

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Burada açıklanan, farklı makro besin kalitesinde diyetlere maruz kalan Drosophila melanogaster larvaları tarafından belirli bir zaman aralığında yenen yiyecek miktarının kolorimetrik olarak ölçülmesini sağlayan bir protokoldür. Bu tahliller nöronal termogenetik ekran bağlamında gerçekleştirilir.
Abstract
Yemleme ve beslenme davranışları, hayvanların gelişimleri, sağlıkları ve zindelikleri için gerekli olan enerji ve besin kaynaklarına erişmelerini sağlar. Bu davranışların nöronal regülasyonunun araştırılması, beslenme homeostazının altında kalan fizyolojik ve moleküler mekanizmaların anlaşılması için gereklidir. Solucanlar, sinekler ve balıklar gibi genetik olarak çekilebilir hayvan modellerinin kullanılması bu tür çalışmaları büyük ölçüde kolaylaştırır. Son on yılda, meyve sineği Drosophila melanogaster, beslenme ve yemleme davranışlarının nöronal kontrolünü araştıran nörobiyologlar tarafından güçlü bir hayvan modeli olarak kullanılmıştır. Şüphesiz değerli olsa da, çoğu çalışma yetişkin sinekleri inceler. Burada, larvalar protein ve karbonhidrat içeriğinde farklı diyetlere maruz kaldıklarında beslenme davranışlarını kontrol eden nöronal substratları araştırmak için daha basit larva sinir sisteminden yararlanan bir protokol açıklıyoruz. Yöntemlerimiz, nöronal termogenetik aktivasyon ekranı bağlamında gerçekleştirilen nicel kolorimetrik seçimsiz besleme testine dayanmaktadır. Bir okuma olarak, larvalar tarafından 1 saat aralıklarla yenen yiyecek miktarı, proteinlerinde karbonhidrat (P:C) oranlarına göre farklılık gösteren üç boya etiketli diyetten birine maruz kaldığında kullanılmıştır. Bu protokolün etkinliği larva Drosophila'danörogenetik bir ekran bağlamında gösterilmiştir Farklı makro besin kalitesindeki diyetlerde yenen yiyecek miktarını düzenleyen aday nöronal popülasyonları belirleyerek. Ayrıca test edilen genotipleri fenotipik sınıflar halinde sınıflandırabildik ve gruplandırabildik. Literatürde mevcut yöntemlerin kısa bir incelemesinin yanı sıra, bu yöntemlerin avantajları ve sınırlamaları tartışılır ve ayrıca, bu protokolün diğer özel deneylere nasıl uyarlanabileceği hakkında bazı öneriler sağlanmaktadır.
Introduction
Tüm hayvanlar hayatta kalmak, büyümek ve üremek için gerekli miktarda besin elde etmek için dengeli bir diyete bağımlıdır1. Ne ve ne kadar yiyeceğinin seçimi, tokluk seviyesi gibi hayvanın iç durumu ve gıda kalitesi2 ,3,4,5gibi çevresel koşullarla ilgili çok sayıda etkileşim faktöründen etkilenir. Protein ve karbonhidratlar iki ana makro besin maddesidir ve hayvanların fizyolojik süreçlerini sürdürmek için dengeli alımı esastır. Bu nedenle, beslenme davranışlarını kontrol eden ve bu makro besinlerin dengeli bir şekilde alımını sürdüren sinir mekanizmalarının anlaşılması son derece önemlidir. Bunun nedeni, yaşam süresi, doğurganlık ve metabolik sağlık gibi yaşam öyküsü özelliklerinin protein alımı 6,7 ,8,9,10seviyelerinden doğrudan etkilenmesidir.
Memeliler de dahil olmak üzere karmaşık hayvanlarla evrimsel olarak korunmuş beslenme alışkanlıkları sergileyen daha basit daha çekişli organizmaların kullanımı bu tür çalışmalar için gereklidir. Daha da önemlisi, bu daha basit hayvan modelleri, karmaşık biyolojik soruları maliyetli, etik ve teknik olarak daha etkili bir bağlamda incelerken iyi bir fırsat sağlar. Son yıllarda, Drosophila, güçlü genetik araç seti, karmaşık ve basmakalıp davranışı ve memeliler ile periferik ve besin algılama mekanizmalarının korunmuş mimarisi ile davranışsal nörobiyologlar için verimli bir model olmuştur11. Sonuçta, umut, bu hayvanda yiyecek alımının nasıl düzenlendiğini anlayarak, daha basit bir sinir sistemiyle, daha sonra insan yeme bozukluklarının altında kalan nöronal arızaları çözebilmemizdir.
Beslenme davranışları için nöronal substratların incelenmesi, nöronal aktivitelerini manipüle ederken hayvanların yiyecek alımını aynı anda ölçebilmeye derinden bağlıdır. Yutulmuş minimum miktarda yiyecek nedeniyle, sinekler tarafından yenen yiyecek miktarını ölçmek son derece zordur ve şu anda mevcut olan tüm yöntemler önemli sınırlamalar sunun. Bu nedenle, altın standart tamamlayıcı metodolojilerin bir kombinasyonunu kullanmaktır12. Yetişkin sinekler tarihsel olarak genetik ve davranışsal bir model olarak tercih edilmiştir. Bununla birlikte, Drosophila larvaları, beslenme davranışını kodlayan nöronal substratları araştırmak için fırsatlar da sunar. Yaklaşık 12.000 nöron içeren larva merkezi sinir sistemi (CNS), yaklaşık 150.000 nöron içeren yetişkinden önemli ölçüde daha az karmaşıktır. Bu düşük karmaşıklık sadece sayısal değil, aynı zamanda işlevseldir, çünkü larva davranışları daha basit lokomotif işlevlerine ve duyusal sistemlere dayanır. Sinir sistemlerinin belirgin basitliğine rağmen, larvalar hala tam beslenme davranışları sergiler ve Drosophila larvalarında gıda yutulmasını ölçmek için bazı yöntemlertanımlanmıştır 5,13,14,15. Nöronal aktivite manipülasyonları ile eşleştirerek, Drosophila larvaları gıda alımının sinirsel düzenlemesini anlamak için oldukça çekişli bir model oluşturabilir.
Burada, farklı makro besin kalitesinde diyetlere maruz kalan larvalarda gıda alımını ölçmek için ayrıntılı bir protokol verilmiştir. Makro besin dengeleme diyetleri olarak adlandırılan diyetler, protein ve karbonhidrat içeriğinde, özellikle proteinden karbonhidrata (P:C) oranlarına göre farklılık göstermiştir: Şekil 1A'da gösterildiği gibi 1:1 (protein bakımından zengin diyet), 1:4 (ara diyet) ve 1:16(proteinden fakir diyet). Kısaca, mavi bir gıda boyası ile boyanmış bu üç izokalorik sakkaroz mayası (SY) bazlı diyet kullanılarak nicel bir seçimsiz beslenme tahlili kuruldu. Maya özü ve sakkaroz protein ve karbonhidrat kaynakları olarak kullanıldığından ve her ikisi de karbonhidrat içerdiğinden, daha önce16 olarak açıklandığı gibi ve Şekil 1B'debelirtildiği gibi, bu iki bileşenin dengesi değiştirilerek P:C oranlarında değişim elde edilmiştir. Ana deneysel adımları gösteren protokole şematik bir genel bakış Şekil 2'de mevcuttur.
Bu protokol, farklı P:C oranlarındaki diyetlerde ve termogenetik nöronal ekran bağlamında larva besleme seviyelerinin düzenlenmesinde spesifik nöronal popülasyonların rolünü araştırmak amacıyla kurulmuştur. Geçici Reseptör Potansiyeli (TRP) ailesinden iyi karakterize edilmiş bir nörogenetik araç kullanıldı: Drosophila Geçici Reseptör Potansiyel kanalı (dTRPA1), bir sıcaklık ve voltaj kapılı katyon kanalıdır, ortam sıcaklıkları 25 °C17'ninüzerine çıktığında eylem potansiyellerinin ateşlenmesine izin verir. dTRPA1 transgene'i ifade etmek için, Janelia Araştırma Kampüsü18,19'dakiFlyLight projesi kapsamında Rubin laboratuvarında kurulan Drosophila genomundan cis-düzenleyici bölgelere dayanan Gal4 hatlarından yararlandık.
