירת lysate phage הגולמי (איור 1)
- בבקבוק מ"ל 50, לחסן מושבה טריה של LE392 (λ LZ1) [pPLate * D] (ראה טבלת מס '1 לפרטים נוספים) ל6 מ"ל של מדיום LB 7 בתוספת עם 10 מיקרוגרם / המ"ל Kanamycin ו100 מיקרוגרם / המ"ל אמפיצילין. לגדול בין הלילה של 30 ° C עם רעד קל (180 סל"ד).
- דלל את 1:100 התרבות לLBM (LB בתוספת 10 mM MgSO 4) וגדל ב 30 מעלות צלזיוס עם רעד קל (180 סל"ד). על מנת לייעל את תשואת phage, לוודא שנפח התרבות הוא לא יותר מעשירית מקיבולת נפח הבקבוק. אנחנו בדרך כלל להכין שתי צלוחיות של קיבולת 2 ליטר או 2.5 ליטר, ולהוסיף תרבות הלילה 2.5 מ"ל למדיום LBM 250 מ"ל בכל בקבוק.
- כאשר צפיפות התאים מגיעה OD 600 ≈ 0.6 (~ 2.5-3 שעות), לגרום lysogen ידי הזזת התרבות ל42 ° C מים באמבטיה שייקר ל18 דקות עם רעד קל (180 סל"ד), ולאחר מכן incubatדואר על 37 מעלות צלזיוס עם רעד קל (180 סל"ד) עד תמוגה גלויה (התרבות הופכת להיות ברורה, ב~ 60-90 דקות).
- הוסף כלורופורם 2% לתרבות, לנער ביד לערבב, ואז דגירה במשך 15 דקות בטמפרטורת חדר. זהירות: ללבוש כפפות כדי לטפל בכלורופורם, ולהימנע מלנשום אותו.
- העברת התרבות לשני בקבוקים 250 מ"ל צנטריפוגות, סרכזת התרבות בSorvall GSA הרוטור ב10000 סל"ד במשך 15 דקות ב 4 ° C. שחזר את supernatant המכיל את חלקיקי phage, והשלך את הגלולה של פסולת. בצע צנטריפוגה שנית, כדי לוודא כדי להיפטר מהפסולת הגלויה.
- השתמש פרוטוקול טיטרציה phage סטנדרטי 8 כדי למדוד את ריכוז פאג. כייל phage צריך להיות 5-10 ~ x 10 9 pfu / מ"ל. השתמש מתח supF כגון LE392 כמחוון המתח בגלל Sam7 המוטציה בגנוטיפ של פאג הניאון, ולהשתמש בחלק עליון ואגרו צלחות אגרו עשה עם NZYM העשיר להשיג הפלאק גדול יותר (Figure 2).
2. טיהור phage (איור 1)
- יוצקים לתוך lysate בקבוק גדול (לדוגמה: 2-ליטר), להוסיף DNase אני וRNase (מיקרוגרם 1 / מ"ל כל אחד) לlysate כדי לעכל את החומצות המשוחררות מחיידקי lysed הגרעין, ודגירת 1 שעה בטמפרטורת חדר.
- הוסף NaCl 1M לlysate, להעביר lysate לבקבוקי 250 מ"ל צנטריפוגות, ודגירה 3 שעות על קרח. צנטריפוגה lysate בSorvall GSA ב 10,000 סל"ד במשך 15 דקות ב 4 ° C. שחזר את supernatant. כייל phage צריך להיות דומה לזה של lysate הגולמי, שהוא ~ 5-10 x 10 9 pfu / מ"ל. התוספת של NaCl מקדמת ניתוק של חלקיקי phage מפסולת חיידקים, ודורש למשקעים יעילים של חלקיקי phage ידי PEG 8.
- יוצק לתוך lysate בקבוק גדול, למשל בקבוק 2-ליטר, להוסיף 10% (w / v) PEG8000 לlysate, מערבבים לאט או לנער לפזר PEG8000 בטמפרטורת חדר. העברת lysate לסנאט 250 מ"לבקבוקי rifuge ואז דגירת הלילה (~ 16 שעות) ב 4 ° C. צנטריפוגה lysate בSorvall GSA הרוטור ב 10,000 סל"ד במשך 15 דקות ב 4 ° C. השלך את supernatant.
- משרה את הגלולה (חלקיקי phage זרזו עם PEG8000) עם phage SM חיץ (4 חיץ SM מ"ל ל250 מ"ל של lysate phage הראשוני). דגירה עם רעד קל או מאוד לא רועד עבור 16 שעות ב 4 ° C.
- מוציא בעדינות lysate (חיץ SM עם חלקיקי phage) לתוך צינור צנטריפוגה 50 מ"ל אפנדורף, ולאחר מכן לשטוף את הגלולה שנותרה עם 0.5 - 1 מ"ל של חיץ SM.
