

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Décrit ici est un protocole qui permet la quantification colorimétrique de la quantité de nourriture consommée dans un intervalle de temps défini par les larves de Drosophila melanogaster exposées à des régimes de qualité macronutriment différente. Ces tests sont menés dans le cadre d’un criblage thermogénétique neuronal.
Abstract
Les comportements de recherche de nourriture et d’alimentation permettent aux animaux d’accéder à des sources d’énergie et de nutriments essentiels à leur développement, à leur santé et à leur forme physique. L’étude de la régulation neuronale de ces comportements est essentielle pour la compréhension des mécanismes physiologiques et moléculaires sous-jacents à l’homéostasie nutritionnelle. L’utilisation de modèles animaux génétiquement tractables tels que les vers, les mouches et les poissons facilite grandement ces types d’études. Au cours de la dernière décennie, la mouche des fruits Drosophila melanogaster a été utilisée comme un modèle animal puissant par les neurobiologistes qui étudient le contrôle neuronal des comportements d’alimentation et de recherche de nourriture. Bien que sans aucun doute précieux, la plupart des études examinent les mouches adultes. Ici, nous décrivons un protocole qui tire parti du système nerveux larvaire plus simple pour étudier les substrats neuronaux contrôlant des comportements d’alimentation lorsque les larves sont exposées à des régimes différents dans leur teneur en protéines et en glucides. Nos méthodes sont basées sur un test quantitatif colorimétrique d’alimentation sans choix, réalisé dans le cadre d’un écran d’activation thermogénétique neuronale. Comme lecture, la quantité de nourriture consommée par les larves sur un intervalle de 1 h a été employée une fois exposée à l’un des trois régimes colorant-marqués qui diffèrent dans leurs rapports protéines aux hydrates de carbone (P:C). L’efficacité de ce protocole est démontrée dans le cadre d’un criblage neurogénétique chez la drosophilelarvaire, en identifiant des populations neuronales candidates régulant la quantité de nourriture consommée dans des régimes de différentes qualité macronutriments. Nous avons également pu classer et regrouper les génotypes testés en classes phénotypiques. Sans compter qu’un bref examen des méthodes actuellement disponibles dans la littérature, les avantages et les limites de ces méthodes sont discutés et, également, quelques suggestions sont fournies au sujet de la façon dont ce protocole pourrait être adapté à d’autres expériences spécifiques.
Introduction
Tous les animaux dépendent d’une alimentation équilibrée pour acquérir les quantités nécessaires à la survie, à la croissance et à la reproduction1. Le choix de ce que et combien manger est influencé par une multitude de facteurs d’interaction liés à l’état interne de l’animal, comme le niveau de satiété, et les conditions environnementales, telles que la qualité de la nourriture2,3,4,5. Les protéines et les glucides sont deux macronutriments majeurs et leur apport équilibré est essentiel pour soutenir les processus physiologiques des animaux. Par conséquent, la compréhension des mécanismes neuronaux contrôlant les comportements d’alimentation et soutenant un apport équilibré de ces macronutriments est extrêmement pertinente. C’est parce que les traits du cycle biologique comme la durée de vie, la fécondité et la santé métabolique sont directement affectés par les niveaux d’apport en protéines6,7,8,9,10.
L’utilisation d’organismes plus simples et plus maniables qui présentent des habitudes alimentaires conservées au cours de l’évolution avec des animaux complexes, y compris des mammifères, est essentielle à ce type d’études. Il est important de faire en 1999, ces modèles animaux plus simples offrent une bonne occasion de disséquer des questions biologiques complexes dans un contexte coûteux, éthiquement et techniquement plus efficace. Au cours des dernières décennies, la drosophile,avec sa puissante boîte à outils génétique, son comportement complexe et stéréotypé et son architecture conservée de mécanismes périphériques et de détection des nutriments avec les mammifères, a été un modèle fructueux pour les neurobiologistes comportementaux11. En fin de compte, l’espoir est qu’en comprenant comment l’apport alimentaire est régulé chez cet animal, avec un système nerveux plus simple, nous pouvons alors commencer à démêler les dysfonctionnements neuronaux sous-jacents aux troubles de l’alimentation humaine.
L’étude des substrats neuronaux pour les comportements alimentaires dépend profondément de la capacité de mesurer simultanément l’apport alimentaire des animaux tout en manipulant leur activité neuronale. En raison des quantités minimales d’aliments ingérés, il est extrêmement difficile de quantifier la quantité d’aliments consommés par les mouches, et toutes les méthodes actuellement disponibles présentent des limites importantes. Ainsi, l’étalon-or est d’utiliser une combinaison de méthodologies complémentaires12. Les mouches adultes ont été historiquement favorisées comme modèle génétique et comportemental. Néanmoins, les larves de drosophiles offrent également des possibilités d’étudier les substrats neuronaux codant pour le comportement alimentaire. Le système nerveux central (SNC) larvaire, avec environ 12 000 neurones, est nettement moins complexe que celui de l’adulte, qui contient environ 150 000 neurones. Cette complexité inférieure n’est pas seulement numérique mais aussi fonctionnelle, car les comportements larvaires reposent sur des fonctions de locomotive et des systèmes sensoriels plus simples. Malgré l’apparente simplicité de leur système nerveux, les larves présentent encore des comportements alimentaires complets, et certaines méthodes pour quantifier l’ingestion de nourriture chez les larves de drosophiles ont été décrites5,13,14,15. En s’associant à des manipulations de l’activité neuronale, les larves de drosophiles peuvent constituer un modèle hautement tractable pour comprendre la régulation neuronale de l’apport alimentaire.
Voici un protocole détaillé pour quantifier l’apport alimentaire chez les larves exposées à des régimes alimentaires de différentes qualité de macronutriments. Les régimes, appelés régimes d’équilibrage des macronutriments, différaient par les teneurs en protéines et en glucides, en particulier en ce qui concerne les rapports protéines/glucides (P:C) : 1:1 (régime riche en protéines), 1:4 (régime intermédiaire) et 1:16 (régime pauvre en protéines), comme le montre la figure 1A. Brièvement, une analyse quantitative d’alimentation sans choix a été établie utilisant ces trois régimes basés sur la saccharose-levure isocalorique (SY) teints avec un colorant alimentaire bleu. Étant donné que l’extrait de levure et le saccharose ont été utilisés comme sources de protéines et de glucides, et que les deux contiennent des glucides, la variation des rapports P:C a été obtenue en modifiant l’équilibre de ces deux composants, comme décrit précédemment16 et comme indiqué à la figure 1B. Un aperçu schématique du protocole, montrant les principales étapes expérimentales, est disponible à la figure 2.
Ce protocole a été établi dans le but d’étudier le rôle des populations neuronales spécifiques sur le règlement des niveaux d’alimentation larvaires dans les régimes de différents rapports de P: C et dans le cadre d’un écran neuronal thermogénétique. Un outil neurogénétique bien caractérisé a été utilisé à partir de la famille du potentiel de récepteur transitoire (TRP): canal de potentiel de récepteur transitoire de la drosophile (dTRPA1), qui est un canal cationique à température et tension fermée, permettant la mise à feu des potentiels d’action lorsque les températures ambiantes s’élèvent au-dessus de 25 ° C17. Pour exprimer le transgène dTRPA1, nous avons profité des lignéesGal4 basées sur les régions cis-régulatrices du génome de la drosophile, établies dans le laboratoire Rubin, dans le cadre du projet FlyLight au Janelia Research Campus18,19.