Burada açıklanan protokol bir aktivasyon ekranı bağlamında kurulmuş olsa da, deneyci tarafından diğer özel ihtiyaçlara veya ilgi alanlarına kolayca uyarlanabilir, yani sıcaklığa duyarlı nöronal susturucu ShibireTS20kullanarak bir bastırma ekranı gerçekleştirmek için dTRPA1'e alternatif olarak. Bu ve diğer uyarlamalar protokol ve tartışma bölümlerinde ele alınmıştır.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Sakkaroz mayası (SY) diyetlerinin hazırlanması
- Makro besin dengeleme ve L3 yetiştirme diyetleri için tüm kuru malzemeleri (agar, maya, sakkaroz) tartın. 1 L yiyecek hazırlamak için gereken bileşenlerin her biri için gram olarak miktarlar Şekil 1B'debelirtilmiştir.
NOT: 60 mm'lik bir Petri kabını doldurmak için yaklaşık 13 mL yiyeceğe ihtiyaç duyulduğunu göz önünde bulundurun. - Tüm malzemeleri steril damıtılmış suda çözün (yiyecekleri hazırlamak için gereken toplam su hacminin yaklaşık% 50'sini kullanın) ve ortamı 5-10 dakika karıştırın.
- 50 dakika boyunca otoklav.
- Ortamların soğumasına izin verdikten sonra, diyetlere sırasıyla% 3 ve% 0.3'lük son konsantrasyonda (v / v) nipagin ve propiyonik asit çözeltileri ekleyin. Makro besin dengeleme diyetlerine, mavi gıda boyasını% 1'lik son konsantrasyona (v / v) ekleyin. Damıtılmış su ile toplam hacimleri tamamlayın.
- Yiyecek diyetlerini 60 mm Petri yemeklerine dikkatlice dökün, böylece dökülen yiyecek miktarı tabakların her birinde yaklaşık olarak aynıdır. Tabakları diyetlerin P:C oranlarıyla etiketle.
NOT: Beslenme tahlil gününde makro besin dengeleme diyetlerini hazırlayın. Mümkün değilse, hazırlanan diyetleri 4 °C'de, kapalı bir kapta maksimum 3 gün boyunca saklayın. Daha uzun depolama süreleri diyeti çok kuru ve sert hale getirir ve larvalar ortama yuva yapamaz.
2. Ebeveyn çizgilerinin genetik çaprazı
NOT: Genetik haçları ayarlamak için Gal4/UAS sistemi21'i kullanın. Bu protokolde, belirli nöronal popülasyonlarda nöronal fonksiyonu etkinleştirmek için, UAS dTRPA1 hattı17'nin dişi bakireleri kullanılmış ve Janelia Gal4 çizgilerinden erkeklere geçmiştir (Şekil 2A). Kullanılan genetik kontrol, dTRPA1 hattı ile Rubin Gal4 koleksiyonunu oluşturmak için kullanılan vektörde Gal4 taşıyan ancak düzenleyici bir parça bulunmayan "boş GAL4" çizgisi arasındaki bir haçın soyuydu (attP2)22. Nöronal baskılamayı teşvik etmek için dTRPA1 yerine ShibireTS20 kodlu bir UAS hattı kullanılabilir.
- Bazı aktif maya macunu ile desteklenmiş L3 yetiştirme diyet plakaları ile 60 mm embriyo toplama kafesleri kurun.
- 5-8 günlük yetişkin UAS dTRPA1 dişi bakireleri ve Janelia Gal4 erkeklerini embriyo toplama kafeslerine aktarın ve çiftleşmenin 24-48 saat boyunca, 25 °C'de, % 60 nem ve 12:12 açık-karanlık döngü ile gerçekleşmesine izin verin (Şekil 2A). 60 mm embriyo toplama kafesleri için haç başına yaklaşık 100 bakire dişi ve 30 erkek kullanın.
- Çiftleşme döneminin sonunda, genetik haçlarda kullanılan L3 yetiştirme diyet plakalarını çıkarın ve atın. Yumurta döşemelerini ve larva evrelemelerini gerçekleştirmek için taze L3 yetiştirme diyet plakaları ile değiştirin.
3. Üçüncü instar larvaların hazırlanması (L3)
- Çift kişilik yetişkin sinekleri taze L3 yetiştirme diyet plakalarına aktarın ve yumurtlamanın 25 °C'de 3-4 saat boyunca gerçekleşmesine izinverin (Şekil 2B). Tüm tabakların diyetin genotipi, P:C oranı ve yumurtanın yumurtlama tarihi ile etiketli olduğundan emin olun.
NOT: Zaman kazanmak için, yumurtaların ekstra işlenmesini önleyen L3 yetiştirme diyetine doğrudan yumurta döşemeyi gerçekleştirin. Küçük ölçekli genetik taramalarda elma suyu agar plakaları kullanılarak yumurtlamanın optimizasyonu elde edilebilir. - Yumurtlama döneminin sonunda, plakaları kafeslerden çıkarın ve plastik kapaklarla örtün. L3 yetiştirme plakalarını desteklemek için maya özü kullanılması durumunda, yumurtlamanın sonundaki tüm artık mayaları çıkardığınızdan emin olun. Bu, larva büyümesi sırasında düzgün olmayan beslenmeyi önlemek için önemlidir.
NOT: Çift kişilik yetişkinler taze L3 yetiştirme diyet plakalarına aktarılabilir, böylece daha fazla yumurta bırakma işlemi yapılır ve daha deneysel larvalar elde edilebilir. Ardışık yumurta bırakmaları, tüm çalışma haftası boyunca aynı yetişkinlerle yapılabilir. - Plaka başına yumurta sayısını tahmin edin ve larva yoğunluğunu plaka başına maksimum 200 embriyoda tutun. Bu tahmin, plakanın dörtte birinde embriyo sayısı sayılarak yapılabilir.
NOT: Aşırı kalabalık bir plaka larva gelişimini geciktirecek ve larva besleme davranışlarını etkileyecektir. - L3 yetiştirme plakalarını 18 °C 'de (izin verilen sıcaklık), % 60 nem ve 12:12 açık-karanlık döngüde kuluçkaya bırakın ve larvaların 9 gün boyunca büyümesine izin verin(Şekil 2B).
- Yumurtlamadan sonraki dokuzuncu günde (AEL), test edilecek genotiplerin her birinden (ve çoğaltmalar için) 10 L3'lük üç grup toplayın. Ayrıca, "sıfır boyalı gıda" kontrolü için 10 L3'lük grupları toplayın. Larva toplamanın, yumurta döşemesini yapmak için kullanılan günün eşdeğer zaman dönemlerinde yapıldığından emin olun (örneğin, yumurtlama 10:00-14:00 arasında gerçekleştiyse, larvaları aynı süre boyunca toplayın 9 gün AEL) ve mümkün olduğunca hafifçe, #5 veya tüy sıklet tokaları kullanılarak gerçekleştirilir. Larvaları bir sonraki adımda belirtildiği gibi doğrudan aktarın (3.6).
NOT: "Sıfır boyalı gıda" kontrol hayvanları, beslenme testinde mavi boya olmadan yiyecek verilen larvalardır. Bu kontrol, larva özlerinin arka plan emiciliğini gidermek için zorunludur. - Toplanan deneysel larvaları 1 mL su içeren plastik çanak ağırlık teknelerine aktarın. Şekil 3'te verilen talimatları izleyerek L2'nin değil, L3'üntoplandığından emin olun.
NOT: L3'ün su veya 1x Fosfat tamponlu salin (PBS) içeren plastik teknelere toplanması, larvaların beslenme testine başlamadan önce iyi nemlendirilmesini sağlamak için önemlidir. Bu, özellikle farklı genotiplerden birkaç deneysel L3 grubu aynı anda toplanıyorsa önemlidir. Her grup için toplama sırasını takip edin, böylece her grup için yiyecek yoksunluğu süresindeki farklılıklar en aza indirilsin. Bu adımda plastik teknelerin kullanımı, larvaların doğrudan su banyosuna yüzmesini sağladığı için isteğe bağlı adım 4.3'ün kullanılmasını kolaylaştırır.