- הוסף נפח שווה של כלורופורם לlysate. בעדינות מערבב את lysate עם כלורופורם על ידי עד היפוך ושוב כמה פעמים. צנטריפוגה ב 4000 סל"ד במשך 15 דקות ב 4 ° C באפנדורף 5804R או צנטריפוגה דומה עליונת ספסל.
- חזור על שלב 2.6 כדי לקבל lysate ברור יותר. כייל phage צריך להיות 1-2 ~ x 10 11 pfu / מ"ל.
- הכן את פתרוני SM / CsCl עם שלוש צפיפויות שונות (ρ) של 1.3 גר '/ מ"ל, 1.5 גרם / מ"ל ו 1.7 גר '/ מ"ל. למדוד את מקדם השבירה (η) כדי לקבל קריאה מדויקת יותר צפיפות. המרת הצפיפות 9 היא ρ = 10.8601 η - 13.4974 ב 25 ° C. ראה טבלה 3 לקבלת פרטים.
- השתמש במזרק עם מחט ארוכה כדי לטעון את הפתרון לתוך צינור 14 מיליליטר אולטרה ברור קמן 40Ti ultracentrifuge. כדי להימנע מערבוב כדי ליצור שיפוע צפיפות טובה יותר, underlaying הפתרון (פתרונות כלומר שכבות של צפיפות גוברת תחת אחד אחר) צריך להיות בשימוש, כלומר, בעדינות לטעון 2 מ"ל של פתרוני SM / CsCl בצו של 1.3 גר '/ מ"ל, 1.5 גרם / מ"ל ו -1.7 ג'/ מ"ל על ידי החדרת המחט עם מזרק 3 מ"ל לתחתית של התחתית.
- עדינות לטעון 8 מ"ל של lysate phage ידי כיסה מהחלק העליון של צינור ultracentrifuge 14 מ"ל. הכן את צינור איזון. צנטריפוגה בקמן SW40Ti הרוטור ב24000 סל"ד במשך 4 שעות ב 4 ° C.
- מוציא בעדינות את הצינור בחדר חשוך ולהאיר מהחלק העליון של הצינור על רקע שחור באמצעות aflashlight. להקת phage צריכה להיות נראית בבירור במיקומו של הממשק בין 1.3 גר '/ מ"ל ו -1.5 שכבות גרמו / מיליליטר SM / CsCl (איור 3 א). לנקב בצד של הצינור מעט מתחת ללהקה עם מחט 21.5 מד עם מזרק 3 מ"ל. עדינות לאסוף ~ 500 μl של השעית פאג. כייל phage צריך להיות 5-10 ~ x 10 11 pfu / מ"ל.
- הנח את השעית phage לתוך צינור 4 מיליליטר אולטרה ברור קמן ultracentrifuge SW60Ti הרוטור. מלא את הצינור עם 1.5 גר '/ מיליליטר SM / פתרון CsCl. הכן את צינור איזון. צנטריפוגה בקמן SW60Ti הרוטור ב35000 סל"ד במשך 24 שעות ב 4 ° C.
- חזור על אותו התהליך כמו בשלב 2.11 לאסוף phage מהלהקה הגלויה. הלהקה צריכה להיות גלויה כפי שמוצג באיור 3 ב.
- טען השעית phage לתוך קלטת קרום דיאליזה (טבלה 2) ודיאליזה שלוש פעמים נגד נפח 1000-קיפול של חיץ SM ב 4 מעלות צלזיוס במשך תקופות של 3 שעות, 3 שעות וovernigHT (~ 16 שעות). מטרת הדיאליזה היא להיפטר מהווה CsCl בהשעית פאג. כייל phage הסופי צריך להיות ~ 5-10 x 10 11 pfu / מ"ל.
3. הכן לוח 1 agarose ג'ל (איור 4)
- נקה 6 שקופיות מיקרוסקופ (75 מ"מ x 50, עובי 1 מ"מ) עם אתנול 70%.
- סדר 5 מגלשות ולאבטח עם קלטת כפי שמוצג באיור 4.
- מערבב 0.09 גרם agarose לתוך מדיום 6 מ"ל בכוס קטנה מכוסית בפלסטיק נצמד (מניב 1.5% agarose). מחמם על כיריים עד שהתמיסה הופכת ברורה.
- יוצק את פתרון agarose על גבי השקופיות המאובטחות.
- הנח את השקופית האחרונה על העליונה, הימנעות בועות אוויר בזהירות. מקום על גבי משקל ולאפשר להתקרר במשך ~ 30 דקות.
- הסר את 4 שקופיות בצד, ולעטוף את הגוש יחד עם שקופיות העליונות ותחתונות עם פלסטיק נצמד. הלוח ניתן לאחסן ב 4 מעלות צלזיוס למשך עד 3 ימים.
4. בדיקת מלאי phage המטוהר
- להכיןלוח PBS-agarose ג'ל כפי שתואר לעיל (סעיף 3).