Bien que le protocole, décrit ici, ait été établi dans le cadre d’un écran d’activation, il peut être facilement adapté par l’expérimentateur à d’autres besoins ou intérêts spécifiques, à savoir effectuer un écran de suppression à l’aide du silencieux neuronal sensible à la température ShibireTS20,en alternative au dTRPA1. Cette adaptation et d’autres sont abordées dans les sections protocole et discussion.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Préparation des régimes de levure de saccharose (SY)
- Peser tous les ingrédients secs (gélose, levure, saccharose) pour l’équilibrage des macronutriments et les régimes d’élevage L3. Les quantités en grammes pour chacun des ingrédients nécessaires à la préparation de 1 L d’aliment sont indiquées à la figure 1B.
REMARQUE : Tiendront compte du fait qu’environ 13 mL de nourriture sont nécessaires pour remplir une boîte de Pétri de 60 mm. - Dissoudre tous les ingrédients dans de l’eau distillée stérile (utiliser environ 50% du volume total d’eau nécessaire pour préparer l’aliment) et remuer le milieu pendant 5-10 min.
- Autoclave pendant 50 min.
- Après avoir permis aux milieux de refroidir, ajoutez des solutions de nipagine et d’acide propionique à l’alimentation, à une concentration finale (v/v) de 3 % et 0,3 %, respectivement. Aux régimes d’équilibrage des macronutriments, ajouter le colorant alimentaire bleu à une concentration finale (v/v) de 1 %. Complétez les volumes totaux avec de l’eau distillée.
- Versez soigneusement les régimes alimentaires dans des boîtes de Pétri de 60 mm, de sorte que la quantité de nourriture versée soit approximativement la même dans chacune des assiettes. Étiquetez les plaques avec les rapports P: C des régimes.
REMARQUE : Préparez les régimes d’équilibrage des macronutriments le jour de l’analyse d’alimentation. Si ce n’est pas possible, conservez les régimes préparés à 4 °C, dans un récipient scellé, pendant une durée maximale de 3 jours. Des périodes de stockage plus longues rendent le régime trop sec et trop dur, et les larves ne peuvent pas s’enfouir dans le milieu.
2. Croisement génétique des lignées parentales
REMARQUE : Utilisez le système Gal4/UAS21 pour configurer les croisements génétiques. Dans ce protocole, afin d’activer la fonction neuronale dans des populations neuronales spécifiques, des vierges femelles de la lignée UAS dTRPA117 ont été utilisées et croisées aux mâles à partir des lignées Janelia Gal4(figure 2A). Le contrôle génétique utilisé était la descendance d’un croisement entre la lignée dTRPA1 et une lignée « GAL4 vide », qui porte Gal4 dans le vecteur utilisé pour générer la collection Rubin Gal4 mais sans fragment régulateur présent (attP2)22. Pour favoriser la suppression neuronale, une ligne UAS codant pour ShibireTS20 peut être utilisée, au lieu de dTRPA1.
- Installer des cages de collecte d’embryons de 60 mm avec des plaques de régime d’élevage L3, complétées par de la pâte de levure active.
- Transférer les femelles vierges uas dTRPA1 adultes et les mâles Janelia Gal4, âgés de 5 à 8 jours, dans les cages de collecte d’embryons et permettre à l’accouplement de se produire pendant 24 à 48 h, à 25 °C, avec une humidité de 60 % et un cycle lumière-obscurité de 12 h 12(figure 2A). Pour les cages de collecte d’embryons de 60 mm, utilisez environ 100 femelles vierges et 30 mâles par croisement.
- À la fin de la période d’accouplement, retirez et jetez les plaques de régime d’élevage L3 utilisées dans les croisements génétiques. Remplacez-les par des assiettes fraîches d’élevage L3, afin d’effectuer les pontes et la stadification des larves.
3. Préparation des larves du troisième stade (L3)
- Transférer les mouches adultes accouplées dans des assiettes fraîches d’élevage L3 et laisser la ponte se produire pendant 3 à 4 h, à 25 °C(figure 2B). Assurez-vous que toutes les plaques sont étiquetées avec le génotype, le rapport P: C du régime alimentaire et la date de ponte de l’œuf.
REMARQUE: Pour gagner du temps, effectuez la ponte directement dans le régime d’élevage L3, ce qui évite une manipulation supplémentaire des œufs. Dans le cas de criblages génétiques à petite échelle, l’optimisation de la ponte peut être obtenue en utilisant des assiettes de gélose au jus de pomme. - À la fin de la période de ponte, retirez les plaques des cages et couvrez-les de couvercles en plastique. Dans le cas où l’extrait de levure est utilisé pour compléter les plaques d’élevage L3, assurez-vous d’éliminer toute la levure résiduelle à la fin de la ponte. Ceci est important pour éviter une alimentation non uniforme pendant la croissance larvaire.
REMARQUE: Les adultes accouplés peuvent être transférés dans des assiettes fraîches de régime d’élevage L3, de sorte que plus de pontes d’œufs sont effectuées et que des larves plus expérimentales peuvent être obtenues. Des pontes consécutives peuvent être effectuées avec les mêmes adultes pendant toute une semaine de travail. - Estimer le nombre d’œufs par plaque et maintenir la densité larvaire à un maximum de 200 embryons par plaque. Cette estimation peut être faite en comptant le nombre d’embryons dans un quart de la plaque.
REMARQUE : Une plaque surpeuplée retardera le développement larvaire et affectera les comportements alimentaires larvaires. - Incuber les plaques d’élevage L3 à 18 °C (température permissive), 60 % d’humidité et un cycle lumière-obscurité de 12:12, et permettre aux larves de croître pendant 9 jours(figure 2B).
- Le neuvième jour après la ponte (AEL), prélever trois groupes de 10 L3 dans chacun des génotypes (et pour les répétitions) à tester. De plus, prélever des groupes de 10 L3 pour le contrôle des « aliments sans colorant ». S’assurer que la collecte des larves est effectuée pendant les périodes équivalentes de la journée utilisée pour faire la ponte (p. ex., si la ponte a eu lieu entre 10 h et 14 h, recueillir les larves pendant la même période de temps de 9 jours AEL) et qu’elle est effectuée, aussi doucement que possible, à l’aide d’une pince #5 ou d’une pince de poids plume. Transférer directement les larves comme indiqué à l’étape suivante (3.6).
NOTA : Les animaux témoins « aliments sans colorant » sont des larves qui, dans le test d’alimentation, reçoivent de la nourriture sans colorant bleu. Ce contrôle est impératif pour éliminer l’absorbance de fond des extraits larvaires. - Transférer les larves expérimentales recueillies dans des bateaux de poids de plat en plastique contenant 1 mL d’eau. Assurez-vous que les L3 sont collectés, et non L2, en suivant les instructions données à la figure 3.
REMARQUE: La collecte de L3 sur des bateaux en plastique contenant de l’eau ou 1x solution saline tamponnée au phosphate (PBS) est importante pour garder les larves bien hydratées avant le début de l’analyse d’alimentation. Ceci est particulièrement important si plusieurs groupes L3 expérimentaux de différents génotypes sont collectés en même temps. Gardez une trace de l’ordre de collecte pour chaque groupe, afin de minimiser les différences dans la durée de la privation de nourriture pour chaque groupe. L’utilisation de bateaux en plastique dans cette étape facilite l’étape facultative 4.3 car elle permet aux larves de flotter directement dans le bain-marie.
4. Activation thermogénétique et test d’alimentation sans choix
REMARQUE: Il est recommandé d’effectuer les tests d’alimentation à peu près à la même heure de la journée afin de minimiser les variations possibles liées aux rythmes circadiens. Aussi, toujours exécuter les expériences de contrôle (la progéniture de la ligne « Gal4 vide » croisée à UAS dTRPA1 et les larves « aliments sans colorant »), en parallèle avec les génotypes d’intérêt.
