

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Beskrivs här är ett protokoll som möjliggör kolorimetrisk kvantifiering av mängden mat som äts inom ett definierat tidsintervall av Drosophila melanogaster larver utsatta för dieter av olika makronäringsämne kvalitet. Dessa analyser utförs i samband med en neuronal termogenetisk skärm.
Abstract
Foderbeteenden gör det möjligt för djur att få tillgång till energikällor och näringsämnen som är nödvändiga för deras utveckling, hälsa och kondition. Att undersöka neuronal reglering av dessa beteenden är avgörande för förståelsen av de fysiologiska och molekylära mekanismerna bakom näringsmässiga homeostas. Användningen av genetiskt dragbara djurmodeller som maskar, flugor och fisk underlättar i hög grad dessa typer av studier. Under det senaste decenniet har fruktflugan Drosophila melanogaster använts som en kraftfull djurmodell av neurobiologer som undersöker neuronal kontroll av utfodring och 40-bekämpning. Även om det utan tvekan är värdefullt, undersöker de flesta studier vuxna flugor. Här beskriver vi ett protokoll som utnyttjar det enklare larv nervsystemet för att undersöka neuronala substrat som styr utfodringsbeteenden när larver utsätts för dieter som skiljer sig åt i deras protein- och kolhydratinnehåll. Våra metoder är baserade på en kvantitativ kolorimetrisk no-choice utfodringsanalys, utförs i samband med en neuronal termogenetisk aktiveringsskärm. Som en avläsning användes mängden mat som äts av larver under ett 1 h intervall när den exponerades för en av de tre färgmärkta dieterna som skiljer sig åt i förhållandet mellan protein och kolhydrater (P:C). Effekten av detta protokoll visas i samband med en neurogenetisk skärm i larv Drosophila, genom att identifiera kandidat neuronal populationer reglera mängden mat som äts i dieter av olika makronäringsämnen kvalitet. Vi kunde också klassificera och gruppera genotyperna som testats i fenotypiska klasser. Förutom en kort genomgång av de för närvarande tillgängliga metoderna i litteraturen diskuteras fördelarna och begränsningarna med dessa metoder och dessutom ges några förslag om hur detta protokoll kan anpassas till andra specifika experiment.
Introduction
Alla djur är beroende av en balanserad kost för att förvärva de nödvändiga mängderna näringsämnen för överlevnad, tillväxt och reproduktion1. Valet av vad och hur mycket man ska äta påverkas av en mängd interagerande faktorer relaterade till djurets inre tillstånd, som mättnadsnivån och miljöförhållandena, såsom livsmedelskvalitet2,3,4,5. Protein och kolhydrater är två stora makronäringsämnen och dess balanserade intag är avgörande för att upprätthålla djurens fysiologiska processer. Därför är förståelsen av de neurala mekanismerna som styr utfodringsbeteenden och upprätthåller ett balanserat intag av dessa makronäringsämnen extremt relevant. Detta beror på att livshistoria egenskaper som livslängd, fecundity och metabolisk hälsa påverkas direkt av nivåerna av proteinintagintag 6,7,8,9,10.
Användningen av enklare mer lätthanterliga organismer som uppvisar evolutionärt bevarade utfodringsvanor med komplexa djur, inklusive däggdjur, är avgörande för denna typ av studier. Viktigt är att dessa enklare djurmodeller ger ett bra tillfälle att dissekera komplexa biologiska frågor i ett kostsamt, etiskt och tekniskt effektivare sammanhang. Under de senaste decennierna har Drosophila, med sin kraftfulla genetiska verktygslåda, invecklade och stereotypa beteende och bevarad arkitektur av perifera och näringsavkännande mekanismer med däggdjur, varit en fruktbar modell för beteendemässiga neurobiologer11. I slutändan är förhoppningen att genom att förstå hur matintag regleras i detta djur, med ett enklare nervsystem, kan vi sedan börja reda ut neuronala funktionsfel som ligger till grund för mänskliga ätstörningar.
Studien av neuronala substrat för utfodringsbeteenden är djupt beroende av att samtidigt kunna mäta djurens matintag samtidigt som deras neuronala aktivitet manipuleras. På grund av de minimala mängder mat som intas är kvantifiering av mängden mat som äts av flugor extremt utmanande, och alla metoder som för närvarande finns tillgängliga utgör betydande begränsningar. Således är guldstandarden att använda en kombination av kompletterande metoder12. Vuxna flugor har historiskt gynnats som en genetisk och beteendemässig modell. Ändå erbjuder Drosophila larver också möjligheter att undersöka neuronala substrat som kodar utfodringsbeteende. Larvens centrala nervsystem (CNS), med cirka 12 000 nervceller, är betydligt mindre komplext än det vuxna, som innehåller cirka 150 000 nervceller. Denna lägre komplexitet är inte bara numerisk utan också funktionell, eftersom larvbeteenden förlitar sig på enklare lokfunktioner och sensoriska system. Trots den uppenbara enkelheten i deras nervsystem uppvisar larver fortfarande fullständiga utfodringsbeteenden, och vissa metoder för att kvantifiera matintag i Drosophila larver har beskrivits5,13,14,15. Genom att para ihop med manipuleringar av neuronal aktivitet kan Drosophila larver utgöra en mycket lätthanterlig modell för att förstå neural reglering av matintag.
Här finns ett detaljerat protokoll för att kvantifiera födointag i larver som utsätts för dieter av olika makronäringsämneskvalitet. Dieterna, så kallade makronäringsämnen balanserande dieter, skilde sig åt i protein- och kolhydratinnehållet, särskilt när det gäller förhållandet mellan protein och kolhydrater (P:C), som visas i figur 1A. Kortfattat fastställdes en kvantitativ no-choice utfodringsanalys med hjälp av dessa tre isocaloric sackarosjäst (SY)-baserade dieter färgade med ett blått matfärgämne. Eftersom jästextrakt och sackaros användes som protein- och kolhydratkällor, och båda innehåller kolhydrater, erhölls variation i P:C-kvoterna genom att ändra balansen mellan dessa två komponenter, som tidigare beskrivits16 och som anges i figur 1B. En schematisk översikt över protokollet, som visar de viktigaste experimentella stegen, finns i figur 2.
Detta protokoll inrättades i syfte att undersöka rollen av specifika neuronal populationer på regleringen av larv utfodring nivåer i dieter av olika P:C förhållanden och i samband med en termogenetisk neuronal skärm. Ett väl karakteriserat neurogenetiskt verktyg användes från transientreceptorpotentialfamiljen (TRP) : Drosophila Transient Receptor Potential channel (dTRPA1), som är en temperatur- och spänningsportad katjonkanal, vilket möjliggör avfyrning av åtgärdspotentialer när omgivningstemperaturen stiger över 25 °C17. För att uttrycka dTRPA1-transgenen utnyttjade vi Gal4-linjerna baserade på cis-regulatoriskaregioner från Drosophila-genomet, etablerat i Rubin-laboratoriet, inom ramen för FlyLight-projektet vid Janelia Research Campus18,19.