- כתם phage המטוהר עם DAPI. מערבב 10 μl של פאג (~ 1 x 10 10 pfu / מ"ל) עם 10 μl של 10 מיקרוגרם / המ"ל DAPI (ריכוז DAPI סופי של 5 מיקרוגרם / מ"ל), דגירה במשך 30 דקות ב 4 ° C או 10 דקות בטמפרטורת חדר.
- μl מקום 1 של תערובת phage / DAPI במרכז 24 מס '1 x 50 המ"מימ coverslip, כיסוי חתיכה קטנה (~ 10 x 10 מ"מ) של הלוח המוכן מראש PBS-agarose. הפיסה הקטנה של לוח agarose היא לחתוך בסכין גילוח לאחר השקופית העליונה בג'ל הכריך מוסר. תמונת המדגם תחת מיקרוסקופ epifluorescence באמצעות ערוצי YFP וDAPI. פאגים בודדים צריכים להיות גלויים כמו "כתמים" עקיף מוגבלי ניאון בשני הערוצים (איור 5). השתמש באותו מיקרוסקופ וsetups מצלמה כמו בשלב 6.2 להלן.
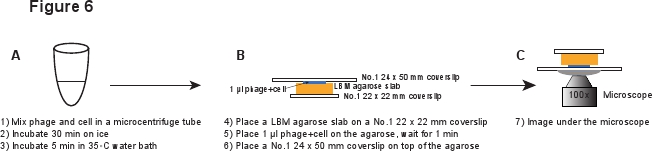
5. זיהום (איור 6)
- בצינור 14 מ"ל פלקון, לחסן מושבה טריה של LE392 [PP RE-MCHerry] (ראה טבלת מס '1 לפרטים נוספים) לתוך 2 מ"ל של מדיום LB בתוספת 100 מיקרוגרם / המ"ל אמפיצילין, 10 המ"מ 4 MgSO 0.2% ומלטוז. לגדול בין הלילה על 37 מעלות צלזיוס עם רעד בגוף בינוני (265 סל"ד).
- דלל 1:1000 התרבות לLBMM (LB בתוספת 10 mM MgSO 4 ו 0.2% מלטוז), כלומר להוסיף תרבות הלילה 5 μl למדיום LBMM 5 מ"ל בבקבוק ml 50. לגדול ל OD 600 ≈ 0.4 על 37 מעלות צלזיוס עם רעד בגוף בינוני (265 סל"ד).
- השתמש בינוני LBM להכין לוח ג'ל LBM-agarose כמפורט בסעיף 3 לעיל.
- צנטריפוגה 1 מ"ל של תאים ב2000 גרמו בmicrocentrifuge עליון ספסל עבור 2 דקות בטמפרטורת חדר. הסירו את supernatant, ועדינות resuspend התאים ל20 LBMM μl קר כקרח כדי להגיע OD 600 & 20.
- מתי לטפל בסחורת phage המטוהרת, להשתמש קצה פיפטה רחב או לחתוך את קצה פיפטה רגיל לעשות את קצה פתיחה רחב יותר, כדי למנוע הטית חלקיקי phage 3. עדינות Mix 20 μl של תאים עם 20 μl מטוהר phage להגיע יחס פאגים לתא ממוצע בטווח של 0.1 - 5. לדגור על קרח למשך 30 דקות כדי לאפשר phage ספיחה, ולאחר מכן מדגיר ° C אמבט מים 35 למשך 5 דקות כדי לעורר הזרקת דנ"א phage 6.
- פיפטה מעלה ומטה מספר פעמים להפריד כל אגרגטים סלולריים. שוב להשתמש קצה פיפטה רחבה כדי למנוע את גזירת פאגים. דלל את התערובת לתוך 1:10 LBMM, למשל תערובת μl 5 לLBMM μl 45.
- מניח חתיכת לוח LBM-agarose (~ 10 x 10 מ"מ) על coverslip מס '1 22 x 22 מ"מ. הלוח המוכן מראש LBM-agarose צריך להיות ממוקם בטמפרטורת חדר למשך שעה לפחות 1 לפני שימוש על מנת להבטיח כי לוח agarose מגיע לטמפרטורת חדר. μl המקום 1 של תערובת phage / תא בלוח agarose ולחכות לדקות 1 כדי לאפשר לתערובת לספוג לתוך לוח agarose. מניח בעדינות 24 מס '1 x 50 מ"מ coverslip בחלק העליון של לוח agarose. הליך זה נועד למנוע הטיה על פאגים מinfתא ected (איור 6).
6. בעקבות גורל תא מתחת למיקרוסקופ
- זהירות הר coverslip על הבמה של מיקרוסקופ. עבור הדמיה, השתמש-הגדלה גבוהה (למשל 100x) אובייקטיבית (ראה מערכת מיקרוסקופ בדיון להלן).