- Mettre en place un incubateur à 30 °C (température non permissive) et maintenir des niveaux élevés d’humidité (au moins 65%) pour éviter la déshydratation larvaire pendant l’essai.
- Avant de commencer l’essai d’alimentation, équilibrer la température des plaques d’essai en les réchauffant à 30 °C pendant 30 min.
- (Facultatif) Choc thermique des larves expérimentales pendant 2 min dans un bain-marie à 37°C. Effectuez cette étape avec les animaux dans les bateaux de poids en plastique contenant de l’eau.
REMARQUE: Le but de cette étape est d’intensifier l’activation neuronale en favorisant la mise à feu des neurones depuis le début du test d’alimentation. - Gardez plusieurs minuteries réglées pour 1 h prêtes. Le nombre de minuteries à utiliser dépend du nombre de groupes expérimentaux testés et du niveau de compétence de l’expérimentateur dans la manipulation des larves.
REMARQUE : L’utilisation de plusieurs minuteries est essentielle pour maintenir la durée de l’essai cohérente pour tous les génotypes. - Égouttez soigneusement l’eau des bateaux en plastique et, à l’aide d’une brosse douce humidifiée, transférez doucement les groupes L3 des bateaux au centre des plaques d’analyse. Remettez les couvercles des plaques et démarrez une minuterie pour chaque plaque (ou groupe de plaques) afin de maintenir des séances d’alimentation précises de 1 h.
- Laisser les larves se nourrir pendant 1 h, à 30 °C, dans l’obscurité(figure 2C).
REMARQUE: La performance de l’essai dans l’obscurité est importante pour contrôler les différences dans les indices visuels entre les régimes, car les régimes diffèrent dans les tons, même s’ils contiennent la même concentration de colorant. - Arrêtez le test d’alimentation en transférant les plaques dans un bain de glace. Appuyez sur la glace autant que possible pour fournir une surface stable pour les plaques.
REMARQUE: Les températures froides favoriseront la fin de l’alimentation en inhibant les comportements d’enfouissement et de creusement. La plupart des larves feront surface dans les assiettes après quelques minutes, facilitant leur récupération dans les étapes suivantes.
5. Extraction de colorants alimentaires
- Préparer des microtubes de 2 mL pour chaque groupe de 10 L3 testés, contenant approximativement les mêmes quantités de billes de verre de 0,5 mm (assez pour remplir la partie inférieure du microtube) et 300 μL de méthanol glacé. Gardez les microtubes dans le froid, à l’aide d’un refroidisseur de banc.
ATTENTION : Le méthanol est très inflammable et toxique. Suivez toutes les procédures de sécurité recommandées pour la manipulation de ce réactif, y compris le travail dans un endroit bien ventilé et le port de gants en nitrile.
REMARQUE: L’utilisation de méthanol est importante pour fixer les échantillons larvaires et éviter les réactions de mélanisation dans la cuticule. - À l’aide de pinces #5 ou de poids plume, récupérez soigneusement les groupes de 10 L3 des plaques d’essai d’alimentation et transférez-les sur les couvercles des plaques d’essai contenant de l’eau. Rincez les larves pour enlever les débris alimentaires sur leur corps tout en manipulant doucement les larves pour éviter toute blessure. Tenir un registre du nombre de larves récupérées pour chaque génotype par répétition, afin que la quantité moyenne d’apport alimentaire par larve puisse être quantifiée.
REMARQUE: Les larves blessées doivent être jetées car elles auront une cuticule mélanisée, ce qui ne convient pas à la quantification colorimétrique. - Transférer les groupes L3 sur les microtubes de 2 mL préparés au 5.1.
- Lyser les tissus larvaires pour extraire le colorant alimentaire des intestins par une méthode de lyse mécanique à l’aide d’un lyseur tissulaire et de billes de verre ajoutées à l’étape 5.1. (si un lyzer tissulaire n’est pas disponible, utilisez un pilon homogénéisant). De préférence, effectuez cette étape à 4 °C(Figure 2D).
REMARQUE: La durée de cette étape dépendra de l’équipement utilisé. À l’aide d’un lyseur tissulaire conventionnel, une extraction de 1 min suffit. En cas de constrictions temporelles, le protocole peut être suspendu à la fin de cette étape et poursuivi ultérieurement. Conserver les échantillons à -20 °C. - Transférer les extraits pour nettoyer les microtubes de 1,5 mL, en inversant directement les microtubes de 2 mL sur les nouveaux microtubes de 1,5 mL. Si elles sont effectuées doucement, la plupart des billes de verre resteront au fond du microtube de 2 mL.
- Dégager les débris cellulaires en centrifugant les extraits, à une vitesse maximale de 10 min, à 4 °C.
- Recueillir les surnageants pour nettoyer les microtubes de 1,5 mL. Si des débris cellulaires sont encore visibles dans les surnageants, répétez les étapes 5.6 et 5.7.
6. Quantification colorimétrique de la consommation alimentaire
- Préparer des solutions étalons, pour générer une courbe d’étalonnage, en effectuant des dilutions en série 1:2 dans le méthanol d’une solution de colorant bleu de départ. Comme blanc, utilisez uniquement du méthanol. La concentration des normes dépend des niveaux d’apport alimentaire des animaux.
NOTA : Dans le cas de l’écran pilote présenté ici, comme les concentrations de colorant obtenues pour les extraits larvaires variaient de 0,02 à 1,93 μL/mL, on a utilisé une courbe standard obtenue en mesurant les absorbances de 8 dilutions en série d’une solution de colorant bleu de 2 μL/mL dans du méthanol. Si nécessaire, augmenter ou diminuer la concentration de ces solutions, selon la concentration de colorant des échantillons expérimentaux. - Transférer 100 μL des échantillons expérimentaux (obtenus à l’étape 5.7), étalons et blancs (étape 6.1) dans les puits d’une microplaque de 96 puits et mesurer l’absorbance à 600 nm, à l’aide d’un lecteur de plaque(figure 2E). Pour éliminer l’absorbance de fond, mesurer l’absorbance des extraits obtenus à partir de larves nourries avec des aliments sans colorant bleu comme un « zéro » pour les extraits larvaires (« contrôle alimentaire zéro colorant »).
- Générer une courbe standard et corréler les valeurs d’absorbance obtenues pour les échantillons de chaque groupe larvaire expérimental avec la quantité d’apport alimentaire (volume en mL). Trouver la consommation alimentaire moyenne par larve en tenant compte du nombre de larves recueillies pour chaque groupe à l’étape 5.2
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Les larves de drosophile régulent leur apport en protéines au prix de l’ingestion de glucides en excès23 (diagramme schématique de la figure 2E). En fait, cette priorisation de l’apport en protéines a été observée chez de nombreux autres animaux et s’appelle la protéine tirant partide 24,25.
Tirant parti de cette réponse comportementale d’alimentation robuste, un écran basé sur le comportement a été conçu dans le but d’identifier les populations neuronales impliquées dans l’équilibrage des macronutriments. Un test d’alimentation sans choix a été établi, qui consistait à permettre à des groupes de L3 (10 individus par groupe) de se nourrir ad libitum pendant 1 heure et dans des conditions d’activation thermogénétique neuronale à l’aide de dTRPA1, dans trois régimes teinés alimentaires isocaloriques (248 Cal/L) contenant des rapports P:C spécifiques (1:1, 1:4 et 1:16)(figure 1 et figure 2C). En lecture, la quantité moyenne de nourriture consommée dans les régimes de macronutriments de différents rapports P:C a été employée. En tirant parti du système Gal4/UAS21 et en utilisant certaines des lignes Janelia Gal4 du Projet FlyLight18,19,l’expression de dTRPA1 a été induite dans des populations neuronales spécifiques.