Även om protokollet, här beskrivet, har upprättats i samband med en aktiveringsskärm, kan det enkelt anpassas av experimenteraren till andra specifika behov eller intressen, nämligen att utföra en undertryckande skärm med den temperaturkänsliga neuronala ljuddämparen ShibireTS20, i alternativ till dTRPA1. Denna och andra anpassningar diskuteras i protokoll- och diskussionsavsnitten.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Beredning av sackarosjäst (SY) dieter
- Väg alla torra ingredienser (agar, jäst, sackaros) för makronäringsämnena balansering och L3-uppfödningsdieter. Mängderna i gram för var och en av de ingredienser som behövs för att förbereda 1 L mat anges i figur 1B.
OBS: Ta hänsyn till att cirka 13 ml mat behövs för att fylla en 60 mm Petri-skål. - Lös upp alla ingredienser i sterilt destillerat vatten (använd cirka 50% av den totala volymen vatten som behövs för att förbereda maten) och rör om mediet i 5-10 min.
- Autoklav i 50 min.
- Efter att ha tillåtit mediumen att svalna, tillsätt nipagin- och propionsyralösningar till dieterna, vid en slutlig koncentration (v/ v) på 3% respektive 0,3%. Till makronäringsämnena balansera dieter, tillsätt blått matfärgämne till en slutlig koncentration (v/v) på 1%. Fyll i de totala volymerna med destillerat vatten.
- Häll försiktigt matdieten till 60 mm Petri-rätter, så att mängden mat som hälls är ungefär densamma i var och en av plattorna. Märk plattorna med P:C-förhållandet mellan dieterna.
OBS: Förbered de makronäringsämnena balansering dieter på dagen för utfodringsanalysen. Om det inte är möjligt, förvara de beredda dieterna vid 4 °C, i en förseglad behållare, under en maximal varaktighet av 3 dagar. Längre lagringsperioder gör kosten för torr och hård, och larverna kan inte gräva in i mediet.
2. Genetiska kors av föräldralinjer
OBS: Använd Gal4/UAS-systemet21 för att ställa in de genetiska korsen. I detta protokoll, för att aktivera neuronal funktion i specifika neuronala populationer, kvinnliga oskulder i UAS dTRPA1 linje17 användes och korsades till män från Janelia Gal4 linjer (Figur 2A). Den genetiska kontrollen som användes var avkomman till en korsning mellan dTRPA1-linjen och en "tom GAL4" -linje, som bär Gal4 i vektorn som används för att generera Rubin Gal4-samlingen men utan något regulatoriskt fragment närvarande (attP2)22. För att främja neuronal suppression kan en UAS-linjekodning ShibireTS20 användas, istället för dTRPA1.
- Ställ in 60 mm embryosamlingsburar med L3-uppfödningsdietplattor, kompletterade med lite aktiv jästpasta.
- Överför de vuxna UAS dTRPA1 kvinnliga oskulderna och Janelia Gal4-hanarna i åldern 5-8 dagar till embryosamlingsburarna och låt parningen ske i 24-48 timmar, vid 25 °C, med 60 % luftfuktighet och en ljusmörk cykel på 12:12(figur 2A). För 60 mm embryosamlingsburar, använd cirka 100 jungfruliga honor och 30 hanar per kors.
- I slutet av parningsperioden, ta bort och kassera L3-uppfödningsdietplattorna som används i de genetiska korsen. Byt ut dem mot färska L3-uppfödningsplattor för att utföra äggläggningen och larv iscensättningen.
3. Beredning av tredje-instar larver (L3)
- Överför de parade vuxna flugorna till färska L3-uppfödningsdieter och låt äggläggningen ske i 3-4 timmar vid 25 °C (figur 2B). Se till att alla tallrikar är märkta med genotypen, P:C-förhållandet mellan kosten och äggets datum.
OBS: För att spara tid, utför äggläggningen direkt i L3-uppfödningsdieten, vilket undviker extra hantering av äggen. Vid småskaliga genetiska screenings kan optimering av äggläggningen erhållas med hjälp av äppeljuiceagarplattor. - I slutet av äggläggningsperioden, ta bort plattorna från burarna och täck dem med plastlock. Om jästextrakt används för att komplettera L3-uppfödningsplattorna, se till att ta bort all restjäst i slutet av äggläggningen. Detta är viktigt för att undvika ojämn utfodring under larvtillväxten.
OBS: De parade vuxna kan överföras till färska L3-uppfödningsdietplattor, så fler äggläggningar utförs och mer experimentella larver kan erhållas. På varandra följande äggläggningar kan utföras med samma vuxna under en hel arbetsvecka. - Uppskatta antalet ägg per tallrik och håll larvtätheten till högst 200 embryon per tallrik. Denna uppskattning kan göras genom att räkna antalet embryon i en fjärdedel av plattan.
OBS: En överfull platta kommer att fördröja larvutvecklingen och påverka larvmatningsbeteenden. - Inkubera L3-uppfödningsplattorna vid 18 °C (tillåtande temperatur), 60 % luftfuktighet och en ljusmörk cykel på 12:12 och låt larverna växa i 9 dagar (figur 2B).
- På nionde dagen efter äggläggning (AEL), samla tre grupper på 10 L3 från var och en av genotyperna (och för replikat) som ska testas. Samla dessutom grupper på 10 L3 för kontrollen "nollfärgningsmedel". Se till att larvsamlingen görs under motsvarande tidsperioder på dagen som används för att göra ägget lägg (t.ex. om äggläggningen inträffade mellan 10:00-14:00, samla larverna under samma tidsperiod 9 dagar AEL) och utförs så försiktigt som möjligt med hjälp av tång #5 eller en fjäderviktstång. Överför larverna direkt enligt nästa steg (3.6).
OBS: Kontrolldjuren "nollfärgsmat" är larver som i utfodringstestet ges mat utan blåfärgningsmedel. Denna kontroll är absolut nödvändig för att ta bort bakgrundsabsorbansen av th larv extrakt. - Överför de insamlade experimentella larverna till plastfatsviktbåtar som innehåller 1 ml vatten. Se till att L3 samlas in, och inte L2, genom att följa anvisningarna i figur 3.
OBS: Insamling av L3 till plastbåtar som innehåller vatten eller 1x fosfatbuffrad saltlösning (PBS), är viktigt för att hålla larverna väl hydratiserade innan utfodringstestet börjar. Detta är särskilt viktigt om flera experimentella L3-grupper från olika genotyper samlas in samtidigt. Håll reda på insamlingsordern för varje grupp, så att skillnader i varaktigheten av matbrist för varje grupp minimeras. Användningen av plastbåtar i detta steg underlättar det valfria steg 4.3 eftersom det gör det möjligt för larver att flyta direkt i vattenbadet.
4. Termogenetisk aktivering och valfri utfodringsanalys
OBS: Det rekommenderas att utföra utfodringsanalyserna vid ungefär samma tid på dagen för att minimera eventuella variationer relaterade till dygnsrytmen. Kör också alltid kontrollexperimenten (avkomman till den "tomma Gal4"-linjen korsade till UAS dTRPA1 och larverna "nollfärgningsmedel" ), parallellt med genotyperna av intresse.
- Ställ in en inkubator på 30 °C (icke-tillåtande temperatur) och håll höga luftfuktighetsnivåer (minst 65 %) för att undvika larvuttorkning under analysen.
- Innan du påbörjar utfodringstestet ska du balansera analysplattornas temperatur genom att värma dem vid 30 °C i 30 minuter.