- לרכוש תמונה מוגדרת למסגרת הזמן הראשונית. סט תמונה זו משמשת כדי לאפיין את המספרים ועמדות הראשוניים של כל פאגים המדביק. קח סדרה של 15 תמונות במרווחי 200 ננומטר Z-ציר (אנכי). תמונה דרך ערוץ YFP. בנוסף, לקחת תמונה אחת בפוקוס בשלב הניגוד וערוצי mCherry. לייעל את עוצמת האור ומשך חשיפה לקבלת אות חזקה מספיק תוך מזעור הלבנה וניזק לתאים (ראה יבוא תמונות בדיון להלן).
- לרכוש סרט זמן לשגות של גורל התא לאחר פגיעה. תמונת המדגם בשלב לעומת זאת, YFP וmCherryערוצים במרווחי זמן של 10 דקות לשעות סביב 4. במהלך סרט זמן לשגות, השתמש תמונה אחת Z-עמדה לכל ערוץ לכל נקודת זמן, כדי להימנע מחשיפה מיותרת של המדגם, אשר עלול להוביל להלבנה וphototoxicity.
7. ניתוח תמונה
- ידני לספור את מספר פאגים ומיקום השיא phage ואורך התא במסגרת הזמן הראשונית. ניתן לעשות זאת באמצעות תוכנה כמו MetaMorph או ImageJ. הקלט את גורל התא (ממוסס, lysogenic או נגוע), זמן תמוגה, וכל מידע מבוקש אחר על ידי משחק בסרט זמן לשגות. כדי לזהות גורל תאים שונים, לראות סרט זמן לשגות בסעיף תוצאות נציג בהמשך.
- בנוסף לניתוח הידני לעיל, מידע כמוני יותר (למשל רמת פלואורסצנטי לאורך זמן בתאים בודדים) ניתן לחלץ באמצעות אלגוריתמי תא לזיהוי ומעקב אוטומטיים שושלת. אנו משתמשים בתכנת Matlab בית שנבנה עבור tracing המשפחה של תאים ורמות קרינה, יחד עם קוד Schnitzcell Matlab לפילוח תא (שנכתב על ידי קבוצת Elowitz בקלטק).
8. תוצאות נציג:
ציפוי phage:
את הפלאק של פאגים כותרתו fluorescently (ב -1.6 צעד וסעיף 2) הוא קטן משמעותי מאלה של זן בר (איור 2). לפיכך, אנו מדגירים את הצלחות לפחות 12 שעות ב37 מעלות צלזיוס החממה לפלאק תהיה גלויה.
Ultracentrifugation phage:
לאחר ultracentrifugation של מדגם phage עם שיפוע צעד CsCl (שלב 2.10), שתי להקות צריכות להיות גלויות (איור 3 א). הלהקה העליונה, בממשק שבין השעית phage ושכבת SM / CsCl 1.3 גרם / מ"ל, מכילה פסולת תא וcapsids phage הריק. הלהקה התחתונה, בממשק שבין SM / CsCl 1.3 גרם / מ"ל ו -1.5 שכבות גרמו / מ"ל, היא להקת פאג. תיהלהקה של ירקרקה מופיעה לphage λ LZ2 הניאון. הלהקה הפרועה הסוג פאג λ IG2903 מופיעה כחלחלה 5. לאחר ultracentrifugation של שיפוע שיווי המשקל CsCl בשלב 2.12, להקת phage אחד צריכה להיות גלויה בחלק האמצעי של הצינור (איור 3 ב '). מאז phage λ ניאון LZ2 מכיל תערובת של GPD-EYFP וGPD capsids, היחס של חלבון לדנ"א הוא גבוה מזה של זן בר. לכן, הלהקה של ניאון phage λ LZ2 היא מעט קלה יותר (נראה כי במיקום גבוה יותר בצינור) מזו של סוג הפראי λ IG290310.
כתמי DAPI:
איור 5 מראה תמונות אופייניות המתקבלות לאחר תיוג phage עם DAPI (סעיף 4). אותות YFP וDAPI של פאג מטוהר בהצלחה צריכים קרוב ל 100% התכתבות. אנחנו בדרך כלל לצפות כי פחות מ 1% מנקודת YFPזה אינו מכיל DAPI (המייצג capsids ללא הגנום הנגיפי). פחות מ 1% מנקודתי DAPI אינם מכילים YFP (מתאים לפאגים שאינם ניאון) 5.