Avec les méthodes décrites dans ce protocole, nous avons pu quantifier la quantité relative de macronutriments consommés, en termes de rapports P:C, pour des animaux sous activation thermogénétique de populations neuronales spécifiques dans le système nerveux larvaire. Cette approche expérimentale a démontré que l’activation de populations distinctes de neurones affectait considérablement l’équilibrage des macronutriments chez les larves du troisième stade (figure 4, tableau 1). Le profil d’alimentation observé pour la lignée témoin (attP2) démontre l’efficacité de la méthode en montrant une augmentation compensatoire attendue de l’apport alimentaire par les larves testées dans des régimes à rapport P:C inférieur (points gris et ligne à la figure 4). De plus, une interaction significative entre les génotypes et le régime alimentaire a été trouvée, ce qui signifie que l’activation thermogénétique de populations neuronales spécifiques modifie la façon dont les larves régulent leur apport alimentaire en réponse à la qualité macronutriments du régime alimentaire.
Les habitudes alimentaires des génotypes testés dans les trois régimes d’équilibrage des macronutriments (1:1, 1:4, 1:16) sont indiquées par les points et les lignes colorés de la figure 4 et l’analyse statistique est disponible dans le tableau 1.
Dans l’écran d’activation, au total, 36 lignées janelia gal4 connues pour être peu exprimées dans le système nerveux larvaire ont été testées. À l’aide de modèles de régression linéaire, nous avons déterminé quels génotypes présentaient un apport alimentaire significativement différent par rapport aux animaux témoins génétiques. Ces différences comprenaient soit des différences dans la quantité absolue d’aliments consommés dans tous les régimes, soit des différences dans la réponse d’équilibrage des macronutriments (pente de la réponse aux différents rapports P:C des régimes).
Dans les trois régimes, R12E06 a mangé beaucoup plus de nourriture que d’animaux témoins. En outre, il a surcompensé l’augmentation de l’apport alimentaire sur les régimes intermédiaires et faibles en protéines, comme l’indique une différence significative dans le terme d’interaction entre l’apport alimentaire et le rapport P: C de l’alimentation (Tableau 1). R22H01 a mangé beaucoup plus que les témoins, mais n’a pas différé dans la réponse d’équilibrage des macronutriments (Tableau 1). Les larves R14B11, R19G11, R21B06, R29C02 et R48F09 ont mangé de petites quantités de nourriture et ont perdu la capacité de compenser la mauvaise qualité des macronutriments de l’alimentation disponible (comme l’indiquent les termes d’interaction significative entre l’apport alimentaire et le rapport P:C de l’alimentation, tableau 1). Enfin, les larves de R45D11 ont mangé beaucoup plus dans le régime riche en protéines contenant un rapport P:C de 1:1 que dans les régimes intermédiaires et pauvres en protéines (1:4 et 1:16), ce qui est le contraire de ce à quoi on pourrait s’attendre sur les régimes à faible teneur en protéines.
Par conséquent, nos méthodes nous ont permis de classer les larves expérimentales, de chaque génotype, en classes phénotypiques liées à la quantité totale de nourriture consommée et à la capacité de prioriser l’apport en protéines en surconsumant dans les régimes de faible rapport P:C. Cinq classes phénotypiques ont été établies pour les animaux de laboratoire (figure 5): 1 – « Manger beaucoup » (plus que les animaux témoins) et surcompenser pour la dilution des protéines; 2 – « Mangez beaucoup mais compensez normalement » ; 3 – « Mangez peu (moins que le contrôle) mais compensez » ; 4 – « Mangez peu et ne compensez pas » ; 5 – « Mangez de manière aberrante » (plus dans les régimes riches en protéines et intermédiaires que dans le régime pauvre en protéines). De plus, pour chacune de ces classes phénotypiques et génotypes, nous montrons les modèles de GFP dans les systèmes nerveux centraux des larves du troisième stade. Ces informations ont été obtenues à partir des données d’imagerie accessibles au public sur la plate-forme en ligne FlyLight Project, où l’on peut accéder aux modèles d’expression de toutes les lignes d’intérêt Rubin Gal426.

Figure 1 : Les régimes de levure de saccharose (SY) utilisés dans notre protocole. (A) Les points bleus représentent les régimes d’équilibrage des macronutriments isocaloriques (248 calories/L) utilisés dans le test d’alimentation, qui diffèrent par les rapports protéines/glucides (P:C) : 1:1, 1:4 et 1:16. Le point beige représente le régime utilisé pour élever les larves expérimentales du troisième stade (L3), qui ont contenu un rapport P:C de 1:2 et une densité calorique de 495 calories/L.(B)Composition détaillée et informations nutritionnelles des régimes basés sur la levure de saccharose (SY). Les composants sont les mêmes pour tous les régimes: gélose, saccharose et levure. La quantité en grammes des composants nécessaires pour préparer 1 L de régime est indiquée. Notez que 1% (v/v) de colorant bleu doit être ajouté aux régimes d’équilibrage des macronutriments et au régime d’élevage L3, des solutions de nipagin et d’acide propionique doivent être ajoutées à une concentration finale (v/v) de 3% et 0,3%, respectivement. Veuillez cliquer ici pour voir une version plus grande de cette figure.

Figure 2 : Représentation schématique des principales étapes impliquées dans notre protocole (A)Croisement génétique des lignées parentales en tirant parti du système Gal4/UAS. Le croisement entre les raies Rubin Gal4 et la raie UAS codant pour dTRPA1, permet l’activation thermogénétique de populations neuronales spécifiques dans le système nerveux central larvaire. (B) Préparation des larves expérimentales du troisième stade (L3). Les femelles parentales ont été autorisées à pondre pendant 3-4 h et la stadification larvaire se produit à la température permissive (18 °C) pendant 9 jours. En option est le choc thermique à 37 °C pendant 2 min avant le test d’alimentation. (C) Activation thermogénétique de la fonction neuronale et essai d’alimentation sans choix pendant 1 h à la température non permissive (30 °C). Trois groupes de 10 L3 expérimentaux de chaque génotype ont été autorisés à se nourrir dans chacun des régimes d’équilibrage de macronutriments contenant des protéines spécifiques aux rapports de glucides (P: C) (1: 1, 1: 4 et 1: 16). (D) Extraction de colorants alimentaires. Lyse mécanique des larves, à l’aide d’un lyseur tissulaire, pour extraire le colorant alimentaire bleu. E)Quantification de l’absorption alimentaire. Quantification colorimétrique de la quantité moyenne d’aliments consommés par larve en quantifiant la concentration de colorant alimentaire dans les extraits larvaires. L’absorbance des échantillons expérimentaux, des étalons et du « zéro » a été mesurée à 600 nm (bleu), à l’aide d’un lecteur de plaque de 96 puits. Veuillez cliquer ici pour voir une version plus grande de cette figure.

Figure 3 : Différences entre les larves de drosophiles du deuxième (L2) et du troisième stade (L3). Les L2 et L3 peuvent être facilement distingués par l’observation de spiracles sous un stéréomicroscope. Les spiracles antérieurs de L2 sont en forme de club, tandis que dans L3 sont ramifiés. D’autres caractéristiques peuvent aider à distinguer les deux stades, mais sont subjectives et moins fiables. Les spiracles postérieurs de L3 ont un anneau orange foncé à leur extrémité, qui est manquant ou faiblement présent dans le L2. La trachée est plus épaisse chez les larves de L3. Illustration de Marisa Oliveira. Veuillez cliquer ici pour voir une version plus grande de cette figure.