- (Valfritt) Värmechocka de experimentella larverna i 2 minuter i ett 37 ° C vattenbad. Utför detta steg med djuren i plastviktbåtarna som innehåller lite vatten.
OBS: Syftet med detta steg är att intensifiera den neuronala aktiveringen genom att främja avfyrningen av nervcellerna sedan början av utfodringsanalysen. - Håll flera timers inställda på 1 timme redo. Antalet timers som ska användas beror på antalet experimentella grupper som testas och på experimenterarens skicklighetsnivå vid hantering av larver.
OBS: Användning av flera timers är avgörande för att hålla analysens varaktighet konsekvent för alla genotyper. - Töm försiktigt vattnet från plastbåtarna och överför försiktigt L3-grupperna från båtarna till mitten av analysplattorna med hjälp av en fuktad mjuk borste. Sätt tillbaka plattornas lock och starta en timer för varje tallrik (eller grupp av plattor) för att upprätthålla exakta 1-h matningssessioner.
- Låt larverna mata i 1 timme, vid 30 °C, i mörker (figur 2C).
OBS: Prestandan hos analysen i mörkret är viktig för att kontrollera för skillnader i visuella signaler mellan dieter, eftersom dieterna kommer att skilja sig åt i toner, även om de innehåller samma färgkoncentration. - Stoppa matningsanalysen genom att överföra plattorna till ett isbad. Tryck ner isen så mycket som möjligt för att ge en stabil yta för plattorna.
OBS: Kalla temperaturer kommer att främja slutet av utfodringen genom att hämma grävnings- och grävbeteenden. De flesta larverna kommer att ytbehandla matplattorna efter några minuter, vilket underlättar deras återhämtning i följande steg.
5. Extraktion av livsmedelsfärger
- Förbered 2 ml mikrorör för varje grupp av 10 L3-testade, innehållande ungefär samma mängder 0,5 mm glaspärlor (tillräckligt för att fylla den nedre delen av mikroröret) och 300 μL iskall metanol. Håll mikrorören i kylan med en bänkkylare.
VARNING: Metanol är mycket brandfarligt och giftigt. Följ alla säkerhetsrutiner som rekommenderas för hantering av detta reagens, inklusive arbete i ett välventilerat område och bär nitrilhandskar.
OBS: Användning av metanol är viktigt för att fixa larvproverna och undvika melaniseringsreaktioner i nagelbandet. - Använd #5 eller fjäderviktstångor, återställ försiktigt grupperna på 10 L3 från matningsanalysplattorna och överför dem till locken på analysplattorna som innehåller lite vatten. Skölj larverna för att ta bort matrester på deras kroppar medan du försiktigt hanterar larverna för att undvika skador. För register över antalet larver som återvinns för varje genotyp per replikat, så att den genomsnittliga mängden matintag per larva kan kvantifieras.
OBS: Skadade larver bör kasseras eftersom de kommer att ha melaniserad nagelband, vilket är olämpligt för kolorimetrisk kvantifiering. - Överför L3-grupperna till de 2 ml mikrorör som är beredda i 5,1.
- Lysa larvvävnaderna för att extrahera matfärgen från tarmarna med en mekanisk lysmetod med hjälp av en vävnadslyzer och glaspärlor som tillsätts i steg 5.1. (om en vävnadslyzer inte är tillgänglig, använd en homogeniserande mortel). Utför företrädesvis detta steg vid 4 °C(figur 2D).
OBS: Hur länge detta steg är beror på vilken utrustning som används. Med hjälp av en konventionell vävnadslyst räcker 1 min extraktion. Vid tidsförträngningar kan protokollet pausas i slutet av det här steget och fortsätta senare. Förvara proverna vid -20 °C. - Överför extrakten för att rengöra 1,5 ml mikrorör genom att direkt invertera de 2 ml mikrorören till de nya 1,5 ml mikrorören. Om de utförs försiktigt kommer de flesta glaspärlor att stanna längst ner på 2 ml mikrorör.
- Rensa cellavfallet genom att centrifugera extrakten, med en maximal hastighet i 10 min, vid 4 °C.
- Samla supernatanter för att rengöra 1,5 ml mikrorör. Om cellulärt skräp fortfarande är synligt i supernatanterna, upprepa steg 5.6 och 5.7.
6. Kolorimetrisk kvantifiering av livsmedelskonsumtionen
- Förbered standardlösningar för att generera en kalibreringskurva genom att utföra seriella 1:2 utspädningar i metanol av en startblå färglösning. Som blind, använd endast metanol. Koncentrationen av normerna är beroende av djurens nivåer av födointag.
OBS: När det gäller pilotskärmen som presenteras här, eftersom färgkoncentrationerna som erhållits för larvextrakten varierade från 0,02 till 1,93 μL/ml, användes en standardkurva som erhållits genom att mäta absorbanserna av 8 seriella utspädningar av en 2 μL/ml blåfärgslösning i metanol. Vid behov öka eller minska koncentrationen av dessa lösningar, beroende på färgkoncentrationen av försöksproverna. - Överför 100 μL av försöksproverna (erhållna i steg 5.7), standarder och blinda (steg 6.1) till brunnarna i en 96-brunns mikroplatta och mät absorbansen vid 600 nm, med hjälp av en plåtläsare (figur 2E). För att ta bort bakgrundsabsorbansen, mät absorbansen av extrakt som erhålls från larver som matas på mat utan blått färgämne som en "noll" för larvextrakten ("nollfärgningsmedelskontroll).
- Generera en standardkurva och korrelera de absorbansvärden som erhålls för proverna från varje experimentell larvgrupp med mängden matintag (volym i ml). Hitta den genomsnittliga matkonsumtionen per larva genom att ta hänsyn till antalet larver som samlats in för varje grupp i steg 5.2
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Drosophila larver reglerar sitt proteinintag till priset av att inta överskott avkolhydrater 23 (schematiskt område i figur 2E). Egentligen har denna prioritering av proteinintag observerats hos många andra djur och kallas proteinet som utnyttjar24,25.
Dra nytta av detta robusta utfodring beteendemässiga svar, en beteendebaserad skärm utformades syftar till att identifiera neuronala populationer som är involverade i makronäringsämnen balansering. En no-choice utfodringsanalys fastställdes, som bestod i att tillåta grupper av L3 (10 individer per grupp) att mata ad libitum i 1 timme och under neuronala termogenetiska aktiveringsförhållanden med hjälp av dTRPA1, i tre isokaloriska (248 Cal/L) livsmedelsfärgade dieter som innehåller specifika P:C-förhållanden (1:1, 1:4 och 1:16) (figur 1 och figur 2C). Som en avläsning användes den genomsnittliga mängden mat som äts i makronäringsämnena dieter av olika P:C-förhållanden. Genom att dra nytta av Gal4/ UAS-systemet21 och använda några av Janelia Gal4-linjerna från FlyLight Project18,19, inducerades uttrycket av dTRPA1 i specifika neuronala populationer.