סרט זמן לשגות:
תאי ממוסס מוכרים על ידי ההצטברות של YFP פלואורסצנטי (ערוץ ירוק) בתוך התא, ואחרי תמוגה תא. תאי Lysogenic מוכרים על ידי ההצטברות של מדי mCherry פלואורסצנטי (אדום) בתוך התא וחידוש צמיחת תאים נורמלים וחילוק. תאים נגועים (או תאים שבו הזיהום נכשל) לא יציגו כל אחד מהפנוטיפים הנ"ל וגדל ומתחלקים באופן נורמלי. איור 7 מראה כמה סטים-תמונה של שלב לעומת זאת, ערוצי mCherry, ואת התמונות המעולפות המקבילות של אלה וYFP שלושה ערוצים, מסרט הזמן לשגות טיפוסי (סעיף 6). את פאגים הבודדים (נקודות ירוקות) הם נראים בבירור במסגרת הזמן הראשוני (סעיף 7 איור). בדרך כלל, מספרשל פאגים נראים על פני התא (כנראה להדביק התאים האלה) ואילו פאגים אחרים unadsorbed, כפי שמוצגים באיור 7 ב (פנל שמאלי). תוצאת הזיהום הופכת להבחין לאורך זמן. מחזור ממוסס מצוין על ידי ייצור התאי של פאגים חדשים (ירוק, איור 7 ג) ואחרי תמוגה תא (תאים שהתפוצץ עם פאגים ירוקים משוחררים, האיור 7D). Lysogeny מצוין על ידי הייצור של mCherry מאמרגן P רי (אדום, איור 7 ג) ואת חידוש צמיחת תאים וחטיבה (אדום, האיור 7D).




איור 1. א.ב.). Phage מטוהר באמצעות סדרה של צעדים (CL פנלים).

איור 2. הפלאק phage. לוחות של פאג הניאון (משמאל) קטנים מאלה של זן בר (מימין) לאחר דגירת צלחות עבור 12 שעות ב 37 ° C.

איור 3. להקות phage לאחר ultracentrifugation. א) שתי להקות גלויים לאחר ultracentrifugation בשיפוע צעד CsCl. 1 העליון מתאים לפסולת תא וcapsids phage הריק; הלהקה התחתונה מכילה phage הרצוי. משמאל: phage ניאון, ימין:. סוג B פראי ) להקת phage אחת גלויה לאחר ultracentrifugation בשיפוע שיווי משקל CsCl. להקת phage הניאון (משמאל) היא ירקרקה, בהשוואה ללהקה כחלחלה לphage סוג הפרוע (מימין).

איור 4. ההליך של הכנת לוחות ג'ל agarose.

איור 5. תמונות של פלורסנט פאגים לאחר צביעת DAPI. פאגים בודדים הם להבחין בקלות, וYFP וDAPI מאותת שיתוף לוקליזציה טובה מאוד.

איור 6. תיאור סכמטי של זיהום phage והתקנת הדמיה. לחץ כאן לצפייה בגרסה בגודל מלא של תמונה זו.
gure 7 "src =" / files/ftp_upload/3363/3363fig7.jpg "/>
איור 7. תמונות אופייניות מסרט הזמן לשגות של זיהום פאג. הראה הם השלב לעומת זאת, ערוצי mCherry, כמו גם כיסוי של השלושה הערוצים YFP ו. (A) תמונות YFP ערוצים ממסגרת הזמן הראשונית. שמאל, בסכום של תמונות YFP ב- z עמדות שונות. השלוש התמונות הנכונות הן תמונות YFP מדגם ב-z עמדות שונות, המתאימים לאזורים שונים על פני התא. (ב), (ג) ו (ד ') תמונות מעולף (משמאל) של שלב הניגוד (באמצע שמאל), ערוצי YFP (אמצע ימין) וmCherry (מימין) בטווחי זמן שונים. (ב) בt = 0, שני תאים נראים, כל אחד הנגוע בphage בודד (נקודות ירוקות), ותא אחד נגוע על ידי 3 פאגים. נצפו גם כמה פאגים unadsorbed. (ג) בt = 80 דקות, שני התאים שנדבקו ב פאגים בודדים, כל אחד נכנסו למסלול ממוסס, כפי שמעידד על ידי הייצור התאי של פאגים חדשים (ירוק). התא הנגוע ב3 פאגים הושקע מסלול lysogenic, כפי שצוין על ידי הייצור של mCherry מאמרגן PRE (אדום). (ד) בt = 2 שעות, מסלול ממוסס הביא תמוגה תא (תא התפוצץ), ואילו תא lysogenic חלק §.
§ פנלים השמאליים של איור 7 (ג) (ד) הם שהודפס מתא, 141, Lanying זנג, סמואל א 'סקינר, Chenghang זונג, ז'אן עם קש, מיכאל Feiss, ועידו גולדינג, קבלת החלטות ברמת subcellular מקובע את התוצאה של זיהום bacteriophage, 682-691, זכויות יוצרים (2010), באישור Elsevier.
| מסנן שם | גנוטיפ הרלוונטי | מקור / התייחסות |
| זני חיידקים | ||
| LE392 | supF | ג'ון Cronan, אוניברסיטת אילינוי |
| זני phage | ||
| λ LZ1 | GPD-EYFP, D-eyfp cI857 Sam7 ב :: kanR | זנג et al. 5 |
| λ LZ2 | GPD-פסיפס, אותו גנוטיפ כλ LZ1 | זנג et al. 5 |
| פלסמידים | ||
| PP רי - mCherry | mCherry תחת השליטה של P רי, המגבר R | זנג et al. 5 |
| pPLate * D | GPD תחת השליטה של אמרגן מאוחר λ, המגבר R | זנג et al. 5 |
טבלת 1. זני חיידקים,פאגים ופלסמידים משמשים בעבודה זו.
| ρ צפיפות (גרם / מ"ל) | CsCl (ז) | SM (מ"ל) | השביר η |
| 1.30 | 39 | 86 | 1.3625 |
| 1.50 | 67 | 82 | 1.3815 |
| 1.70 | 95 | 75 | 1.3990 |
לוח 3. פתרוני CsCl ערוך SM חיץ (100 מ"ל) לשיפועי צעד.