Figure 4 : Quantité d’aliments consommés par larve dans des conditions d’activation thermogénétique neuronale dans trois régimes d’équilibrage des macronutriments contenant des rapports protéines/glucides (P:C) spécifiques. Niveaux moyens de la quantité d’aliments consommés par larve (mL) dans 3 régimes d’équilibrage de macronutriments contenant les rapports P:C spécifiques de 1:1, 1:4 et 1:16. Des groupes de 10 larves du troisième stade, de chaque génotype, ont été autorisés à se nourrir pendant 1 heure, dans des conditions d’activation thermogénétique neuronale, en utilisant dTRPA1, à 30 °C. Les génotypes testés (descendances larvaires des croisements génétiques entre les lignées Rubin Gal4 et la lignée UAS dTRPA1) sont indiqués par des points et des lignes de couleurs différentes. En tant que témoin génétique (indiqué en gris), la descendance larvaire d’un croisement entre la lignée « Gal4 vide » (attP2) et l’UAS dTRPA1 a été utilisée. Les noms donnés aux génotypes, indiqués dans la légende, étaient liés aux lignes « Rubin GAL4 » utilisées. Veuillez cliquer ici pour voir une version plus grande de cette figure.

Figure 5 : Regroupement des lignées testées en 5 classes phénotypiques principales. Les classes phénotypiques indiquées par les nombres étaient basées sur la combinaison des phénotypes observés en termes de quantité totale d’aliments consommés et de capacité à maintenir la réponse de priorisation de l’apport en protéines: 1 - manger beaucoup (plus que les animaux témoins) et ont pu compenser la dilution des protéines en mangeant trop; 2 – manger beaucoup et n’ont pas été en mesure de compenser; 3 - manger peu (moins que le contrôle) mais compenser; 4 – manger peu et n’ont pas été en mesure de compenser; et 5 - une classe phénotypique supplémentaire, appelée « aberrante », dans laquelle les larves ne se sont pas comportées comme prévu en réponse à la dilution en macronutriments de la teneur en protéines dans l’alimentation, mangeant plus dans les régimes riches en protéines et intermédiaires que dans le régime pauvre en protéines. Pour chaque génotype, le modèle d’expression de la GFP dans le système nerveux central des larves du troisième stade est montré. Ces données d’imagerie des lignées Rubin Gal4 utilisées dans ce test ont été extraites de la plateforme en ligne26du projet FlyLight accessible au public. Veuillez cliquer ici pour voir une version plus grande de cette figure.
| Table Anova (essais de type II) | |||||
| Réponse : Concentration/L3 | |||||
| Somme carrée | Df | Valeur F | Pr(>F) | ||
| nourriture | 0.086832 | 1 | 113.5358 | < 2.2e-16 | *** |
| génotype | 0.078443 | 10 | 10.2567 | 9.762e à 15 | *** |
| Alimentation : Génotype | 0.064038 | 10 | 8.3733 | 6.416e-12 | *** |
| Résidus | 0.215673 | 282 | |||
| Codes de signification : 0 '***' 0,001 '**' 0,01 '*' 0,05 '.' 0.1 ' ' 1 | |||||
| Tableau récapitulatif (les coefficients ci-dessous sont comparés au génotype de contrôle de l’attP) : | |||||
| estimer | Erreur std. | Valeur t | Pr(>|t|) | ||
| (Intercepter) | 0.064245 | 0.004316 | 14.886 | < 2e-16 | *** |
| nourriture | -0.058117 | 0.007206 | -8.066 | 2.10e-14 | *** |
| Génotype R12E06 | 0.040243 | 0.008961 | 4.491 | 1.03e-05 | *** |
| Génotype R14B11 | -0.053347 | 0.014361 | -3.715 | 0.000245 | *** |
| Génotype R19G11 | -0.044880 | 0.010788 | -4.160 | 4.23e-05 | *** |
| Génotype R21B06 | -0.051912 | 0.009363 | -5.544 | 6.79e-08 | *** |
| Génotype R22H01 | 0.017682 | 0.007296 | 2.423 | 0.016004 | * |
| Génotype R29C02 | -0.043102 | 0.011113 | -3.879 | 0.000131 | *** |
| Génotype R40D06 | -0.005341 | 0.009876 | -0.541 | 0.589102 | |
| Génotype R45C03 | 0.004064 | 0.009876 | 0.412 | 0.680997 | |
| Génotype R45D11 | -0.052579 | 0.009876 | -5.324 | 2.08e-07 | *** |
| Génotype R48F09 | -0.044612 | 0.011362 | -3.926 | 0.000108 | *** |
| Aliment : Génotype R12E06 | -0.037763 | 0.015440 | -2.446 | 0.015067 | * |
| Aliment : Génotype R14B11 | 0.058054 | 0.027100 | 2.142 | 0.033031 | * |
| Aliment : Génotype R19G11 | 0.051532 | 0.017726 | 2.907 | 0.003937 | ** |
| Aliment : Génotype R21B06 | 0.054403 | 0.015689 | 3.467 | 0.000607 | *** |
| Aliment : Génotype R22H01 | -0.020863 | 0.012377 | -1.686 | 0.092979 | . |
| Aliment : Génotype R29C02 | 0.048996 | 0.018714 | 2.618 | 0.009317 | ** |
| Aliment : Génotype R40D06 | 0.003804 | 0.016550 | 0.230 | 0.818371 | |
| Aliment : Génotype R45C03 | 0.034117 | 0.016550 | 2.061 | 0.040177 | * |
| Aliment : Génotype R45D11 | 0.090661 | 0.016550 | 5.478 | 9.53e-08 | *** |
| Aliment : Génotype R48F09 | 0.051184 | 0.019045 | 2.688 | 0.007625 | ** |
| Codes de signification : 0 '***' 0,001 '**' 0,01 '*' 0,05 '.' 0.1 ' ' 1 | |||||
| Erreur type résiduelle : 0,02765 sur 282 degrés de liberté | |||||
| R au carré multiple : 0,516, R au carré ajusté : 0,4799 | |||||
| Statistique F: 14.31 sur 21 et 282 DF, valeur de p: < 2.2e-16 | |||||
Tableau 1 : Tableau ANOVA pour l’effet de l’activation thermogénétique neuronale et de la qualité des macronutriments de l’alimentation disponible sur la quantité d’apport alimentaire. Un modèle linéaire a été ajusté afin de déterminer les génotypes présentant un comportement alimentaire significativement différent de celui des animaux témoins.