Med de metoder som beskrivs i detta protokoll kunde vi kvantifiera den relativa mängden makronäringsämnen som konsumeras, när det gäller P:C-förhållanden, för djur under termogenetisk aktivering av specifika neuronala populationer i larv nervsystemet. Detta experimentella tillvägagångssätt visade att aktivering av distinkta populationer av nervceller påverkade makronäringsämnesbalanseringen avsevärt i tredje-instar larver(figur 4, tabell 1). Det utfodringsmönster som observerats för kontrolllinjen (attP2) visar metodens effektivitet genom att visa en förväntad kompensatorisk ökning av födointaget av larver som testats i dieter med lägre P:C-förhållande (grå prickar och linje i figur 4). Dessutom hittades en betydande interaktion mellan genotyperna och kosten, vilket innebär att den termogenetiska aktiveringen av specifika neuronala populationer förändrar hur larver reglerar sitt matintag som svar på kostens makronäringsämnena.
Matningsmönstren för de genotyper som testats i de tre makronäringsämnena balansering dieter (1:1, 1:4, 1:16) visas av de färgade punkterna och linjerna i figur 4 och den statistiska analysen finns i tabell 1.
På aktiveringsskärmen testades totalt 36 Janelia Gal4-linjer som är kända för att vara glest uttryckta i larvnervsystemet. Med linjär regressionsmodeller bestämde vi vilka genotyper som uppvisade betydligt annorlunda matintag med hänvisning till de genetiska kontrolldjuren. Dessa skillnader inkluderade antingen skillnader i den absoluta mängden mat som äts över alla dieter, eller skillnader i makronäringsämnets balanseringsrespons (lutning av svaret på dietens olika P:C-förhållanden).
I alla tre dieterna åt R12E06 betydligt mer mat än kontrolldjur. Dessutom överkompenserade det ökningen av livsmedelsintaget på mellanliggande och lågproteindieter, vilket framgår av en betydande skillnad i interaktionstermen mellan matintag och P:C-förhållandet mellan kosten (tabell 1). R22H01 åt betydligt mer än kontroller men skilde sig inte åt i makronäringsbalanssvaret (tabell 1). Larver R14B11, R19G11, R21B06, R29C02 och R48F09 åt små mängder mat och förlorade förmågan att kompensera för den dåliga makronäringsämneskvaliteten i den tillgängliga kosten (vilket framgår av de betydande interaktionsvillkoren mellan matintag och P:C-förhållandet i kosten, tabell 1). Slutligen åt R45D11 larver betydligt mer i den proteinrika kosten som innehåller ett P:C-förhållande på 1:1 än i mellanliggande och i de proteinfattiga dieterna (1:4 och 1:16), vilket är motsatsen till vad man kan förvänta sig på lågproteindieterna.
Därför tillät våra metoder oss att klassificera de experimentella larverna, från varje genotyp, till fenotypiska klasser relaterade till den totala mängden mat som äts och förmåga att prioritera proteinintag genom att överkonsumera i dieterna för lågt P:C-förhållande. Fem fenotypiska klasser inrättades för försöksdjuren (figur 5): 1 – "Ät mycket" (mer än kontrolldjuren) och överkompensera för proteinutspädning. 2 – "Ät mycket men kompensera normalt"; 3 – "Ät lite (mindre än kontrollen) men kompensera"; 4 – "Ät lite och kompensera inte"; 5 – "Ät avvikande" (mer i proteinrika och mellanliggande dieter än i den proteinfattiga kosten). Dessutom, för var och en av dessa fenotypiska klasser och genotyper, visar vi GFP-mönstren i centrala nervsystemet hos tredje-instar larver. Denna information erhölls från offentligt tillgängliga bilddata i FlyLight Project onlineplattform, där man kan få tillgång till uttrycksmönstren för alla RubinGal4-intresselinjer 26.

Figur 1: Sackarosjäst-jäst (SY) dieter som används i vårt protokoll. (A) De blå punkterna representerar de isokaloiska (248 kalorier/L) makronäringsämnen balanserande dieter som används i utfodringsanalysen, som skiljer sig åt i förhållandet protein till kolhydrater (P:C) : 1:1, 1:4 och 1:16. Den beige pricken representerar den diet som används för att föda upp de experimentella tredje-instar larverna (L3), som innehöll ett P:C-förhållande på 1:2 och en kaloritäthet på 495 kalorier/L. (B)Detaljerad sammansättning och näringsinformation om sackarosjästjäst (SY) baserade dieter. Komponenterna är desamma för alla dieter: agar, sackaros och jäst. Mängden i gram av de komponenter som behövs för att förbereda 1 L diet visas. Observera att 1% (v/v) av blått färgämne måste tillsättas till makronäringsämnena balansering dieter och till L3 uppfödning diet nipagin och propionsyra lösningar måste läggas till en slutlig koncentration (v/v) av 3% respektive 0,3%. Klicka här om du vill visa en större version av den här figuren.

Figur 2: Schematisk representation av de viktigaste stegen i vårt protokoll (A) Genetiska kors av föräldralinjer som utnyttjar Gal4 / UAS-systemet. Korsningen mellan Rubin Gal4-linjerna och UAS-linjen kodning dTRPA1 möjliggör termogenetisk aktivering av specifika neuronala populationer i larvens centrala nervsystem. B)Beredning av de experimentella tredje-instar larverna (L3). Föräldrarnas kvinnor tilläts lägga ägg i 3-4 h och larv iscensättning sker vid den tillåtande temperaturen (18 °C) i 9 dagar. Som tillval är värmestöten vid 37 °C i 2 minuter före utfodringstestet. C)Termogenetisk aktivering av neuronal funktion och valfri utfodringsanalys i 1 timme vid icke-tillåtande temperatur (30 °C). Tre grupper av 10 experimentella L3 från varje genotyp tilläts mata i var och en av de makronäringsämnen balansera dieter som innehåller specifika protein till kolhydrater (P:C) förhållanden (1:1, 1:4 och 1:16). D)Extraktion av livsmedelsfärger. Mekanisk lys av larver, med hjälp av en vävnadslyst, för att extrahera det blå matfärgämnet. E)Kvantifiering av livsmedelsintaget. Kolorimetrisk kvantifiering av den genomsnittliga mängden mat som äts per larva genom att kvantifiera färgämneskoncentrationen i larvextrakten. Absorbansen av experimentella prover, standarder och "noll" mättes vid 600 nm (blå), med hjälp av en 96-brunns plattläsare. Klicka här om du vill visa en större version av den här figuren.

Figur 3: Skillnader mellan andra (L2) och tredje-instars Drosophila larver (L3). L2 och L3 kan lätt särskiljas genom observation av spiracles under ett stereomikroskop. L2:s främre spiracles är klubbliknande, medan de i L3 är förgrenade. Andra egenskaper kan hjälpa till att skilja de två instars men är subjektiva och mindre tillförlitliga. De bakre spiraclesna av L3 har en mörk orange ring på spetsen, som saknas eller är svagt närvarande i L2. Luftstrupen är tjockare i L3 larver. Illustration av Marisa Oliveira. Klicka här om du vill visa en större version av den här figuren.