4. Termogenetik aktivasyon ve seçeneksiz besleme tahlilleri
NOT: Sirkadiyen ritimlerle ilgili olası varyasyonları en aza indirmek için beslenme testlerinin günün yaklaşık aynı saatinde yapılması önerilir. Ayrıca, her zaman kontrol deneylerini çalıştırın ("boş Gal4" hattının soyu UAS dTRPA1 ve "sıfır boyalı gıda" larvalarına geçti), ilgi genotiplerine paralel olarak.
- 30 °C'ye (izin vermeyen sıcaklık) bir inkübatör kurun ve yüksek nem seviyelerini (en az% 65) koruyun test sırasında larva dehidratasyonunu önlemek için.
- Besleme testini başlatmadan önce, test plakalarının sıcaklığını 30 ° C'de 30 dakika ısıtarak dengelayın.
- (İsteğe bağlı) Deneysel larvaları 37 ° C su banyosunda 2 dakika ısı şoku. Bu adımı, biraz su içeren plastik ağırlık teknelerindeki hayvanlarla gerçekleştirin.
NOT: Bu adımın amacı, beslenme testinin başlangıcından itibaren nöronların ateşini teşvik ederek nöronal aktivasyonu yoğunlaştırmaktır. - Birden fazla zamanlayıcıyı 1 saat boyunca hazır tutun. Kullanılacak zamanlayıcı sayısı, test edilen deney gruplarının sayısına ve deneycinin larvaları ele almadaki beceriklilik seviyesine bağlıdır.
NOT: Birden çok zamanlayıcının kullanımı, tahlil süresini tüm genotipler için tutarlı tutmak için kritik öneme sahiptir. - Suyu plastik teknelerden dikkatlice boşaltın ve nemlendirilmiş yumuşak bir fırça kullanarak L3 gruplarını teknelerden tahlil plakalarının ortasına hafifçe aktarın. Plakaların kapaklarını geri koyun ve doğru 1-h besleme seanslarını korumak için her plaka (veya plaka grubu) için bir zamanlayıcı başlatın.
- Larvaların karanlıkta 30 °C'de 1 saat boyunca beslenmesine izin verin (Şekil 2C).
NOT: Testlerin karanlıktaki performansı, diyetler aynı boya konsantrasyonunu içermelerine rağmen tonlarda farklılık göstereceğinden, diyetler arasındaki görsel ipuçlarındaki farklılıkları kontrol etmek için önemlidir. - Plakaları bir buz banyosuna aktararak besleme testini durdurun. Plakalar için sabit bir yüzey sağlamak için mümkün olduğunca buzu bastırın.
NOT: Soğuk sıcaklıklar, yuvalama ve kazma davranışlarını engelleyerek beslenmenin sonunu teşvik edecektir. Larvaların çoğu birkaç dakika sonra yemek tabaklarını yüzeye çıkarır ve aşağıdaki adımlarda iyileşmelerini kolaylaştırır.
5. Gıda boyası ekstraksiyonu
- Test edilen her 10 L3 grubu için yaklaşık aynı miktarlarda 0,5 mm cam boncuk (mikrotüp alt kısmını dolduracak kadar) ve 300 μL buz gibi metanol içeren 2 mL mikrotüpler hazırlayın. Mikrotüpleri soğukta tutun, tezgah soğutucusu kullanarak.
DİkKAT: Metanol oldukça yanıcı ve toksiktir. İyi havalandırılmış bir alanda çalışmak ve nitril eldiven giymek de dahil olmak üzere bu reaktifin kullanımı için önerilen tüm güvenlik prosedürlerini izleyin.
NOT: Metanol kullanımı larva örneklerini düzeltmek ve kıtiküldeki melanizasyon reaksiyonlarını önlemek için önemlidir. - #5 veya tüy sıklet-toparlama kullanarak, 10 L3'lük grupları besleme tahlil plakalarından dikkatlice kurtarın ve bir miktar su içeren test plakalarının kapaklarına aktarın. Larvaları durulayarak vücutlarındaki yiyecek kalıntılarını temizleyin ve herhangi bir yaralanmayı önlemek için larvaları nazikçe ele alın. Larva başına ortalama yiyecek alımı miktarının ölçülebilmesi için, çoğaltma başına her genotip için kurtarılan larva sayısının bir kaydını tutun.
NOT: Yaralı larvalar, melanizlenmiş kütiküllere sahip olacakları için, kolorimetrik niceleme için uygun olmadıkları için atılmalıdır. - L3 gruplarını 5.1'de hazırlanan 2 mL mikrotüplere aktarın.
- 5.1 adımında eklenen bir doku lyzer ve cam boncuk kullanarak mekanik bir lizis yöntemiyle gıda boyasını bağırsaklardan çıkarmak için larva dokularını Lyse. (bir doku lyzer mevcut değilse, homojenleştirici bir pestle kullanın). Tercihen, bu adımı 4 °C'de gerçekleştirin (Şekil 2D).
NOT: Bu adımın süresi kullanılan ekipmana bağlı olacaktır. Geleneksel bir doku likörü kullanarak, 1 dk ekstraksiyon yeterlidir. Zaman daralması durumunda, protokol bu adımın sonunda duraklatılabilir ve daha sonra devam edilebilir. Numuneleri -20 °C'de saklayın. - 2 mL mikrotüpleri doğrudan yeni 1,5 mL mikrotüplere ters çevirerek özleri 1,5 mL mikrotüpleri temizlemeye aktarın. Hafifçe gerçekleştirilirse, cam boncukların çoğu 2 mL mikrotüplerin dibinde kalacaktır.
- Özleri 4 °C'de 10 dakika boyunca maksimum hızda santrifüjleyarak hücresel kalıntıları temizleyin.
- 1,5 mL mikrotüpleri temizlemek için süpernatantları toplayın. Hücresel döküntüler süpernatantlarda hala görünüyorsa, 5.6 ve 5.7 adımlarını yineleyin.
6. Gıda tüketiminin kolorimetrik nicelemesi
- Başlangıç mavisi boya çözeltisinin metanolünde seri 1:2 seyreltmeler gerçekleştirerek kalibrasyon eğrisi oluşturmak için standart çözümler hazırlayın. Boş olarak, yalnızca metanol kullanın. Standartların konsantrasyonu, hayvanların yiyecek alım seviyelerine bağlıdır.
NOT: Burada sunulan pilot ekran durumunda, larva özleri için elde edilen boya konsantrasyonları 0,02 ila 1,93 μL/mL arasında değiştiğinden, metanoldeki 2 μL/mL mavi boya çözeltisinin 8 seri seyreltmesinin emicilerinin ölçülmesiyle elde edilen standart bir eğri kullanılmıştır. Gerekirse, deneysel örneklerin boya konsantrasyonuna bağlı olarak bu çözeltilerin konsantrasyonunun artması veya azaltılması. - Deneysel örneklerin 100 μL'sini (adım 5.7'de elde edilir), standartları ve boşu (adım 6.1) 96 kuyulu bir mikro plakanın kuyularına aktarın ve bir plaka okuyucu kullanarak emiciliği 600 nm olarak ölçün (Şekil 2E). Arka plan emiciliğini gidermek için, mavi boya olmadan yiyeceklerle beslenen larvalardan elde edilen özlerin emiciliğini larva özleri için "sıfır" olarak ölçün ("sıfır boyalı gıda kontrolü).
- Standart bir eğri oluşturun ve her deneysel larva grubundan numuneler için elde edilen absorbans değerlerini gıda alımı miktarıyla (mL'deki hacim) ilişkilendirin. 5.2 adımında her grup için toplanan larva sayısını dikkate alarak larva başına ortalama gıda tüketimini bulun
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Drosophila larvaları, fazla karbonhidratları alma pahasına protein alımlarını düzenler23 (Şekil 2E'dekişematik arsa). Aslında, protein alımının bu önceliklendirmesi diğer birçok hayvanda gözlenmiştir veproteinin 24,25.