| génotype | Gène associé | origine | Numéro de stock BDSC |
| w[*] ; P{UAS-TrpA1(B). K}attP2 / TM6B, Tb[1] | Bloomington | 26264 | |
| w[1118] ; P{GAL4.1Uw}attP2 | Janelia | 68384 | |
| w[1118] ; P{GMR12E06-GAL4}attP2 | net (CG11450) | Janelia | Na |
| w[1118] ; P{GMR14B11-GAL4}attP2 / TM3, Sb[1] | dnc (CG32498) | Janelia | 49255 |
| w[1118] ; P{GMR19G11-GAL4}attP2 | CG33696 | Janelia | 48864 |
| w[1118] ; P{GMR21B06-GAL4}attP2 | oa2 (CG6919) | Janelia | 49857 |
| w[1118] ; P{GMR22H01-GAL4}attP2 | fru (CG14307) | Janelia | 49001 |
| w[1118] ; P{GMR29C02-GAL4}attP2 | Ptp69D (CG10975) | Janelia | 48088 |
| w[1118] ; P{GMR40D06-GAL4}attP2 | cnc (CG17894) | Janelia | 48616 |
| w[1118] ; P{GMR45C03-GAL4}attP2 | kni (CG4717) | Janelia | 47936 |
| w[1118] ; P{GMR45D11-GAL4}attP2 | pnt (CG17077) | Janelia | 49563 |
| w[1118] ; P{GMR48F09-GAL4}attP2 | dpr8 (CG32600) | Janelia | 50377 |
Tableau 2 : Lignées de drosophiles utilisées dans ce travail. Informations détaillées sur toutes les lignes utilisées : nom de code, génotype, gène associé, origine et numéro du Bloomington Drosophila Stock Center (BDSC).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Avec ce protocole, on pourrait tester la capacité des larves sous activation thermogénétique de populations neuronales spécifiques à réguler les niveaux d’apport en protéines et en glucides, deux macronutriments majeurs, lorsqu’elles sont exposées à des régimes de composition P:C différente. Cette méthode a été testée dans le cadre d’un dépistage préliminaire des larves visant à identifier les populations neuronales associées au contrôle de l’apport alimentaire dans des régimes de qualité macronutriment différente. Ces travaux contribuent également à démontrer que les larves de drosophiles sont des modèles animaux précieux pour étudier la base neuronale des comportements alimentaires associés à l’homéostasie des nutriments.
Outre les informations fournies sous forme de notes dans le protocole, nous aimerions examiner plus avant certains aspects importants. Comme dans tout test comportemental, des mesures doivent être prises par l’expérimentateur pour minimiser la variation associée au comportement animal. Un aspect très important qu’il faut garder à l’esprit est lié à l’importance d’obtenir des animaux synchronisés sur le plan du développement. L’utilisation de larves L3 précoces, bien synchronisées à leur stade de développement, diminuera les variations comportementales exposées par les animaux lors de l’analyse d’alimentation27. La synchronisation des larves est obtenue par de courtes pontes d’œufs et en contrôlant la densité des larves dans les cultures. N’utilisez pas de périodes de ponte plus longues que celles que nous indiquons dans le protocole (3-4 heures). De plus, le contrôle de la densité larvaire jusqu’à un nombre maximal de 200 animaux par plaque permettra d’éviter les retards de développement et d’éliminer les variations supplémentaires dans le comportement alimentaire. Veuillez noter que le premier œuf pond après l’accouplement, doit être jeté pour maintenir l’homogénéité et obtenir un développement larvaire mieux synchronisé. Les femelles ont fécondé les œufs dans l’oviducte et les pondent à divers stades de développement, ce qui rend difficile le maintien de l’uniformité entre la collection larvaire. Il est impératif qu’au moins la plaque de collecte des œufs de la première heure soit jetée avant la collecte finale. S’il vous plaît, prenez en considération que le stress induit aux animaux pendant la manipulation larvaire peut également avoir un impact négatif sur le comportement. Essayez d’être aussi doux que possible, en utilisant une brosse douce et humidifiée à l’eau. Enfin, gardez à l’esprit qu’un nombre élevé de répliques génère un jeu de données plus fiable.
Comme dans tout protocole expérimental, nos méthodes présentent certaines limites. L’utilisation d’une méthode colorimétrique pour quantifier l’apport alimentaire en fonction de l’accumulation d’un colorant alimentaire dans les intestins des animaux implique certaines précautions liées à la durée de l’essai. Pour les mouches adultes, il a été démontré qu’il existe un risque important d’atteindre un état stable pour l’accumulation de colorants, dans lequel le taux d’égestion est égal au taux d’absorption, réduisant ainsi la précision de la méthode28. Bien qu’il n’y ait aucune preuve de ce qui se passe chez les larves, nous avons décidé d’effectuer un test d’alimentation avec une durée maximale de 60 min. Cette durée est pratique et compatible avec l’écran à haut débit. De plus, le fait de garder la durée totale du protocole aussi courte que possible permet d’achever toutes les étapes des sections 4, 5 et 6 en une seule journée ouvrable. S’il est nécessaire de modifier la durée de l’essai d’alimentation, des durées d’essai allant de 60 à 120 minutes devraient permettre une quantification efficace de l’apport alimentaire entre les génotypes, comme démontré précédemment29. La sensibilité des méthodes de mort des aliments est également relativement faible lorsque de petites quantités d’aliments sont consommées, ce qui réduit considérablement la résolution parmi les génotypes présentant de très faibles niveaux d’apport alimentaire. Nous avons mis en place nos tests d’alimentation en utilisant un paradigme sans choix. Un seul type de régime est disponible pour chaque groupe expérimental de larves, ce qui ne permet pas aux animaux de réguler indépendamment les niveaux de consommation de protéines et de glucides. En outre, parce que nous utilisons des régimes chimiquement non définis, il est difficile de garder le contrôle des concentrations de nutriments qui pourraient affecter directement les modèles d’alimentation des larves. Pour surmonter ces problèmes, ou pour confirmer et disséquer davantage les résultats trouvés sur un écran préliminaire, l’expérimentateur pourrait envisager la possibilité d’établir un contexte nutritionnel expérimental précis et contrôlé, en utilisant des milieux synthétiques (holidiques) définis10 et en mettant en place des tests de choix alimentaire comme décrit précédemment30. Tout en utilisant un protocole impliquant la modulation neuronale thermogénétique, il est important de considérer que les changements de température nécessaires pourraient affecter directement les sorties comportementales des animaux. Une utilisation complémentaire des approches optogénétiques serait intéressante pour contrôler les faux positifs induits par la température, mais l’utilisation de l’optogénétique dans le contexte des essais d’alimentation larvaire est techniquement difficile, car les larves d’alimentation passent la plupart du temps enfouis dans le substrat alimentaire.
Néanmoins, plusieurs points forts de notre approche expérimentale peuvent être énumérés. La simplicité et le débit relativement élevé de notre méthode permettent de quantifier l’apport alimentaire pour plusieurs génotypes lorsqu’ils sont exposés à différentes conditions nutritionnelles. Les comportements alimentaires au stade larvaire sont plus facilement quantifiables que chez les mouches adultes, ce qui permet la génération de meilleures lectures fonctionnelles. Il est également moins difficile d’établir des tests d’alimentation ressemblant à l’environnement naturel chez les larves que pour les adultes, comme on l’a déjà vu31. De plus, par rapport à d’autres méthodes précédemment établies pour quantifier l’alimentation des larves, à savoir celles basées sur le comptage manuel du nombre de contractions des crochets bucco-bouches pendant une certaine période de temps32, notre méthode colorimétrique permet des études de dépistage génétique à plus grande échelle. D’autres méthodes sont simplement basées sur la notation de la proportion de larves avec des aliments tenés dans leurs intestins, ne permettant pas une quantification précise des niveaux d’apport alimentaire33,34. En ce qui concerne le contrôle neurogénétique de la fonction neuronale, le fait que le transgène TRPA1 soit inactif à 18 °C garantit que l’activité neuronale n’est pas affectée tout au long du développement larvaire. Cela garantit que l’activation neuronale expérimentale sera effectuée exclusivement pendant le test d’alimentation et non pendant le développement larvaire. De plus, nous aimerions mentionner, une fois de plus, que notre protocole peut être facilement adapté aux besoins et aux intérêts spécifiques de l’expérimentateur. Par exemple, la suppression de la fonction neuronale, au lieu de l’activation, peut être facilement obtenue en substituant le dTRPA1 à une ligne UAS codant pour le silencieux neuronal sensible à la température ShibireTS20. De plus, si les niveaux d’alimentation présentés par les larves expérimentales sont très faibles, ce qui rend difficile la quantification de l’apport alimentaire, il est possible d’effectuer une étape supplémentaire de 30 min de famine larvaire avant l’analyse d’alimentation (avant les étapes de la section 4 du protocole), comme décrit précédemment15. Cette étape de privation de nourriture peut être particulièrement intéressante si vous étudiez les modulateurs des comportements liés à la faim. Enfin, dans des études antérieures, en utilisant des méthodes colorimétriques quantitatives, il a été démontré que l’étiquetage des aliments avec du colorant bleu n’a aucune influence sur l’alimentation12. Néanmoins, nous pensons que l’utilisation de méthodes complémentaires, plus précises et sensibles, comme le radiomarquage de l’aliment12, à des stades plus avancés d’une étude, visant à confirmer ou à disséquer davantage les résultats trouvés au cours des étapes préliminaires serait un bon complément de notre méthode et devrait être considéré par l’expérimentateur. Pour toutes ces raisons, nous croyons en l’attractivité de nos méthodes pour réaliser des dépistages génétiques (en particulier des écrans primaires) visant à identifier les populations neuronales impliquées dans l’assemblage de circuits neuronaux codant pour les comportements d’alimentation.