Figur 4: Mängden mat som äts per larva under neuronala termogenetiska aktiveringsförhållanden i tre makronäringsämnen som innehåller specifika förhållandet mellan protein och kolhydrater (P:C). Genomsnittliga nivåer av mängden mat som äts per larva (mL) i 3 makronäringsämnen balanserande dieter som innehåller de specifika P:C-förhållandet 1:1, 1:4 och 1:16. Grupper av 10 tredje-instar larver, från varje genotyp, tilläts att mata under 1 timme, under neuronal termogenetisk aktivering villkor, med hjälp av dTRPA1, vid 30 °C. De genotyper som testats (larv avkomma från de genetiska korsningarna mellan Rubin Gal4-linjerna och UAS dTRPA1-linjen) indikeras av prickar och linjer av olika färger. Som en genetisk kontroll (anges i grått) användes larv avkomman från en korsning mellan linjen "tom Gal4" (attP2) och UAS dTRPA1. Namnen som gavs till genotyperna, som anges i förklaringen, var relaterade till raderna "Rubin GAL4" som användes. Klicka här om du vill visa en större version av den här figuren.

Figur 5: Gruppering av de linjer som testats i 5 huvud fenotypiska klasser. De fenotypiska klasser som anges i siffror baserades på kombinationen av fenotyper som observerats i form av total mängd mat som ätits och förmågan att upprätthålla proteinintagsprioriteringssvaret: 1 - ät mycket (mer än kontrolldjuren) och kunde kompensera för proteinutspädning genom överätning; 2 – ät mycket och kunde inte kompensera; 3 - ät lite (mindre än kontrollen) men kompensera; 4 – ät lite och kunde inte kompensera; och 5 - en extra fenotypisk klass, som kallades "avvikande", där larverna inte betedde sig som förväntat som svar på makronäringsämnets utspädning av proteininnehållet i kosten och åt mer i proteinrika och mellanliggande dieter än i den proteinfattiga kosten. För varje genotyp visas GFP-uttrycksmönstret i de centrala nervsystemet hos tredje-instar larver. Dessa bilddata från Rubin Gal4-linjerna som används i denna analys extraherades från den offentligt tillgängliga FlyLight Projectonlineplattformen 26. Klicka här om du vill visa en större version av den här figuren.
| Anova-tabell (typ II-tester) | |||||
| Svar: Koncentration/L3 | |||||
| Summa kvm | Df | F-värde | Pr(>F) | ||
| mat | 0.086832 | 1 | 113.5358 | < 2,2e-16 | *** |
| genotyp | 0.078443 | 10 | 10.2567 | 9.762e-15 | *** |
| Föda : Genotyp | 0.064038 | 10 | 8.3733 | 6.416e-12 | *** |
| Rester | 0.215673 | 282 | |||
| Signifikanskoder: 0'**' 0.001 '**' 0.01 '*' 0.05'. 0.1 ' ' 1 | |||||
| Sammanfattningstabell (koefficienterna nedan jämförs med genotypen för attP-kontroll): | |||||
| uppskatta | Std. Fel | t värde | Pr(>|t|) | ||
| - Jag är inte så bra på att se till att | 0.064245 | 0.004316 | 14.886 | < 2e-16 | *** |
| mat | -0.058117 | 0.007206 | -8.066 | 2.10e-14 | *** |
| Genotyp R12E06 | 0.040243 | 0.008961 | 4.491 | 1.03e-05 | *** |
| Genotyp R14B11 | -0.053347 | 0.014361 | -3.715 | 0.000245 | *** |
| Genotyp R19G11 | -0.044880 | 0.010788 | -4.160 | 4.23e-05 | *** |
| Genotyp R21B06 | -0.051912 | 0.009363 | -5.544 | 6.79e-08 | *** |
| Genotyp R22H01 | 0.017682 | 0.007296 | 2.423 | 0.016004 | * |
| Genotyp R29C02 | -0.043102 | 0.011113 | -3.879 | 0.000131 | *** |
| Genotyp R40D06 | -0.005341 | 0.009876 | -0.541 | 0.589102 | |
| Genotyp R45C03 | 0.004064 | 0.009876 | 0.412 | 0.680997 | |
| Genotyp R45D11 | -0.052579 | 0.009876 | -5.324 | 2.08e-07 | *** |
| Genotyp R48F09 | -0.044612 | 0.011362 | -3.926 | 0.000108 | *** |
| Föda : Genotyp R12E06 | -0.037763 | 0.015440 | -2.446 | 0.015067 | * |
| Föda : Genotyp R14B11 | 0.058054 | 0.027100 | 2.142 | 0.033031 | * |
| Livsmedel : Genotyp R19G11 | 0.051532 | 0.017726 | 2.907 | 0.003937 | ** |
| Föda : Genotyp R21B06 | 0.054403 | 0.015689 | 3.467 | 0.000607 | *** |
| Föda : Genotyp R22H01 | -0.020863 | 0.012377 | -1.686 | 0.092979 | . |
| Föda : Genotyp R29C02 | 0.048996 | 0.018714 | 2.618 | 0.009317 | ** |
| Föda : Genotyp R40D06 | 0.003804 | 0.016550 | 0.230 | 0.818371 | |
| Föda : Genotyp R45C03 | 0.034117 | 0.016550 | 2.061 | 0.040177 | * |
| Föda : Genotyp R45D11 | 0.090661 | 0.016550 | 5.478 | 9.53e-08 | *** |
| Föda : Genotyp R48F09 | 0.051184 | 0.019045 | 2.688 | 0.007625 | ** |
| Signifikanskoder: 0'**' 0.001 '**' 0.01 '*' 0.05'. 0.1 ' ' 1 | |||||
| Kvarvarande standardfel: 0,02765 på 282 frihetsgrader | |||||
| Multipel R-kvadrat: 0,516, Justerad R-kvadrat: 0,4799 | |||||
| F-statistik: 14,31 på 21 och 282 DF, p-värde: < 2,2e-16 | |||||
Tabell 1: ANOVA-tabell för effekten av neuronal termogenetisk aktivering och makronäringsämnets kvalitet på den tillgängliga kosten på mängden matintag. En linjär modell monterades för att fastställa de genotyper som uppvisar ett utfodringsbeteende som skiljer sig väsentligt från kontrolldjuren.
| genotyp | Associerad gen | ursprung | BDSC-lagernummer |
| w[*] . P{UAS-TrpA1(B). K}attP2 / TM6B, Tb[1] | Bloomington | 26264 | |
| w[1118] ; P{GAL4.1Uw}attP2 | Janelia (olika) | 68384 | |
| w[1118] ; P{GMR12E06-GAL4}attP2 | netto (CG11450) | Janelia (olika) | Na |
| w[1118] ; P{GMR14B11-GAL4}attP2 / TM3, Sb[1] | dnc (CG32498) | Janelia (olika) | 49255 |
| w[1118] ; P{GMR19G11-GAL4}attP2 | CG33696 | Janelia (olika) | 48864 |
| w[1118] ; P{GMR21B06-GAL4}attp2 | oa2 (CG6919) | Janelia (olika) | 49857 |
| w[1118] ; P{GMR22H01-GAL4}attP2 | fru (CG14307) | Janelia (olika) | 49001 |
| w[1118] ; P{GMR29C02-GAL4}attP2 | Ptp69D (CG10975) | Janelia (olika) | 48088 |
| w[1118] ; P{GMR40D06-GAL4}attP2 | cnc (CG17894) | Janelia (olika) | 48616 |
| w[1118] ; P{GMR45C03-GAL4}attP2 | kni (CG4717) | Janelia (olika) | 47936 |
| w[1118] ; P{GMR45D11-GAL4}attP2 | pnt (CG17077) | Janelia (olika) | 49563 |
| w[1118] ; P{GMR48F09-GAL4}attP2 | dpr8 (CG32600) | Janelia (olika) | 50377 |
Tabell 2: Drosophila linjer som används i detta arbete. Detaljerad information om alla rader som används: kodnamn, genotyp, associerad gen, ursprung och Bloomington Drosophila Stock Center (BDSC) nummer.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Med detta protokoll kan man testa larvernas förmåga under termogenetisk aktivering av specifika neuronala populationer att reglera intagsnivåerna av protein och kolhydrater, två stora makronäringsämnen, när de utsätts för dieter med olika P:C-sammansättning. Denna metod testades i samband med en larv preliminär screening syftar till att identifiera neuronal populationer är associerade med kontroll av matintag över dieter av olika makronäringsämne kvalitet. Detta arbete bidrar också till att visa att Drosophila larver är värdefulla djurmodeller för att undersöka den neuronala grunden för utfodring beteenden i samband med näringsmässiga homeostas.