Bu sağlam beslenme davranışsal tepkiden yararlanarak, makro besin dengelemesinde yer alan nöronal popülasyonları tanımlamayı amaçlayan davranış tabanlı bir ekran tasarlanmıştır. Başka seçeneksiz bir beslenme tahlili kuruldu, L3 gruplarının (grup başına 10 kişi) dTRPA1 kullanarak 1 saat boyunca ve nöronal termogenetik aktivasyon koşulları altında, belirli P:C oranlarını (1:1, 1:4 ve 1:16) içeren üç izocalorik (248 Cal/L) gıda boyalı diyette(Şekil 1 ve Şekil 2C)reklam libitumunu beslemesine izin verilmesinden oluşuyordu. Okuma olarak, farklı P:C oranlarındaki makro besin diyetlerinde yenen ortalama yiyecek miktarı kullanılmıştır. Gal4/UAS sistemi21'den yararlanarak ve FlyLight Project18,19'danbazı Janelia Gal4 hatlarını kullanarak, dTRPA1 ifadesi belirli nöronal popülasyonlarda indüklendi.
Bu protokolde açıklanan yöntemlerle, larva sinir sistemindeki spesifik nöronal popülasyonların termogenetik aktivasyonu altındaki hayvanlar için tüketilen makro besinlerin göreceli miktarını P:C oranları açısından ölçebildik. Bu deneysel yaklaşım, farklı nöron popülasyonlarının aktive etmenin üçüncü instar larvalarda makro besin dengelemesini önemli ölçüde etkilediğini göstermiştir (Şekil 4, Tablo 1). Kontrol hattı için gözlenen beslenme düzeni (attP2), daha düşük P:C oranı diyetlerinde test edilen larvalar tarafından gıda alımının beklenen telafi edici artışını göstererek yöntemin etkinliğini gösterir (Şekil 4'tegri noktalar ve çizgi). Dahası, genotipler ve diyet arasında önemli bir etkileşim bulundu, bu da belirli nöronal popülasyonların termogenetik aktivasyonunun, larvaların diyetin makro besin kalitesine yanıt olarak besin alımlarını düzenleme şeklini değiştirdiği anlamına gelir.
Üç makro besin dengeleme diyetinde (1:1, 1:4, 1:16) test edilen genotiplerin beslenme düzenleri Şekil 4'teki renkli noktalar ve çizgilerle gösterilir ve istatistiksel analiz Tablo 1'demevcuttur.
Aktivasyon ekranında larva sinir sisteminde seyrek olarak ifade edildiği bilinen toplam 36 Janelia Gal4 hattı test edilmiştir. Doğrusal regresyon modellerini kullanarak, hangi genotiplerin genetik kontrol hayvanlarına atıfta bulunarak önemli ölçüde farklı gıda alımı sergilediğini belirledik. Bu farklılıklar, tüm diyetlerde yenen mutlak yiyecek miktarındaki farklılıkları veya makro besin dengeleme yanıtındaki farklılıkları (diyetlerin farklı P:C oranlarına yanıtın eğimi) içeriyordu.
Her üç diyette de R12E06, kontrol hayvanlarından önemli ölçüde daha fazla yiyecek yedi. Ek olarak, gıda alımı ile diyetin P:C oranı arasındaki etkileşim terimindeki önemli bir farkla belirtildiği gibi, ara ve düşük proteinli diyetlerde gıda alımındaki artışı aşırı telafi etti (Tablo 1). R22H01 kontrollerden önemli ölçüde daha fazla yemek yedi, ancak makro besin dengeleme yanıtında farklılık görmedi (Tablo 1). R14B11, R19G11, R21B06, R29C02 ve R48F09 larvaları az miktarda yiyecek yediler ve mevcut diyetin zayıf makro besin kalitesini telafi etme yeteneğini kaybettiler (gıda alımı ile diyetin P:C oranı arasındaki önemli etkileşim terimleriyle belirtildiği gibi, Tablo 1). Son olarak, R45D11 larvaları, 1:1 P:C oranı içeren protein bakımından zengin diyette, ara ve proteinden fakir diyetlere (1:4 ve 1:16) göre önemli ölçüde daha fazla yedi, bu da düşük proteinli diyetlerde beklenebileceklerin tam tersidir.
Bu nedenle, yöntemlerimiz deneysel larvaları, her genotipten, yenilen toplam yiyecek miktarı ve düşük P:C oranı diyetlerinde aşırıya sayılarak protein alımına öncelik verme yeteneği ile ilgili fenotipik sınıflara sınıflandırmamıza izin sağladı. Deney hayvanları için beş fenotipik sınıf kuruldu (Şekil 5): 1 – "Çok yiyin" (kontrol hayvanlarından daha fazla) ve protein seyreltmesi için aşırı telafi; 2 – "Çok yiyin ama normal şekilde telafi edin"; 3 – "Az yiyin (kontrolden daha az) ama telafi edin"; 4 – "Az yiyin ve telafi etmeyin"; 5 – "Sapkın beslenin" (protein bakımından zengin ve ara diyetlerde protein fakiri diyetten daha fazla). Ek olarak, bu fenotip sınıflarının ve genotiplerin her biri için, üçüncü instar larvaların merkezi sinir sistemlerinde GFP kalıplarını gösteriyoruz. Bu bilgiler, tüm Rubin Gal4 ilgi alanlarının ifade kalıplarına erişilebilen FlyLight Project çevrimiçi platformundaki herkese açık görüntüleme verilerinden elde edildi26.

Şekil 1: Protokolümüzde kullanılan sakkaroz maya (SY) diyetleri. (A) Mavi noktalar, beslenme testinde kullanılan ve proteinden karbonhidrat (P:C) oranlarına farklılık gösteren izocalorik (248 kalori/L) makro besin dengeleme diyetlerini temsil eder: 1:1, 1:4 ve 1:16. Bej nokta, 1:2 P:C oranı ve 495 kalori / L kalori yoğunluğu içeren deneysel üçüncü instar larvalarını (L3) geri getirmekiçin kullanılan diyeti temsil eder. Bileşenler tüm diyetler için aynıdır: agar, sakkaroz ve maya. 1 L diyet hazırlamak için gereken bileşenlerin gram miktarı gösterilir. Makro besin dengeleme diyetlerine %1 (v/v) ve L3 yetiştirme diyetine nipagin ve propiyonik asit çözeltilerinin sırasıyla %3 ve %0,3'lük son konsantrasyona (v/v) eklenmesi gerektiğini unutmayın. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 2: Protokolümüzde yer alan ana adımların şematik gösterimi (A) Gal4/UAS sisteminden yararlanarak ebeveyn çizgilerinin genetik çaprazı. Rubin Gal4 hatları ile dTRPA1 kodlayan UAS hattı arasındaki haç, larva merkezi sinir sistemindeki belirli nöronal popülasyonların termogenetik aktivasyonuna izin verir. (B) Deneysel üçüncü instar larvalarının (L3) hazırlanması. Ebeveyn dişilerin 3-4 saat boyunca yumurta bırakmasına izin verildi ve larva evrelemesi 9 gün boyunca izin verilen sıcaklıkta (18 °C) meydana gelir. İsteğe bağlı, besleme testsinden önce 2 dakika boyunca 37 °C'deki ısı şokudur. (C) Nöronal fonksiyonun termogenetik aktivasyonu ve izin vermeyen sıcaklıkta (30 °C) 1 saat boyunca seçeneksiz besleme tahlili. Her genotipten 10 deneysel L3'ten oluşan üç grubun, karbonhidratlara özgü protein (P:C) oranları (1:1, 1:4 ve 1:16) içeren makro besin dengeleme diyetlerinin her birinde beslenmesine izin verildi. (D) Gıda boyası ekstraksiyonu. Mavi gıda boyasını çıkarmak için bir doku lizeri kullanarak larvaların mekanik lizisi. (E) Gıda alımı nicelemesi. Larva özlerindeki gıda boya konsantrasyonu ölçülerek larva başına yenen ortalama yiyecek miktarının kolorimetrik ölçülmesi. Deneysel örneklerin, standartların ve "sıfırın" emiciliği, 96 kuyulu bir plaka okuyucu kullanılarak 600 nm (mavi) olarak ölçüldü. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 3: İkinci (L2) ve üçüncü instars Drosophila larvaları (L3) arasındaki farklar. L2 ve L3, spiracles'in stereomikroskop altında gözlemlenmesiyle kolayca ayırt edilebilir. L2'nin ön spiracles'i kulüp benzeridir, L3'te ise dallıdır. Diğer özellikler iki instarı ayırt etmeye yardımcı olabilir, ancak öznel ve daha az güvenilirdir. L3'ün arka spiracles ucunda L2'de eksik veya zayıf bir şekilde bulunan koyu turuncu bir halka vardır. Nefes borusu L3 larvalarında daha kalındır. Marisa Oliveira tarafından illüstrasyon. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 4: Karbonhidratlara özgü protein (P:C) oranları içeren üç makro besin dengeleme diyetinde nöronal termogenetik aktivasyon koşulları altında larva başına yenen yiyecek miktarı. 1:1, 1:4 ve 1:16'nın belirli P:C oranlarını içeren 3 makro besin dengeleme diyetinde larva başına (mL) yenen ortalama yiyecek miktarı. Her genotipten 10 üçüncü instar larva grubunun, 30 °C'de dTRPA1 kullanarak nöronal termogenetik aktivasyon koşulları altında 1 saat boyunca beslenmesine izin verildi. Test edilen genotipler (Rubin Gal4 çizgileri ile UAS dTRPA1 çizgisi arasındaki genetik haçlardan larva soyları) farklı renklerdeki noktalar ve çizgilerle gösterilir. Genetik bir kontrol olarak (gri ile belirtilir), "boş Gal4" çizgisi (attP2) ve UAS dTRPA1 arasındaki bir haçtan larva soyu kullanılmıştır. Efsanede belirtilen genotiplere verilen isimler, kullanılan "Rubin GAL4" satırlarıyla ilgiliydi. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.