Pour terminer, nous aimerions mentionner le fait que des milliers de lignées larvaires Gal4 établies sur le Campus de recherche Janelia sont accessibles au public, au Bloomington Drosophila Stock Center, et qu’une grande quantité d’informations sur les modèles d’expression des larves26 etdes adultes 19 CNS est également accessible au public dans la base de données d’images FlyLight (http://www.janelia.org/gal4-gen1). Ces ressources permettent d’élaborer des cartes neuronales putatives structure-fonction des neurones régulant le comportement alimentaire chez les larves de drosophiles. Ceci est possible en intégrant les informations phénotypiques générées dans les écrans neuronaux avec les modèles d’expression des pilotes utilisés. Nous pensons que nos méthodes constituent une approche valide pour générer des cartes neuronales préliminaires pour les comportements d’alimentation associés à l’équilibrage des macronutriments dans le cerveau de la drosophile.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Les auteurs n’ont rien à divulguer.
Acknowledgments
Nous tenons à remercier l’Instituto Gulbenkian de Ciência (IGC) de nous avoir donné accès à une partie de l’équipement expérimental décrit dans ce protocole. Ce travail a été soutenu par la Fondation portugaise pour la science et la technologie (FCT), LISBOA-01-0145-FEDER-007660, PTDC/NEU- NMC/2459/2014, IF/00697/2014 et La Caixa HR17-00595 à PMD et par une bourse future du Conseil australien de la recherche (FT170100259) à CKM.
Materials
| Name | Company | Catalog Number | Comments |
| 1.5 mL microtubes | Sarstedt AG & Co. | 72.690.001 | |
| 10xPBS | Nytech | MB18201 | |
| 2.0 mL microtubes | Sarstedt AG & Co. | 72.695.500 | |
| 60 mm petri dishes | Greiner Bio-one, Austria | 628161 | |
| 96 well microplates | Santa Cruz Biotechnology | SC-204453 | |
| Agar | Pró-vida, Portugal | ||
| Bench cooler | Nalgene, USA | Labtop Cooler 5115-0032 | |
| Blue food dye | Rayner, Billingshurst, UK | ||
| Cell disruption media | Scientific Industries, Inc. | 888-850-6208 | (0.5 mm glass beads) |
| Dish weight boats | Santa Cruz Biotechnology | SC-201606 | |
| Embryo collection cage for 60 mm petri dishes | Flystuff, Scientific Laboratory Supplies, UK | FLY1212 (59-100) | |
| Featherweight forceps | BioQuip Products, USA | 4750 | |
| Fly food for stocks maintenance | 1 L food contains: 10 g Agar, 100 g Yeast Extract, 50 g Sucrose, 30 mL Nipagin, 3 mL propionic acid | ||
| Forceps #5 | Dumont | 0108-5-PS | Standard tips, INOX, 11cm |
| Incubator | LMS Ltd, UK | Series 2, Model 230 | For thermogenetic feeding assay (30?C) |
| Incubator | Percival Scientific, USA | DR36NL | To stage larvae (19?C) |
| Janelia lines | Janelia Research Campus | Detailed information in Table 2 | |
| Macronutrient balancing diets | Composition and nutritional information in Figure 1 | ||
| Methanol | VWR | CAS number: 67-56-1 | |
| Nipagin (Methyl 4-hydroxybenzoate) | Sigma-Aldrich | H5501 | |
| Nitrile gloves | VWR, USA | ||
| Refrigerated centrifuge | Eppendorf, Germany | 5804 R / Serial number: 5805CI364293 | |
| Rubin Gal4 ines | Janelia Research Campus | Stoks available at Bloomington Drosophila Stock Center | |
| ShibireTS UAS line | Bloomington Drosophila Stock Center | BDSC number: 66600 | Provided by Carlos Ribeiro Group |
| Soft brushes | For sorting anaesthetised fruit flies | ||
| Spectrophotometer plate reader | Thermo Fisher Scientific | Multiskan Go 51119300 | |
| Stereo microscope | Nikon | 1016625 | |
| Sucrose | Sidul, Portugal | ||
| Third-instar larvae (L3) rearing diet | Composition and nutritional information in Figure 1 | ||
| Timer | |||
| Tissue lyzer / bead beater | MP Biomedicals, USA | FastPrep-24 6004500 | |
| TRPA1 UAS line | Bloomington Drosophila Stock Center | BDSC number: 26264 | Expresses TrpA1 under UAS control; may be used to activate neurons experimentally at 25 ?C |
| Water bath | Sheldon Manufacturing Inc., USA | W20M-2 / 03068308 / 9021195 | |
| Yeast extract | Pró-vida, Portugal | 51% Protein, 15% Carbohydrate |
References
- Raubenheimer, D. Nature of nutrition - a unifying framework from animal adaptation to human. , (2012).
- Carvahlo, M. J. a, Mirth, C. K. Coordinating morphology with behavior during development: an integrative approach from a fly perspective. Frontiers in Ecology and Evolution. , (2015).
- Steck, K., et al. Internal amino acid state modulates yeast taste neurons to support protein homeostasis in Drosophila. Elife. 7, 31625 (2018).
- Itskov, P. M., Ribeiro, C. The dilemmas of the gourmet fly: the molecular and neuronal mechanisms of feeding and nutrient decision making in Drosophila. Frontiers in Neuroscience. 7, 12 (2013).
- Bjordal, M., Arquier, N., Kniazeff, J., Pin, J. P., Leopold, P. Sensing of amino acids in a dopaminergic circuitry promotes rejection of an incomplete diet in Drosophila. Cell. 156 (3), 510-521 (2014).
- Grandison, R. C., Piper, M. D., Partridge, L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature. 462 (7276), 1061-1064 (2009).
- Lee, K. P., et al. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proceedings of the National Academy of Sciences of the United States of America. 105 (7), 2498-2503 (2008).
- Levine, M. E., et al. Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population. Cell Metabolism. 19 (3), 407-417 (2014).
- Solon-Biet, S. M., et al. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metabolism. 19 (3), 418-430 (2014).