Förutom den information som lämnas som anteckningar i protokollet vill vi diskutera några viktiga aspekter ytterligare. Som i alla beteendemässiga analyser måste åtgärder vidtas av experimenteraren för att minimera variationen i samband med djurs beteende. En mycket viktig aspekt som man bör tänka på är relaterad till vikten av att få utvecklingssynkronerade djur. Användningen av tidiga L3 larver som är väl synkroniserade i deras utvecklingsstadium, kommer att minska de beteendevariationer som djuren uppvisar under utfodringsanalysen27. Synkronisering av larver uppnås genom korta äggläggningar och genom att kontrollera larvernas densitet i kulturerna. Använd inte längre perioder av äggläggning än de vi anger i protokollet (3-4 timmar). Att kontrollera larvtätheten till högst 200 djur per tallrik kommer också att undvika utvecklingsfördröjningar och eliminera ytterligare variation i utfodringsbeteendet. Observera att det första ägget ligger efter parning, måste kasseras för att upprätthålla homogenitet och få bättre synkroniserad larvutveckling. Kvinnor har befruktade ägg i äggledaren och lägger dem i olika utvecklingsstadier vilket gör det svårt att upprätthålla enhetlighet bland larvsamlingen. Det är absolut nödvändigt att åtminstone den första timmens äggsamlingsplatta kasseras före den slutliga samlingen. Tänk på att den stress som induceras till djuren under larvhantering också kan påverka beteendet negativt. Försök att vara så skonsam som möjligt genom att använda en mjuk och vattenfuktad borste. Slutligen bör du tänka på att ett stort antal replikat genererar en mer tillförlitlig datauppsättning.
Som i alla experimentella protokoll presenterar våra metoder vissa begränsningar. Att använda en kolorimetrisk metod för att kvantifiera matintag baserat på ackumulering av ett livsmedelsfärgämne i djurens tarmar innebär vissa försiktighetsåtgärder i samband med analysens varaktighet. För vuxna flugor visades det att det finns en betydande risk för att nå ett stabilt tillstånd för färgansamling, där hastigheten på egestion är lika med intagshastigheten, vilket minskar noggrannheten hos metoden28. Även om det inte finns några bevis för att detta händer i larver, bestämde vi oss för att utföra en utfodringsanalys med en maximal varaktighet på 60 minuter. Denna varaktighet är bekväm och kompatibel med hög genomströmningsskärm. Att hålla protokollets totala varaktighet så kort som möjligt gör det också möjligt att slutföra alla steg i avsnitten 4, 5 och 6 på en arbetsdag. Om det är nödvändigt att ändra utfodringstestets varaktighet bör analystiderna på mellan 60 och 120 minuter möjliggöra en effektiv kvantifiering av livsmedelsintaget mellan genotyper, vilket tidigare visats29. Känsligheten hos matdöende metoder är också relativt låg när små mängder mat konsumeras, vilket avsevärt minskar upplösningen bland genotyper som uppvisar mycket låga nivåer av matintag. Vi satte upp våra foderanalyser med ett valfri paradigm. Endast en diettyp är tillgänglig för varje experimentell grupp larver, vilket inte tillåter djur att självständigt reglera nivåerna av proteiner och kolhydrater konsumtion. Dessutom, eftersom vi använder kemiskt odefinierade dieter, är det svårt att hålla kontroll över näringskoncentrationer som direkt kan påverka mönstren för larvmatning. För att övervinna dessa problem, eller för att bekräfta och ytterligare dissekera träffar som finns på en preliminär skärm, kanske experimenteraren vill överväga möjligheten att upprätta ett exakt och kontrollerat experimentellt näringssammanhang, genom att använda definierade syntetiska (holidiska) medier10 och ställa in livsmedelsvalsanalyser som tidigare beskrivits30. När du använder ett protokoll som involverar termogenetisk neuronal modulering, är det viktigt att tänka på att de nödvändiga temperaturförskjutningarna direkt kan påverka djurens beteendemässiga utgångar. En kompletterande användning av optogenetiska metoder skulle vara intressant att kontrollera för temperaturinducerade falska positiva, men användningen av optogenetik i samband med larvmatningsanalyser är tekniskt utmanande, eftersom utfodring av larver tillbringar större delen av tiden grävde i livsmedelssubstratet.
Ändå kan flera styrkor i vårt experimentella tillvägagångssätt räknas upp. Enkelheten och den relativt höga genomströmningen av vår metod möjliggör kvantifiering av matintag för flera genotyper när de utsätts för olika näringsförhållanden. Utfodringsbeteenden i larvstadiet är lättare kvantifierbara än hos vuxna flugor, vilket möjliggör generering av bättre funktionella avläsningar. Det är också mindre utmanande att fastställa utfodringsanalyser som liknar den naturliga miljön i larver än det är för vuxna, som det tidigare har diskuterats31. Dessutom, jämfört med andra tidigare etablerade metoder för att kvantifiera utfodring i larver, nämligen de som bygger på manuell räkning av antalet munkrok sammandragningar under en viss tidsperiod32, möjliggör vår kolorimetriska metod genetiska screeningstudier i större skalor. Vissa andra metoder är helt enkelt baserade på att poängsätta andelen larver med färgad mat i tarmarna, vilket inte tillåter en korrekt kvantifiering av matintagsnivåer33,34. När det gäller neurogenetisk kontroll av neuronal funktion, det faktum att TRPA1 transgene är inaktiv vid 18 °C säkerställer att neuronal aktivitet inte påverkas under hela larv utveckling. Detta säkerställer att den experimentella neuronala aktiveringen kommer att utföras uteslutande under utfodringsanalysen och inte under larvutvecklingen. Dessutom vill vi en gång till nämna att vårt protokoll lätt kan anpassas till experimentörens specifika behov och intressen. Till exempel kan undertryckandet av neuronalfunktionen, istället för aktivering, lätt erhållas genom att ersätta dTRPA1 för en UAS-linje som kodar den temperaturkänsliga neuronala ljuddämparen ShibireTS20. Om de utfodringsnivåer som uppvisas av de experimentella larverna är mycket låga, vilket gör det svårt att kvantifiera matintaget, är det möjligt att utföra ett extra steg på 30 min larv svält före utfodringstestet (före stegen i avsnitt 4 i protokollet), som tidigarebeskrivits 15. Detta matbriststeg kan vara särskilt intressant om du undersöker modulatorer av hungerdrivna beteenden. Slutligen, i tidigare studier, med kvantitativa kolorimetriska metoder, visades det att märkning av mat med blått färgämne inte har någon inverkan på utfodring12. Vi anser dock att användningen av kompletterande, mer exakta och känsliga metoder, som radiomärkning av maten12, i mer avancerade stadier av en studie, som syftar till att bekräfta eller ytterligare dissekera träffar som hittats under preliminära stadier skulle vara ett bra komplement till vår metod och bör övervägas av experimenteraren. Av alla dessa skäl tror vi på attraktionskraften hos våra metoder för att utföra genetiska skärmar (särskilt primära skärmar) som syftar till att identifiera neuronala populationer som är involverade i montering av neuronala kretsar som kodar utfodringsbeteenden.