Şekil 5: 5 ana fenotipik sınıfta test edilen çizgilerin gruplandırımı. Sayılarla belirtilen fenotip sınıfları, yenilen toplam yiyecek miktarı ve protein alımı önceliklendirme yanıtını koruma yeteneği açısından gözlenen fenotiplerin kombinasyonuna dayanıyordu: 1 - çok yiyin (kontrol hayvanlarından daha fazla) ve aşırı yiyerek protein seyreltmesini telafi edebildi; 2 – çok yiyin ve telafi edemedik; 3 - az yiyin (kontrolden daha az) ama telafi edin; 4 – az yiyin ve telafi edemedik; ve 5 - larvaların diyetteki protein içeriğinin makro besin seyreltilmesine yanıt olarak beklendiği gibi davranmadığı, protein bakımından zengin ve ara diyetlerde protein fakiri diyetten daha fazla yediği "anormal" olarak adlandırılan ekstra bir fenotipik sınıf. Her genotip için, üçüncü instar larvaların merkezi sinir sistemlerindeki GFP ifade deseni gösterilmiştir. Bu testte kullanılan Rubin Gal4 hatlarının bu görüntüleme verileri kamuya açık FlyLight Project çevrimiçi platformundan çıkarılmıştır26. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
| Anova Tablosu (Tip II testleri) | |||||
| Yanıt: Konsantrasyon/L3 | |||||
| Toplam Metrekare | Df | F değeri | Pr(>F) | ||
| gıda | 0.086832 | 1 | 113.5358 | < 2.2e-16 | *** |
| Genotip | 0.078443 | 10 | 10.2567 | 9.762e-15 | *** |
| Gıda : Genotip | 0.064038 | 10 | 8.3733 | 6.416e-12 | *** |
| Artıklar | 0.215673 | 282 | |||
| Anlam kodları: 0 '***' 0.001 '**' 0.01 '*' 0.05 '.' 0.1 ' ' 1 | |||||
| Özet Tablo (aşağıdaki katsayılar attP kontrol genotipi ile karşılaştırılır): | |||||
| tahmin | Std. Hatası | t değeri | Pr(>||) | ||
| (Kesişme noktası) | 0.064245 | 0.004316 | 14.886 | < 2e-16 | *** |
| gıda | -0.058117 | 0.007206 | -8.066 | 2.10e-14 | *** |
| Genotip R12E06 | 0.040243 | 0.008961 | 4.491 | 1.03e-05 | *** |
| Genotip R14B11 | -0.053347 | 0.014361 | -3.715 | 0.000245 | *** |
| Genotip R19G11 | -0.044880 | 0.010788 | -4.160 | 4.23e-05 | *** |
| Genotip R21B06 | -0.051912 | 0.009363 | -5.544 | 6.79e-08 | *** |
| Genotip R22H01 | 0.017682 | 0.007296 | 2.423 | 0.016004 | * |
| Genotip R29C02 | -0.043102 | 0.011113 | -3.879 | 0.000131 | *** |
| Genotip R40D06 | -0.005341 | 0.009876 | -0.541 | 0.589102 | |
| Genotip R45C03 | 0.004064 | 0.009876 | 0.412 | 0.680997 | |
| Genotip R45D11 | -0.052579 | 0.009876 | -5.324 | 2,08e-07 | *** |
| Genotip R48F09 | -0.044612 | 0.011362 | -3.926 | 0.000108 | *** |
| Gıda : Genotip R12E06 | -0.037763 | 0.015440 | -2.446 | 0.015067 | * |
| Gıda : Genotip R14B11 | 0.058054 | 0.027100 | 2.142 | 0.033031 | * |
| Gıda : Genotip R19G11 | 0.051532 | 0.017726 | 2.907 | 0.003937 | ** |
| Gıda : Genotip R21B06 | 0.054403 | 0.015689 | 3.467 | 0.000607 | *** |
| Gıda : Genotip R22H01 | -0.020863 | 0.012377 | -1.686 | 0.092979 | . |
| Gıda : Genotip R29C02 | 0.048996 | 0.018714 | 2.618 | 0.009317 | ** |
| Gıda : Genotip R40D06 | 0.003804 | 0.016550 | 0.230 | 0.818371 | |
| Gıda : Genotip R45C03 | 0.034117 | 0.016550 | 2.061 | 0.040177 | * |
| Gıda : Genotip R45D11 | 0.090661 | 0.016550 | 5.478 | 9.53e-08 | *** |
| Gıda : Genotip R48F09 | 0.051184 | 0.019045 | 2.688 | 0.007625 | ** |
| Anlam kodları: 0 '***' 0.001 '**' 0.01 '*' 0.05 '.' 0.1 ' ' 1 | |||||
| Artık standart hata: 282 serbestlik derecesinde 0,02765 | |||||
| Çoklu R karesi: 0.516, Ayarlanmış R karesi: 0.4799 | |||||
| F-istatistiği: 21 ve 282 DF'de 14.31, p-değeri: < 2.2e-16 | |||||
Tablo 1: Nöronal termogenetik aktivasyonun ve diyetin makro besin kalitesinin gıda alımı miktarına etkisi için ANOVA tablosu. Kontrol hayvanlarından önemli ölçüde farklı bir besleme davranışı sergileyen genotipleri belirlemek için doğrusal bir model takıldı.