Discussion
זני חיידקים, וphage פלסמידים:
מתח LE392 הוא supF. הוא נבחר כדי לדכא Sam7 המוטציה בגנום phage (ראה טבלת מס '1 לפרטים נוספים). לפיכך, lysogens המושרה סופו של דבר lyse ושחרור חלקיקי phage, כפי שיהיה גם תאים נגועים שבחרו מסלול מוסס. התאים גדלים Lysogenic של 30 ° C בשל הנוכחות של הטמפרטורה הרגישה CI 857 אלל בגנום פאג. לאחר אינדוקצית חום, GPD-EYFP והפראי הסוג GPD הם שיתוף המתבטאים מהגנום של λ LZ1 וpPlate פלסמיד * D בהתאמה. כתוצאה מכך, של capsid החדש שנוצר phage λ LZ2 מכיל תערובת של חלבוני GPD GPD-EYFP ו. phage פסיפס זה הוא יציב מבחינה מבנית ומספיק ניאון כדי לאפשר זיהוי של פאגים בודדים 5. PP רי - mCherry הוא כתב פלסמיד המשמש לאיתור בחירת lysogenic pathway. RE P האמרגן מופעל על ידי כת"ש בהקמת lysogeny 1,11. PP רי - mCherry 5 נגזר מPE-GFP 11 על ידי החלפת GFP עם 12 mCherry. לפרטים נוספים ראה את העבודה שלנו לפני 5.
פרמטרי מצב צמיחה:
במהלך אינדוקצית lysogen (סעיף 1), רעידות קלות ב 180 סל"ד נותנות תשואת וירוס טובה 13. השימוש בגלוקוז במדיום הגידול יש להימנע ככל שחילוף חומרים של הגלוקוז מייצר מוצרים מטבוליות חומציים, וחלקיקי מבדה בוגרים אינם יציבים ב pH חומצי 13. התוספת של 4 MgSO נועדה לייצב phage 3 capsid. לפאגים נושאים סוג CI הפרוע (במקום CI 857), lysogen יכול להיגרם באמצעות mitomycin הסוכן ה-DNA ניזק 3 ג'. בשלב 1.3, הדגירה על 37 המעלות צלזיוס צריכה בדרך כלל לא תעלה על 90 דקות. זה usef אול כדי לבדוק את צפיפות התא על ידי OD 600 בכל 30 דקות. לlysate טוב, OD 600 יורד לכ 0.2 או פחות, ונותר 600 OD הוא תוצאה של פסולת תא. דגירה ארוכה מדי עלולה לגרום לתשואה נמוכה מאז phage phage החדש שנוצר יכול להתחיל להזריק DNA שלהם לתוך תא פסולת. כדי להשיג להקת phage גלויה (לפחות 1 10 11 חלקיקי phage x) בשלבים ו2.11 2.13, לגדול תרבות מ"ל לפחות 500 בשלב 1.2. התוספת של 0.2% למלטוז מדיום גידול בצעדי 5.1 ו 5.2 מיועדת לזירוז ביטוי של כבש, הקולטן לפאג מבדה ספיחת 3,14. דילול 1000-לקפל במקום 100-לקפל בשלב 5.2 נועד להקטין את רמת רקע mCherry מRE הכתב פלסמיד PP - mCherry. בשלב 5.5 להזרקת דנ"א phage מפעיל, 35 ° C נבחר כדי להימנע מגיוס של אלל cI857 הרגיש לטמפרטורה.
טיהור phage:
jove_content "> שלבי טיהור phage (שלבים 2.1 עד 2.11) יכולים להיות מוחלפים עם פרוטוקולי טיהור אחרות 5, אבל ultracentrifugation הסופי דרך שיפוע שיווי המשקל CsCl (צעדים ו2.12 2.13) הוא בלתי נמנע. הרוטורים דלי Swinging יש צורך בצעדי 2.10 2.12 ועד להבטיח להקות phage גלויות חדות. קבלת מניות phage טהורות יכולה בקלות לקחת עד שבוע, ולכן יש צורך לבדוק את כייל phage לאורך הדרך כדי לוודא ששום דבר משתבש במהלך שלבי ביניים.טיפול phage:
במהלך כל הליכי הטיהור בסעיף 2, זה קריטי לטפל phage lysate עדינות, כדי למנוע הטית זנבות phage מראשי phage. במהלך הדבקת תא בסעיף 5 (למשל, צעדי 5.5 דרך 5.7), הוא גם קריטי כדי למנוע ההטיה של חלקיקי phage מהתא הנגוע. שים לב שאם phage הוא טעון מהתא הנגוע לאחר הזרקת ה-DNA שלו, התוצאה היא זיהום "אפל", כלומר בתוצאת fection תהיה נצפתה בניסוי, אבל phage מדביק לא. כדי למזער את בעיות כאלה, אנו משתמשים קצה פיפטה רחבה בכל פעם שטיפול פאגים או תערובת phage / תא.