- Piper, M. D., et al. A holidic medium for Drosophila melanogaster. Nature Methods. 11 (1), 100-105 (2014).
- Jones, W. D. The expanding reach of the GAL4/UAS system into the behavioral neurobiology of Drosophila. BMB Reports. 42 (11), 705-712 (2009).
- Deshpande, S. A., et al. Quantifying Drosophila food intake: comparative analysis of current methodology. Nature Methods. 11 (5), 535-540 (2014).
- Neckameyer, W. S. A trophic role for serotonin in the development of a simple feeding circuit. Developmental Neuroscience. 32 (3), 217-237 (2010).
- Gasque, G., Conway, S., Huang, J., Rao, Y., Vosshall, L. B. Small molecule drug screening in Drosophila identifies the 5HT2A receptor as a feeding modulation target. Scientific Reports. 3, (2013).
- Schoofs, A., et al. Selection of motor programs for suppressing food intake and inducing locomotion in the Drosophila brain. PLoS Biology. 12 (6), 1001893 (2014).
- Pocas, G. M., Crosbie, A. E., Mirth, C. K. When does diet matter? The roles of larval and adult nutrition in regulating adult size traits in Drosophila melanogaster. Journal of Insect Physiology. , 104051 (2020).
- Hamada, F. N., et al. An internal thermal sensor controlling temperature preference in Drosophila. Nature. 454 (7201), 217-220 (2008).
- Pfeiffer, B. D., et al. Tools for neuroanatomy and neurogenetics in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 105 (28), 9715-9720 (2008).
- Jenett, A., et al. A GAL4-driver line resource for Drosophila neurobiology. Cell Reports. 2 (4), 991-1001 (2012).
- Kitamoto, T. Conditional modification of behavior in Drosophila by targeted expression of a temperature-sensitive shibire allele in defined neurons. Journal of Neurobiology. 47 (2), 81-92 (2001).
- Brand, A. H., Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118 (2), 401-415 (1993).
- Shirangi, T. R., Stern, D. L., Truman, J. W. Motor control of Drosophila courtship song. Cell Reports. 5 (3), 678-686 (2013).
- Mirth, C. M. J. Food intake and food choice are altered by the developmental transition at critical weight in Drosophila melanogaster. Animal Behaviour. 126, 195-208 (2017).
- Simpson, S. J., Raubenheimer, D.
- Raubenheimer, D., Simpson, S. J. Integrative models of nutrient balancing: application to insects and vertebrates. Nutrition Research Reviews. 10 (1), 151-179 (1997).
- Li, H. H., et al. A GAL4 driver resource for developmental and behavioral studies on the larval CNS of Drosophila. Cell Reports. 8 (3), 897-908 (2014).
- Bhatt, P. K., Neckameyer, W. S. Functional analysis of the larval feeding circuit in Drosophila. Journal of Visualized Experiments. (81), e51062 (2013).
- Wong, R., Piper, M. D. W., Blanc, E., Partridge, L. Pitfalls of measuring feeding rate in the fruit fly Drosophila melanogaster. Nature Methods. 5 (3), 214-215 (2008).
- Almeida-Carvalho, M. J., et al. The Ol1mpiad: concordance of behavioural faculties of stage 1 and stage 3 Drosophila larvae. Journal of Experimental Biology. 220, Pt 13 2452-2475 (2017).
- Rodrigues, M. A., et al. Drosophila melanogaster larvae make nutritional choices that minimize developmental time. Journal of Insect Physiology. 81, 69-80 (2015).
- Wong, R., Piper, M. D., Wertheim, B., Partridge, L.
- Wu, Q., et al. Developmental control of foraging and social behavior by the Drosophila neuropeptide Y-like system. Neuron. 39 (1), 147-161 (2003).
- Wu, Q., Zhang, Y., Xu, J., Shen, P. Regulation of hunger-driven behaviors by neural ribosomal S6 kinase in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 102 (37), 13289-13294 (2005).
- Lingo, P. R., Zhao, Z., Shen, P. Co-regulation of cold-resistant food acquisition by insulin- and neuropeptide Y-like systems in Drosophila melanogaster. Neuroscience. 148 (2), 371-374 (2007).
Tags
Neurosciences Numéro 160 Drosophile larves dépistage neuronal thermogénétique comportement d’alimentation des larves équilibrage des macronutriments protéines glucides analyse de l’apport alimentaire colorant alimentaire quantification colorimétriqueErratum
Formal Correction: Erratum: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae
Posted by JoVE Editors on 10/06/2020.
Citeable Link.
An erratum was issued for: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae. A figure was updated.
Figure 1 was updated from:
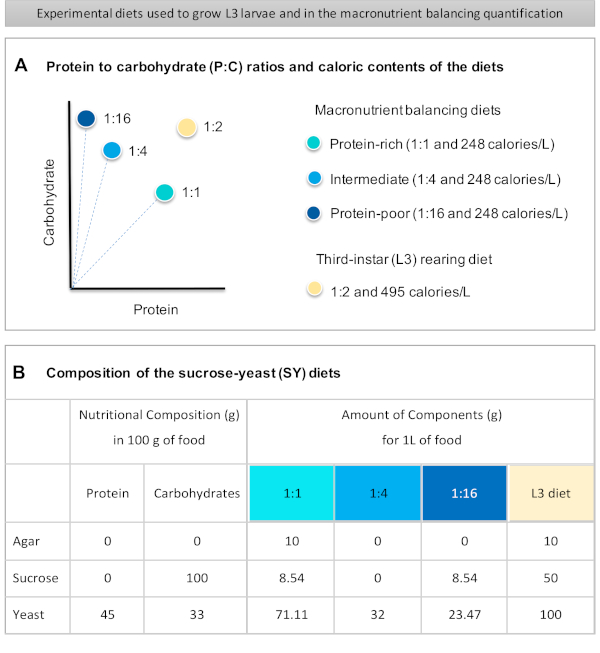
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
to:
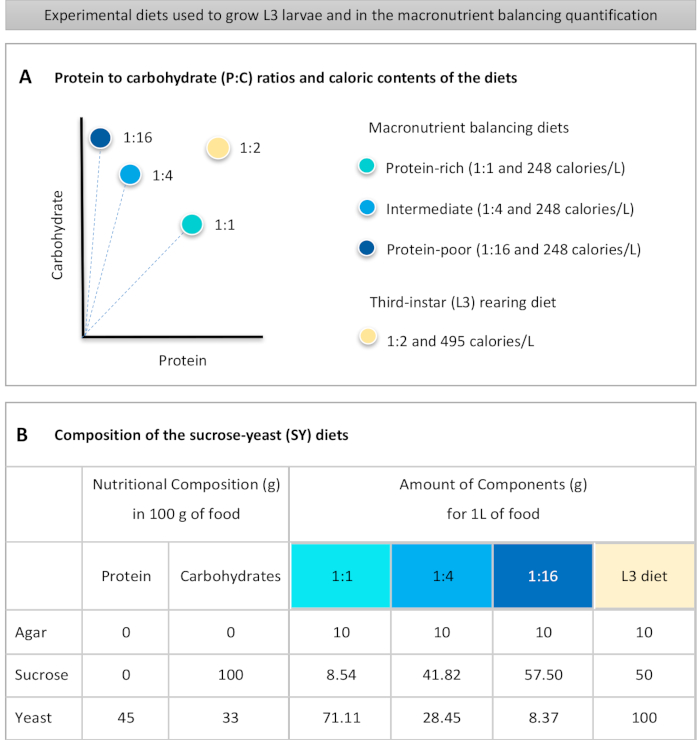
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