| Genotip | İlişkili Gen | köken | BDSC Stok Numarası |
| w[*] ; P{UAS-trpA1(b). K}attP2 / TM6B, Tb[1] | Bloomington | 26264 | |
| w[1118] ; P{GAL4.1Uw}attP2 | Janelia | 68384 | |
| w[1118] ; P{GMR12E06-GAL4}attP2 | net (CG11450) | Janelia | Na |
| w[1118] ; P{GMR14B11-GAL4}attP2 / TM3, Sb[1] | dnc (CG32498) | Janelia | 49255 |
| w[1118] ; P{GMR19G11-GAL4}attP2 | CG33696 | Janelia | 48864 |
| w[1118] ; P{GMR21B06-GAL4}attP2 | oa2 (CG6919) | Janelia | 49857 |
| w[1118] ; P{GMR22H01-GAL4}attP2 | fru (CG14307) | Janelia | 49001 |
| w[1118] ; P{GMR29C02-GAL4}attP2 | Ptp69D (CG10975) | Janelia | 48088 |
| w[1118] ; P{GMR40D06-GAL4}attP2 | cnc (CG17894) | Janelia | 48616 |
| w[1118] ; P{GMR45C03-GAL4}attP2 | kni (CG4717) | Janelia | 47936 |
| w[1118] ; P{GMR45D11-GAL4}attP2 | pnt (CG17077) | Janelia | 49563 |
| w[1118] ; P{GMR48F09-GAL4}attP2 | dpr8 (CG32600) | Janelia | 50377 |
Tablo 2: Bu çalışmada kullanılan Drosophila çizgileri. Kullanılan tüm satırların ayrıntılı bilgileri: kod adı, genotip, ilişkili gen, köken ve Bloomington Drosophila Stok Merkezi (BDSC) numarası.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Bu protokolle, farklı P:C bileşimindeki diyetlere maruz kaldığında, iki ana makro besin olan protein ve karbonhidratların alım seviyelerini düzenlemek için belirli nöronal popülasyonların termogenetik aktivasyonu altındaki larvaların yeteneğini test edebilirsiniz. Bu yöntem, farklı makro besin kalitesindeki diyetler arasında gıda alımının kontrolü ile ilişkili nöronal popülasyonları tanımlamayı amaçlayan bir larva ön taraması bağlamında test edildi. Bu çalışma ayrıca Drosophila larvalarının besin homeostazına bağlı beslenme davranışlarının nöronal temelini araştırmak için değerli hayvan modelleri olduğunu göstermek için katkıda bulunur.
Protokolde not olarak verilen bilgilere ek olarak, bazı önemli hususları daha fazla tartışmak istiyoruz. Herhangi bir davranışsal tahlilde olduğu gibi, hayvan davranışıyla ilişkili varyasyonu en aza indirmek için deneyci tarafından önlemler alınmalıdır. Akılda tutulması gereken çok önemli bir husus, gelişimsel olarak senkronize hayvanlar elde etmenin önemi ile ilgilidir. Gelişim evrelerinde iyi senkronize olan erken L3 larvalarının kullanımı, hayvanların beslenme sırasında sergiledikleri davranışsal varyasyonları azaltacaktır27. Larvaların senkronizesi, kısa yumurta serimleri ve kültürlerdeki larva yoğunluğunu kontrol ederek elde edilir. Protokolde belirttiğimizlerden (3-4 saat) daha uzun süre yumurtlama kullanmayın. Ayrıca, larva yoğunluğunu plaka başına maksimum 200 hayvana kadar kontrol etmek gelişimsel gecikmeleri önleyecek ve beslenme davranışındaki ek varyasyonu ortadan kaldıracaktır. Lütfen, çiftleşmeden sonra ilk yumurtanın homojenliği korumak ve daha iyi senkronize larva gelişimi elde etmek için atılması gerektiğini unutmayın. Dişiler yumurtaları yumurta kanalında döllediler ve larva toplama arasında tekdüzeliği korumayı zorlaştıran çeşitli gelişim aşamalarında bıraktılar. Son koleksiyondan önce en az ilk saat yumurta toplama plakasının atılması zorunludur. Lütfen, larva taşıma sırasında hayvanlara indüklenen stresin davranışı da olumsuz etkileyebileceğini göz önünde bulundurun. Yumuşak ve su nemlendirilmiş bir fırça kullanarak mümkün olduğunca nazik olmaya çalışın. Son olarak, yüksek sayıda çoğaltmanın daha güvenilir bir veri kümesi oluşturduğunu unutmayın.
Her deneysel protokolde olduğu gibi, yöntemlerimiz bazı sınırlamalar sunar. Hayvanların bağırsaklarında bir gıda boyası birikimine dayanarak gıda alımını ölçmek için kolorimetrik bir yöntem kullanmak, tahlil süresiyle ilgili bazı önlemler içerir. Yetişkin sinekler için, egestion oranının alım oranına eşit olduğu, yöntemin doğruluğunu azalttığı boya birikimi için sabit bir duruma ulaşmanın önemli bir riski olduğu gösterilmiştir28. Larvalarda bunun olduğuna dair bir kanıt olmamasına rağmen, maksimum 60 dakika süreli bir besleme tahlilini yapmaya karar verdik. Bu süre kullanışlı ve yüksek verimli ekranla uyumludur. Ayrıca, protokolün toplam süresini mümkün olduğunca kısa tutmak, bölüm 4, 5 ve 6'daki tüm adımların bir iş gününde tamamlanmasını sağlar. Besleme testinin süresini değiştirmek gerekiyorsa, 60 ila 120 dakika arasında değişen tahlil süreleri, daha önce gösterildiği gibi genotipler arasında gıda alımının verimli bir şekilde ölçülmesine izin vermelidir29. Az miktarda gıda tüketildiğinde gıda ölüm yöntemlerinin hassasiyeti de nispeten düşüktür, bu da çok düşük gıda alımı seviyeleri gösteren genotipler arasındaki çözünürlüğü önemli ölçüde azaltır. Beslenme testlerimizi seçeneksiz bir paradigma kullanarak kurduk. Her bir deneysel larva grubu için sadece bir diyet türü mevcuttur, bu da hayvanların protein ve karbonhidrat tüketim seviyelerini bağımsız olarak düzenlemesine izin vermez. Ayrıca, kimyasal olarak tanımlanmamış diyetler kullandığımız için, larva besleme kalıplarını doğrudan etkileyebilecek besin konsantrasyonlarını kontrol etmek zordur. Bu sorunların üstesinden gelmek veya bir ön ekranda bulunan isabetleri doğrulamak ve daha fazla parçalamak için, deneyci, tanımlanmış sentetik (holidik) ortamlar10 kullanarak ve daha önce açıklandığı gibi gıda seçimi tahlilleri kurarak hassas ve kontrollü bir deneysel beslenme bağlamı oluşturma olasılığını düşünmek isteyebilir30. Termogenetik nöronal modülasyon içeren bir protokol kullanırken, gerekli sıcaklık kaymalarının hayvanların davranışsal çıkışlarını doğrudan etkileyebileceğini göz önünde bulundurmak önemlidir. Optogenetik yaklaşımların tamamlayıcı bir kullanımı, sıcaklığa bağlı yanlış pozitifleri kontrol etmek için ilginç olacaktır, ancak larva besleme tahlilleri bağlamında optogenetik kullanımı teknik olarak zordur, çünkü larvaları beslemek zamanın çoğunu gıda substratında yuvalanmış olarak geçirir.