DAPI בדיקה:
מכתים את מניות phage עם DAPI (סעיף 4) היא שיטה מהירה ויעילה כדי לבדוק את הטוהר של מניית פאג. זה גם יכול לשמש כדי לבדוק לפירוק אפשרי של מניות phage קיימות לאורך זמן. למניות טהורות, שיתוף הלוקליזציה של אותות YFP וDAPI תחת מיקרוסקופ פלואורסצנטי צריכה להיות קרוב ל 100%. אנחנו בדרך כלל לצפות כי פחות מ 1% מנקודתי YFP אינם מכילים DAPI (המייצג capsids ללא הגנום הנגיפי), המצביע על כך שהחלקיקים אלה לא לארוז את ה-DNA הנגיפית בהצלחה או כבר הזריק DNA שלהם במקום אחר. פחות מ 1% מנקודתי DAPI אינם מכילים YFP (מתאים לפאגים שאינם ניאון). אם זה לא המקרה, צעדי 2.12 דרך 2.14 צורך לחזור בorder לטהר שוב. בכל קשורים לפרמטרי הדמיה, התקנת מיקרוסקופ בשלב 4.3 היא לא קריטית כמו בסעיף 5, כי אין הדמיה ארוכת טווח חי תאים נדרשת כאן. עם זאת, שמירה על הגדרות מיקרוסקופיה כמו בסעיף 5 היא שימושית אם רוצה לכייל את עוצמת הקרינה של חלקיק phage אחת. אם לוח PBS-agarose הוא לא מאוד נקי, או יותר מדי DAPI צבע משמש, כמה נקודות מתאימות ל-DNA phage DAPI עשויות להיות מוקפות ב" הילה ". אם צבע DAPI מעט מדי נעשה שימוש, את האות מערוץ DAPI עשויה להיות חלשה מאוד.
מערכת מיקרוסקופ:
להדמיה בסעיף 6, נסתכל במיקרוסקופ epifluorescence הפוך מסחרי (Eclipse TE2000-E, ניקון) עם מטרת 100x (התכנית Fluo, צמצם מספרי 1.40, טבילת שמן) וקובע מסנן רגיל (ניקון). מקור האור פלואורסצנטי הוא מנורת קשת עם שליטה בעוצמת אור. התכונות הבאות הן מבוקר מחשב: X, Y ו Z position; שדה בהיר ותריסי קרינה; ובחירת מסנן קרינה. תכונת פוקוס אוטומטי נדרשה. אחרת, ייתכן שההתמקדות בקלות להיסחף במהלך סרט זמן לשגות (בדרך כלל 4 שעות ארוכות). היכולת לרכוש מרובות (x, y) עמדות בכל נקודת זמן היא שימושית, שכן היא מאפשרת לעקוב אחר אירועי זיהום מרובים במקביל. אנחנו בדרך כלל לרכוש 8 עמדות במה בכל סרט, בעקבות אירועים עד 100 זיהום. המצלמה אנו משתמשים היא מקורר 512x512 CCD עם פיקסל מצלמת מיקרומטר 16x16 עם טווח דינמי של 16 סיביים (Cascade512, Photometrics). רכישה מתבצעת באמצעות MetaMorph תוכנה (התקנים מולקולריים). מיקרוסקופ צריך להיות ממוקם בחדר עם הטמפרטורה מבוקרת, או לחלופין, את שלב המיקרוסקופ צריך להיות מוקף בחדר עם בקרת הטמפרטורה.
יבוא תמונות:
עבור הדמית חית תאים, זה הוא קריטי כדי למנוע חשיפה מיותרת של המדגם, אשר עלול להוביל להלבנה ופtotoxicity. לכן, הדרך טובה ביותר לאפיין את המערכת שלך כדי למצוא את חשיפת אור אופטימלית המאפשרת זיהוי פלואורסצנטי ואילו לא מוביל לצמיחת תאי הלבנה מוגזמת או עיכוב. כדי לקבל תמונה טובה פלואורסצנטי, לשחק עם עוצמת האור המרגש, זמן חשיפה ורווח מצלמה. בצעדים 6.2-6.3, מרווח המסגרת 10 דקות נבחר למטרה למזער חשיפה לאור. בכל מסגרת, רק תמונה אחת בפוקוס יש צורך בשלב ניגוד (להכרת תא) וערוצי ניאון (לקביעת גורל תא). בנקודת הזמן הראשונה, לעומת זאת, תמונות מרובות Z-עמדה דרך ערוץ YFP נדרשות כדי ללכוד את כל פאגים מדביקים על פני התא. זמן חשיפת YFP במסגרת הראשונית עלול גם צריך להיות גבוה יותר מזו המשמשת לסרט זמן לשגות במסגרות הזמן מאוחר יותר.