Som en sista anmärkning vill vi nämna det faktum att tusentals larv Gal4-linjer etablerade i Janelia Research Campus är offentligt tillgängliga, på Bloomington Drosophila Stock Center och en stor mängd information om larv26 och vuxna19 CNS-uttrycksmönster är också offentligt tillgängliga i FlyLight Image Database (http://www.janelia.org/gal4-gen1). Dessa resurser gör det möjligt att utarbeta förmodade strukturfunktion neuronala kartor över de neuroner som reglerar utfodringsbeteendet hos Drosophila larver. Detta är möjligt genom att integrera fenotypisk information som genereras i neuronala skärmar med uttrycksmönstren hos de förare som används. Vi anser att våra metoder utgör ett giltigt tillvägagångssätt för att generera preliminära neuronala kartor för utfodring beteenden i samband med makronäringsämnen balansering i Drosophila hjärnan.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Författarna har inget att avslöja.
Acknowledgments
Vi vill tacka Instituto Gulbenkian de Ciência (regeringskonferensen) för att ha gett oss tillgång till en del av den experimentella utrustning som beskrivs i detta protokoll. Detta arbete stöddes av Portugisiska stiftelsen för vetenskap och teknik (FCT), LISBOA-01-0145-FEDER-007660, PTDC/NEU- NMC/2459/2014, IF/00697/2014 och La Caixa HR17-00595 till PMD och av Australian Research Council Future Fellowship (FT170100259) till CKM.
Materials
| Name | Company | Catalog Number | Comments |
| 1.5 mL microtubes | Sarstedt AG & Co. | 72.690.001 | |
| 10xPBS | Nytech | MB18201 | |
| 2.0 mL microtubes | Sarstedt AG & Co. | 72.695.500 | |
| 60 mm petri dishes | Greiner Bio-one, Austria | 628161 | |
| 96 well microplates | Santa Cruz Biotechnology | SC-204453 | |
| Agar | Pró-vida, Portugal | ||
| Bench cooler | Nalgene, USA | Labtop Cooler 5115-0032 | |
| Blue food dye | Rayner, Billingshurst, UK | ||
| Cell disruption media | Scientific Industries, Inc. | 888-850-6208 | (0.5 mm glass beads) |
| Dish weight boats | Santa Cruz Biotechnology | SC-201606 | |
| Embryo collection cage for 60 mm petri dishes | Flystuff, Scientific Laboratory Supplies, UK | FLY1212 (59-100) | |
| Featherweight forceps | BioQuip Products, USA | 4750 | |
| Fly food for stocks maintenance | 1 L food contains: 10 g Agar, 100 g Yeast Extract, 50 g Sucrose, 30 mL Nipagin, 3 mL propionic acid | ||
| Forceps #5 | Dumont | 0108-5-PS | Standard tips, INOX, 11cm |
| Incubator | LMS Ltd, UK | Series 2, Model 230 | For thermogenetic feeding assay (30?C) |
| Incubator | Percival Scientific, USA | DR36NL | To stage larvae (19?C) |
| Janelia lines | Janelia Research Campus | Detailed information in Table 2 | |
| Macronutrient balancing diets | Composition and nutritional information in Figure 1 | ||
| Methanol | VWR | CAS number: 67-56-1 | |
| Nipagin (Methyl 4-hydroxybenzoate) | Sigma-Aldrich | H5501 | |
| Nitrile gloves | VWR, USA | ||
| Refrigerated centrifuge | Eppendorf, Germany | 5804 R / Serial number: 5805CI364293 | |
| Rubin Gal4 ines | Janelia Research Campus | Stoks available at Bloomington Drosophila Stock Center | |
| ShibireTS UAS line | Bloomington Drosophila Stock Center | BDSC number: 66600 | Provided by Carlos Ribeiro Group |
| Soft brushes | For sorting anaesthetised fruit flies | ||
| Spectrophotometer plate reader | Thermo Fisher Scientific | Multiskan Go 51119300 | |
| Stereo microscope | Nikon | 1016625 | |
| Sucrose | Sidul, Portugal | ||
| Third-instar larvae (L3) rearing diet | Composition and nutritional information in Figure 1 | ||
| Timer | |||
| Tissue lyzer / bead beater | MP Biomedicals, USA | FastPrep-24 6004500 | |
| TRPA1 UAS line | Bloomington Drosophila Stock Center | BDSC number: 26264 | Expresses TrpA1 under UAS control; may be used to activate neurons experimentally at 25 ?C |
| Water bath | Sheldon Manufacturing Inc., USA | W20M-2 / 03068308 / 9021195 | |
| Yeast extract | Pró-vida, Portugal | 51% Protein, 15% Carbohydrate |
References
- Raubenheimer, D. Nature of nutrition - a unifying framework from animal adaptation to human. , (2012).
- Carvahlo, M. J. a, Mirth, C. K. Coordinating morphology with behavior during development: an integrative approach from a fly perspective. Frontiers in Ecology and Evolution. , (2015).
- Steck, K., et al. Internal amino acid state modulates yeast taste neurons to support protein homeostasis in Drosophila. Elife. 7, 31625 (2018).
- Itskov, P. M., Ribeiro, C. The dilemmas of the gourmet fly: the molecular and neuronal mechanisms of feeding and nutrient decision making in Drosophila. Frontiers in Neuroscience. 7, 12 (2013).
- Bjordal, M., Arquier, N., Kniazeff, J., Pin, J. P., Leopold, P. Sensing of amino acids in a dopaminergic circuitry promotes rejection of an incomplete diet in Drosophila. Cell. 156 (3), 510-521 (2014).
- Grandison, R. C., Piper, M. D., Partridge, L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature. 462 (7276), 1061-1064 (2009).
- Lee, K. P., et al. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proceedings of the National Academy of Sciences of the United States of America. 105 (7), 2498-2503 (2008).
- Levine, M. E., et al. Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population. Cell Metabolism. 19 (3), 407-417 (2014).
- Solon-Biet, S. M., et al. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metabolism. 19 (3), 418-430 (2014).
- Piper, M. D., et al. A holidic medium for Drosophila melanogaster. Nature Methods. 11 (1), 100-105 (2014).
- Jones, W. D. The expanding reach of the GAL4/UAS system into the behavioral neurobiology of Drosophila. BMB Reports. 42 (11), 705-712 (2009).