Bununla birlikte, deneysel yaklaşımımızın çeşitli güçlü yönleri numaralandırılabilir. Yöntemimizin basitliği ve nispeten yüksek verimi, farklı beslenme koşullarına maruz kaldığında birkaç genotip için gıda alımının ölçülmesine izin verir. Larva aşamasında beslenme davranışları yetişkin sineklere göre daha kolay ölçülebilir ve daha iyi işlevsel okumaların üretilmesini sağlar. Ayrıca larvalarda doğal çevreye benzeyen beslenme tahlilleri oluşturmak, daha önce tartışıldığı gibi yetişkinlere göre daha az zordur31. Ayrıca, larvalarda beslenmeyi ölçmek için daha önce belirlenmiş diğer yöntemlerle karşılaştırıldığında, yani belirli bir süre boyunca ağız kancası kasılmalarının sayısını manuel olarak saymaya dayananlar32, kolorimetrik yöntemimiz daha büyük ölçeklerde genetik tarama çalışmalarına olanak tanır. Diğer bazı yöntemler, larvaların oranını bağırsaklarında boyalı yiyeceklerle puanlamaya dayanır, gıda alım seviyelerinin doğru bir şekilde ölçülmesine izin vermez33,34. Nöronal fonksiyonun nörogenetik kontrolü ile ilgili olarak, TRPA1 transgene'in 18 °C'de aktif olmaması, nöronal aktivitenin larva gelişimi boyunca etkilenmemesini sağlar. Bu, deneysel nöronal aktivasyonun larva gelişimi sırasında değil, sadece beslenme tahlilinde gerçekleştirilmesini sağlar. Ek olarak, bir kez daha belirtmek isteriz ki, protokolümüz deneycinin belirli ihtiyaç ve ilgi alanlarına kolayca uyarlanabilir. Örneğin, aktivasyon yerine nöronal fonksiyonun baskılanması, dTRPA1'in sıcaklığa duyarlı nöronal susturucu ShibireTS20'yikodlayan bir UAS hattı için ikame edilmesiyle kolayca elde edilebilir. Ayrıca, deneysel larvalar tarafından sergilenen beslenme seviyeleri çok düşükse, gıda alımını ölçmeyi zorlaştırıyorsa, daha önce açıklandığı gibi, besleme testlerinden önce (protokolün bölüm 4'teki adımlardan önce) 30 dakikalık ekstra bir larva açlığı adımı gerçekleştirmekmümkündür 15. Açlık odaklı davranışların modülatörlerini araştırıyorsanız, bu yiyecek yoksunluğu adımı özellikle ilginç olabilir. Son olarak, önceki çalışmalarda, nicel kolorimetrik yöntemler kullanılarak, yiyecekleri mavi boya ile etiketlemenin beslenme üzerinde hiçbir etkisi olmadığı gösterilmiştir12. Bununla birlikte, tamamlayıcı, daha doğru ve hassas yöntemlerin kullanılmasının, gıdanın radyolabelingi gibi12, bir çalışmanın daha ileri aşamalarında, ön aşamalarda bulunan isabetleri onaylamayı veya daha fazla incelemeyi hedeflemenin yöntemimizin iyi bir tamamlayıcısı olacağını ve deneyci tarafından düşünülmesi gerektiğini düşünüyoruz. Tüm bu nedenlerden dolayı, beslenme davranışlarını kodlayan nöronal devrelerin montajında rol oynayan nöronal popülasyonları tanımlamayı amaçlayan genetik ekranları (özellikle birincil ekranları) gerçekleştirme yöntemlerimizin çekiciliğine inanıyoruz.
Son not olarak, Janelia Araştırma Kampüsü'nde kurulan binlerce larva Gal4 hattının Bloomington Drosophila Stock Center'da halka açık olduğunu ve larva26 ve yetişkin19 CNS ifade kalıpları hakkında çok miktarda bilgiye FlyLight Image Database'de (http://www.janelia.org/gal4-gen1) de herkese açık olduğunu belirtmek isteriz. Bu kaynaklar, Drosophila larvalarında beslenme davranışını düzenleyen nöronların putatif yapı fonksiyonlu nöronal haritalarını detaylandırmayı mümkün klar. Bu, nöronal ekranlarda oluşturulan fenotipik bilgilerin kullanılan sürücülerin ifade kalıplarıyla entegre edilmesiyle mümkündür. Yöntemlerimizin, Drosophila beynindeki makro besin dengelemesi ile ilişkili davranışları beslemek için ön nöronal haritalar oluşturmak için geçerli bir yaklaşım oluşturduğuna inanıyoruz.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Yazarların açıklayacak bir şeyi yok.
Acknowledgments
Instituto Gulbenkian de Ciência'ya (IGC) bu protokolde açıklanan deneysel ekipmanın bir kısmına erişim sağladığı için teşekkür ederiz. Bu çalışma Portekiz Bilim ve Teknoloji Vakfı (FCT), LISBOA-01-0145-FEDER-007660, PTDC/NEU- NMC/2459/2014, IF/00697/2014 ve La Caixa HR17-00595 pmd ve avustralya araştırma konseyi gelecek bursu (FT170100259) tarafından CKM için.
Materials
| Name | Company | Catalog Number | Comments |
| 1.5 mL microtubes | Sarstedt AG & Co. | 72.690.001 | |
| 10xPBS | Nytech | MB18201 | |
| 2.0 mL microtubes | Sarstedt AG & Co. | 72.695.500 | |
| 60 mm petri dishes | Greiner Bio-one, Austria | 628161 | |
| 96 well microplates | Santa Cruz Biotechnology | SC-204453 | |
| Agar | Pró-vida, Portugal | ||
| Bench cooler | Nalgene, USA | Labtop Cooler 5115-0032 | |
| Blue food dye | Rayner, Billingshurst, UK | ||
| Cell disruption media | Scientific Industries, Inc. | 888-850-6208 | (0.5 mm glass beads) |
| Dish weight boats | Santa Cruz Biotechnology | SC-201606 | |
| Embryo collection cage for 60 mm petri dishes | Flystuff, Scientific Laboratory Supplies, UK | FLY1212 (59-100) | |
| Featherweight forceps | BioQuip Products, USA | 4750 | |
| Fly food for stocks maintenance | 1 L food contains: 10 g Agar, 100 g Yeast Extract, 50 g Sucrose, 30 mL Nipagin, 3 mL propionic acid | ||
| Forceps #5 | Dumont | 0108-5-PS | Standard tips, INOX, 11cm |
| Incubator | LMS Ltd, UK | Series 2, Model 230 | For thermogenetic feeding assay (30?C) |
| Incubator | Percival Scientific, USA | DR36NL | To stage larvae (19?C) |
| Janelia lines | Janelia Research Campus | Detailed information in Table 2 | |
| Macronutrient balancing diets | Composition and nutritional information in Figure 1 | ||
| Methanol | VWR | CAS number: 67-56-1 | |
| Nipagin (Methyl 4-hydroxybenzoate) | Sigma-Aldrich | H5501 | |
| Nitrile gloves | VWR, USA | ||
| Refrigerated centrifuge | Eppendorf, Germany | 5804 R / Serial number: 5805CI364293 | |
| Rubin Gal4 ines | Janelia Research Campus | Stoks available at Bloomington Drosophila Stock Center | |
| ShibireTS UAS line | Bloomington Drosophila Stock Center | BDSC number: 66600 | Provided by Carlos Ribeiro Group |
| Soft brushes | For sorting anaesthetised fruit flies | ||
| Spectrophotometer plate reader | Thermo Fisher Scientific | Multiskan Go 51119300 | |
| Stereo microscope | Nikon | 1016625 | |
| Sucrose | Sidul, Portugal | ||
| Third-instar larvae (L3) rearing diet | Composition and nutritional information in Figure 1 | ||
| Timer | |||
| Tissue lyzer / bead beater | MP Biomedicals, USA | FastPrep-24 6004500 | |
| TRPA1 UAS line | Bloomington Drosophila Stock Center | BDSC number: 26264 | Expresses TrpA1 under UAS control; may be used to activate neurons experimentally at 25 ?C |
| Water bath | Sheldon Manufacturing Inc., USA | W20M-2 / 03068308 / 9021195 | |
| Yeast extract | Pró-vida, Portugal | 51% Protein, 15% Carbohydrate |
References
- Raubenheimer, D. Nature of nutrition - a unifying framework from animal adaptation to human. , (2012).
- Carvahlo, M. J. a, Mirth, C. K. Coordinating morphology with behavior during development: an integrative approach from a fly perspective. Frontiers in Ecology and Evolution. , (2015).
- Steck, K., et al. Internal amino acid state modulates yeast taste neurons to support protein homeostasis in Drosophila. Elife. 7, 31625 (2018).
- Itskov, P. M., Ribeiro, C. The dilemmas of the gourmet fly: the molecular and neuronal mechanisms of feeding and nutrient decision making in Drosophila. Frontiers in Neuroscience. 7, 12 (2013).
- Bjordal, M., Arquier, N., Kniazeff, J., Pin, J. P., Leopold, P. Sensing of amino acids in a dopaminergic circuitry promotes rejection of an incomplete diet in Drosophila. Cell. 156 (3), 510-521 (2014).