ניתוח תמונה:
ספירת חלקיקי phage בזהירות רבה מסביב לתא השטח בשלב 7.1. כמואמור לעיל, לנקוט בשורה של Z-ערימות דרך ערוץ YFP בשלב 6.2. עם זאת, זה עדיין עשוי להשאיר כמה חלקיקי ניאון phage מחוץ למיקוד, המאתגר את הספירה. אורך התא במסגרת הזמן הראשונית נמדד באמצעות תוכנת Metamorph. אורך התא גם יכול להימדד ImageJ או כלי תוכנה אחרות. בנוסף, תכנית נבנתה Matlab אוטומטית בית יכולה להיות מאוד שימושית בקבלת מידע כגון שינוי פלואורסצנטי לאורך הזמן לאורך שושלות תאים.
Disclosures
אין ניגודי האינטרסים הכריזו.
Acknowledgments
אנו מודים למיכאל Feiss וז'אן עם הקש להדרכה על יצירת phage וטיהור. אנו מודים למיכאל Elowitz לאספקת תוכנה לזיהוי התא, Schnitzcell. עבודה במעבדת גולדינג נתמכת על ידי מענקים מהמכון הלאומי לבריאות (R01GM082837), הקרן הלאומי למדע (082265, PFC: מרכז לפיסיקה של תאי חיים), קרן וולש (גרנט Q-1759) ומדע Frontier אנוש התכנית (RGY 70/2008).
Materials
| Name | Company | Catalog Number | Comments |
| Chloroform | Fisher Scientific | C298-500 | |
| NaCl | Fisher Scientific | S271-3 | |
| DNase I | Sigma-Aldrich | D4527-10KU | |
| RNase | Sigma-Aldrich | R4642-10MG | |
| PEG8000 | Fisher Scientific | BP233-1 | |
| SM buffer | TEKnova, Inc. | S0249 | |
| NZYM | TEKnova, Inc. | N2062 | |
| CsCl | Sigma-Aldrich | C3011-250G | |
| Syringe | BD Biosciences | 309585 | |
| Needle | BD Biosciences | 305176 | |
| Dialysis cassette | Thermo Fisher Scientific, Inc. | 66333 | |
| Microscope slide | Corning | 2947-75x50 | |
| Agarose | Fisher Scientific | BP160-100 | |
| SW40Ti ultra-clear tube | Beckman Coulter Inc. | 344060 | |
| SW60Ti ultra-clear tube | Beckman Coulter Inc. | 344062 | |
| SW40Ti rotor | Beckman Coulter Inc. | 331302 | |
| SW60Ti rotor | Beckman Coulter Inc. | 335649 | |
| Refractometer | Fisher Scientific | 13-947 | |
| Epifluorescence microscope | Nikon Instruments | Eclipse TE2000-E | |
| Table 2. Reagents and equipment. | |||
References
- Oppenheim, A. B., Kobiler, O., Stavans, J., Court, D. L., Adhya, S.
- Ptashne, M. A genetic switch : phage lambda revisited. , 3rd edn, Cold Spring Harbor Laboratory Press. (2004).
- Hendrix, R. W. Lambda II. , Cold Spring Harbor Laboratory. (1983).
- Hershey, A. D. The Bacteriophage lambda. , Cold Spring Harbor Laboratory. (1971).
- Zeng, L. Decision making at a subcellular level determines the outcome of bacteriophage infection. Cell. 141, 682-691 (2010).
- Edgar, R. Bacteriophage infection is targeted to cellular poles. Mol. Microbiol. , (2008).
- Ausubel, F. M. Current protocols in molecular biology. , John Wiley & Sons. (1994).
- Sambrook, J., Russell, D. W. Molecular cloning : a laboratory manual. , 3rd edn, Cold Spring Harbor Laboratory Press. (2001).
- Fasman, G. D. Practical handbook of biochemistry and molecular biology. , CRC press. (1989).
- Kaiser, A. D. On the internal structure of bacteriophage lambda. J. Gen. Physiol. 49, 171-178 (1966).
- Kobiler, O. Quantitative kinetic analysis of the bacteriophage lambda genetic network. Proc Natl Acad Sci. 102, 4470-4475 (2005).
- Shaner, N. C. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 22, 1567-1572 (2004).
- Personal communication with M. Feiss. , Forthcoming.
- Schwartz, M. The adsorption of coliphage lambda to its host: effect of variations in the surface density of receptor and in phage-receptor affinity. J. Mol. Biol. 103, 521-536 (1976).

{kind=link}