- Deshpande, S. A., et al. Quantifying Drosophila food intake: comparative analysis of current methodology. Nature Methods. 11 (5), 535-540 (2014).
- Neckameyer, W. S. A trophic role for serotonin in the development of a simple feeding circuit. Developmental Neuroscience. 32 (3), 217-237 (2010).
- Gasque, G., Conway, S., Huang, J., Rao, Y., Vosshall, L. B. Small molecule drug screening in Drosophila identifies the 5HT2A receptor as a feeding modulation target. Scientific Reports. 3, (2013).
- Schoofs, A., et al. Selection of motor programs for suppressing food intake and inducing locomotion in the Drosophila brain. PLoS Biology. 12 (6), 1001893 (2014).
- Pocas, G. M., Crosbie, A. E., Mirth, C. K. When does diet matter? The roles of larval and adult nutrition in regulating adult size traits in Drosophila melanogaster. Journal of Insect Physiology. , 104051 (2020).
- Hamada, F. N., et al. An internal thermal sensor controlling temperature preference in Drosophila. Nature. 454 (7201), 217-220 (2008).
- Pfeiffer, B. D., et al. Tools for neuroanatomy and neurogenetics in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 105 (28), 9715-9720 (2008).
- Jenett, A., et al. A GAL4-driver line resource for Drosophila neurobiology. Cell Reports. 2 (4), 991-1001 (2012).
- Kitamoto, T. Conditional modification of behavior in Drosophila by targeted expression of a temperature-sensitive shibire allele in defined neurons. Journal of Neurobiology. 47 (2), 81-92 (2001).
- Brand, A. H., Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118 (2), 401-415 (1993).
- Shirangi, T. R., Stern, D. L., Truman, J. W. Motor control of Drosophila courtship song. Cell Reports. 5 (3), 678-686 (2013).
- Mirth, C. M. J. Food intake and food choice are altered by the developmental transition at critical weight in Drosophila melanogaster. Animal Behaviour. 126, 195-208 (2017).
- Simpson, S. J., Raubenheimer, D.
- Raubenheimer, D., Simpson, S. J. Integrative models of nutrient balancing: application to insects and vertebrates. Nutrition Research Reviews. 10 (1), 151-179 (1997).
- Li, H. H., et al. A GAL4 driver resource for developmental and behavioral studies on the larval CNS of Drosophila. Cell Reports. 8 (3), 897-908 (2014).
- Bhatt, P. K., Neckameyer, W. S. Functional analysis of the larval feeding circuit in Drosophila. Journal of Visualized Experiments. (81), e51062 (2013).
- Wong, R., Piper, M. D. W., Blanc, E., Partridge, L. Pitfalls of measuring feeding rate in the fruit fly Drosophila melanogaster. Nature Methods. 5 (3), 214-215 (2008).
- Almeida-Carvalho, M. J., et al. The Ol1mpiad: concordance of behavioural faculties of stage 1 and stage 3 Drosophila larvae. Journal of Experimental Biology. 220, Pt 13 2452-2475 (2017).
- Rodrigues, M. A., et al. Drosophila melanogaster larvae make nutritional choices that minimize developmental time. Journal of Insect Physiology. 81, 69-80 (2015).
- Wong, R., Piper, M. D., Wertheim, B., Partridge, L.
- Wu, Q., et al. Developmental control of foraging and social behavior by the Drosophila neuropeptide Y-like system. Neuron. 39 (1), 147-161 (2003).
- Wu, Q., Zhang, Y., Xu, J., Shen, P. Regulation of hunger-driven behaviors by neural ribosomal S6 kinase in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 102 (37), 13289-13294 (2005).
- Lingo, P. R., Zhao, Z., Shen, P. Co-regulation of cold-resistant food acquisition by insulin- and neuropeptide Y-like systems in Drosophila melanogaster. Neuroscience. 148 (2), 371-374 (2007).
Tags
Neurovetenskap Utgåva 160 Drosophila larver termogenetisk neuronal skärm larvmatningsbeteende makronäringsämnen balansering protein kolhydrater matintagsanalys matfärgning kolorimetrisk kvantifieringErratum
Formal Correction: Erratum: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae
Posted by JoVE Editors on 10/06/2020.
Citeable Link.
An erratum was issued for: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae. A figure was updated.
Figure 1 was updated from:
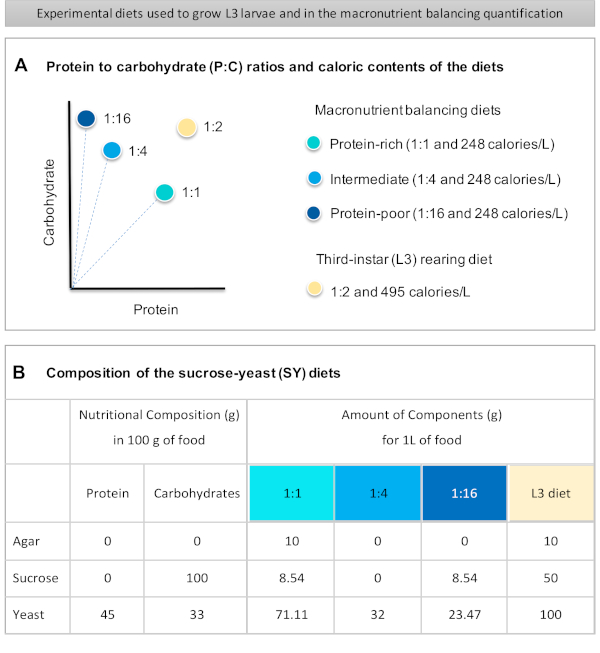
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
to:
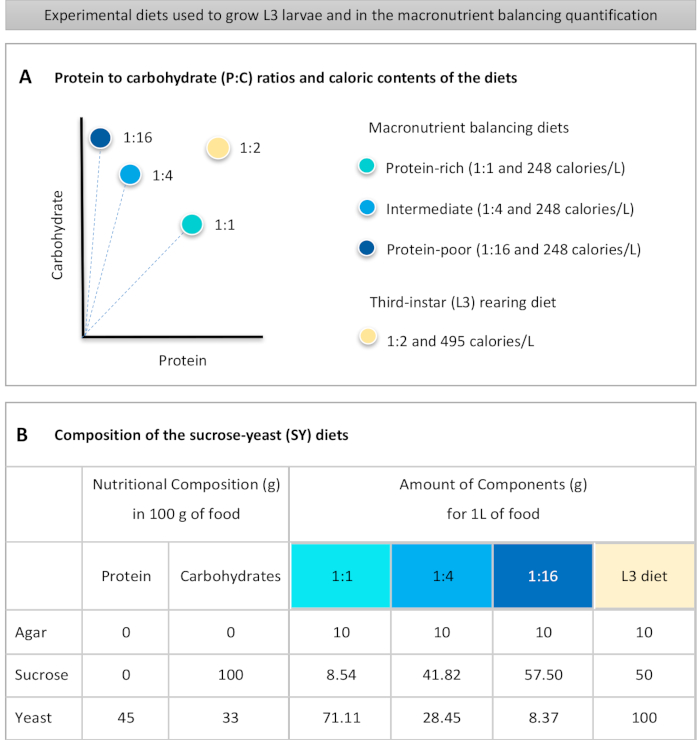
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