- Grandison, R. C., Piper, M. D., Partridge, L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature. 462 (7276), 1061-1064 (2009).
- Lee, K. P., et al. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proceedings of the National Academy of Sciences of the United States of America. 105 (7), 2498-2503 (2008).
- Levine, M. E., et al. Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population. Cell Metabolism. 19 (3), 407-417 (2014).
- Solon-Biet, S. M., et al. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metabolism. 19 (3), 418-430 (2014).
- Piper, M. D., et al. A holidic medium for Drosophila melanogaster. Nature Methods. 11 (1), 100-105 (2014).
- Jones, W. D. The expanding reach of the GAL4/UAS system into the behavioral neurobiology of Drosophila. BMB Reports. 42 (11), 705-712 (2009).
- Deshpande, S. A., et al. Quantifying Drosophila food intake: comparative analysis of current methodology. Nature Methods. 11 (5), 535-540 (2014).
- Neckameyer, W. S. A trophic role for serotonin in the development of a simple feeding circuit. Developmental Neuroscience. 32 (3), 217-237 (2010).
- Gasque, G., Conway, S., Huang, J., Rao, Y., Vosshall, L. B. Small molecule drug screening in Drosophila identifies the 5HT2A receptor as a feeding modulation target. Scientific Reports. 3, (2013).
- Schoofs, A., et al. Selection of motor programs for suppressing food intake and inducing locomotion in the Drosophila brain. PLoS Biology. 12 (6), 1001893 (2014).
- Pocas, G. M., Crosbie, A. E., Mirth, C. K. When does diet matter? The roles of larval and adult nutrition in regulating adult size traits in Drosophila melanogaster. Journal of Insect Physiology. , 104051 (2020).
- Hamada, F. N., et al. An internal thermal sensor controlling temperature preference in Drosophila. Nature. 454 (7201), 217-220 (2008).
- Pfeiffer, B. D., et al. Tools for neuroanatomy and neurogenetics in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 105 (28), 9715-9720 (2008).
- Jenett, A., et al. A GAL4-driver line resource for Drosophila neurobiology. Cell Reports. 2 (4), 991-1001 (2012).
- Kitamoto, T. Conditional modification of behavior in Drosophila by targeted expression of a temperature-sensitive shibire allele in defined neurons. Journal of Neurobiology. 47 (2), 81-92 (2001).
- Brand, A. H., Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118 (2), 401-415 (1993).
- Shirangi, T. R., Stern, D. L., Truman, J. W. Motor control of Drosophila courtship song. Cell Reports. 5 (3), 678-686 (2013).
- Mirth, C. M. J. Food intake and food choice are altered by the developmental transition at critical weight in Drosophila melanogaster. Animal Behaviour. 126, 195-208 (2017).
- Simpson, S. J., Raubenheimer, D.
- Raubenheimer, D., Simpson, S. J. Integrative models of nutrient balancing: application to insects and vertebrates. Nutrition Research Reviews. 10 (1), 151-179 (1997).
- Li, H. H., et al. A GAL4 driver resource for developmental and behavioral studies on the larval CNS of Drosophila. Cell Reports. 8 (3), 897-908 (2014).
- Bhatt, P. K., Neckameyer, W. S. Functional analysis of the larval feeding circuit in Drosophila. Journal of Visualized Experiments. (81), e51062 (2013).
- Wong, R., Piper, M. D. W., Blanc, E., Partridge, L. Pitfalls of measuring feeding rate in the fruit fly Drosophila melanogaster. Nature Methods. 5 (3), 214-215 (2008).
- Almeida-Carvalho, M. J., et al. The Ol1mpiad: concordance of behavioural faculties of stage 1 and stage 3 Drosophila larvae. Journal of Experimental Biology. 220, Pt 13 2452-2475 (2017).
- Rodrigues, M. A., et al. Drosophila melanogaster larvae make nutritional choices that minimize developmental time. Journal of Insect Physiology. 81, 69-80 (2015).
- Wong, R., Piper, M. D., Wertheim, B., Partridge, L.
- Wu, Q., et al. Developmental control of foraging and social behavior by the Drosophila neuropeptide Y-like system. Neuron. 39 (1), 147-161 (2003).
- Wu, Q., Zhang, Y., Xu, J., Shen, P. Regulation of hunger-driven behaviors by neural ribosomal S6 kinase in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 102 (37), 13289-13294 (2005).
- Lingo, P. R., Zhao, Z., Shen, P. Co-regulation of cold-resistant food acquisition by insulin- and neuropeptide Y-like systems in Drosophila melanogaster. Neuroscience. 148 (2), 371-374 (2007).
Tags
Nörobilim Sayı 160 Drosophila,larvalar termogenetik nöronal ekran larva besleme davranışı makro besin dengeleme protein karbonhidratlar gıda alımı tahlil gıda boyası kolorimetrik nicelemeErratum
Formal Correction: Erratum: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae
Posted by JoVE Editors on 10/06/2020.
Citeable Link.
An erratum was issued for: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae. A figure was updated.
Figure 1 was updated from:
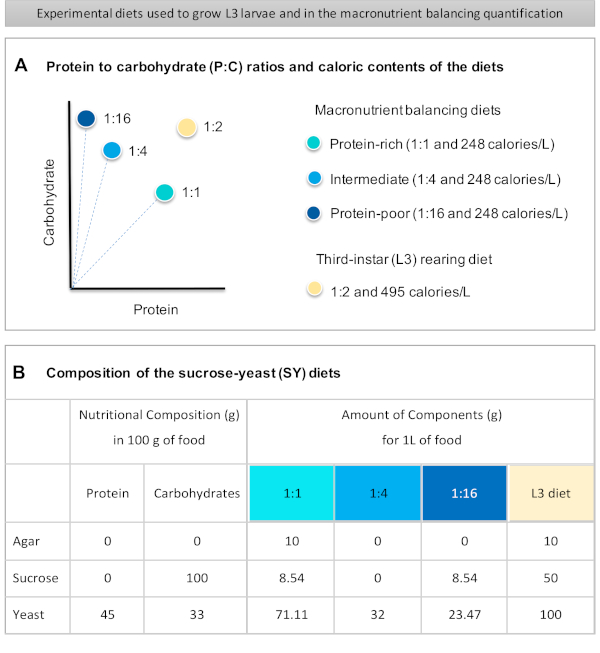
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
to:
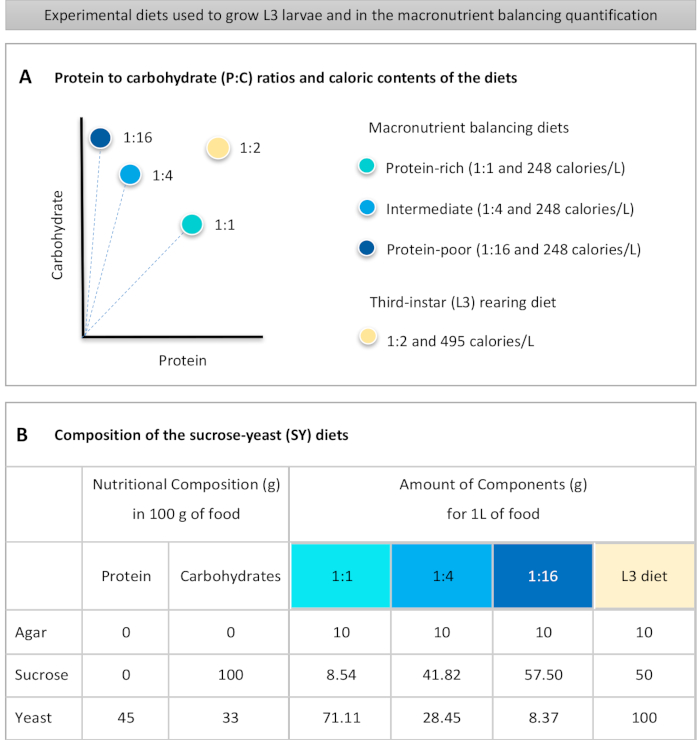
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
