

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Beskrevet her er en protokol, der muliggør den kolorimetriske kvantificering af mængden af mad, der spises inden for et defineret tidsinterval af Drosophila melanogaster larver udsat for kost af forskellig makronæringskvalitet. Disse assays udføres i forbindelse med en neuronal termogentisk skærm.
Abstract
Fouragering og fodring adfærd giver dyr adgang til energikilder og næringsstoffer afgørende for deres udvikling, sundhed og fitness. Undersøgelse af neuronal regulering af disse adfærdsmåder er afgørende for forståelsen af de fysiologiske og molekylære mekanismer, der ligger til grund for ernæringsmæssige homøostase. Brugen af genetisk tractable dyremodeller som orme, fluer og fisk letter i høj grad disse typer undersøgelser. I det sidste årti er frugtfluen Drosophila melanogaster blevet brugt som en kraftfuld dyremodel af neurobiologer, der undersøger den neuronale kontrol af fodring og fourageringsadfærd. Mens utvivlsomt værdifulde, undersøger de fleste undersøgelser voksne fluer. Her beskriver vi en protokol, der udnytter det enklere larvenervesystem til at undersøge neuronale substrater, der styrer fodringsadfærd, når larver udsættes for kost, der adskiller sig i deres protein- og kulhydratindhold. Vores metoder er baseret på en kvantitativ kolorimetrisk no-choice fodring assay, udført i forbindelse med en neuronal termogentisk-aktivering skærm. Som en aflæsning blev mængden af mad spist af larver over et 1 timers interval brugt, når den blev udsat for en af de tre farvemærkede kostvaner, der varierer i deres protein til kulhydrater (P: C) nøgletal. Effekten af denne protokol demonstreres i forbindelse med en neurogenetisk skærm i larve Drosophila, ved at identificere kandidat neuronale populationer, der regulerer mængden af mad, der spises i kostvaner af forskellig makronæringskvalitet. Vi var også i stand til at klassificere og gruppere genotyper testet i fænotypiske klasser. Ud over en kort gennemgang af de metoder, der i øjeblikket er tilgængelige i litteraturen, diskuteres fordelene og begrænsningerne ved disse metoder, og der gives også nogle forslag til, hvordan denne protokol kan tilpasses andre specifikke eksperimenter.
Introduction
Alle dyr er afhængige af en afbalanceret kost for at erhverve de nødvendige mængder næringsstoffer til overlevelse, vækst og reproduktion1. Valget af hvad og hvor meget at spise er påvirket af en lang række interagerende faktorer relateret til dyrets indre tilstand, som mæthedsniveauet og miljøforholdene, såsom fødevarekvalitet2,3,4,5. Protein og kulhydrater er to store makronæringsstoffer, og dets afbalancerede indtag er afgørende for at opretholde dyrenes fysiologiske processer. Derfor er forståelsen af de neurale mekanismer, der styrer fodringsadfærd og opretholder et afbalanceret indtag af disse makronæringsstoffer, yderst relevant. Dette skyldes, at livshistorieegenskaber som levetid, fecundity og metabolisk sundhed er direkte påvirket af niveauerne afproteinindtagsindtag 6,7,8,9,10.
Brugen af enklere mere tractable organismer, der udviser evolutionært bevarede fodringsvaner med komplekse dyr, herunder pattedyr, er afgørende for denne type undersøgelser. Det er vigtigt, at disse enklere dyremodeller giver en god mulighed for at dissekere komplekse biologiske spørgsmål i en dyr, etisk og teknisk mere effektiv sammenhæng. I de sidste årtier har Drosophila, med sin kraftfulde genetiske værktøjskasse, indviklet og stereotyp adfærd og bevaret arkitektur af perifere og næringsstoffølsomhedsmekanismer med pattedyr, været en frugtbar model for adfærdsmæssige neurobiologer11. I sidste ende er håbet, at ved at forstå, hvordan fødeindtagelse er reguleret i dette dyr, med et enklere nervesystem, kan vi derefter begynde at udrede neuronale funktionsfejl, der ligger til grund for menneskelige spiseforstyrrelser.
Undersøgelsen af neuronale substrater til fodring af adfærd er dybt afhængig af at kunne måle dyrs fødeindtagelse samtidig, mens de manipulerer deres neuronale aktivitet. På grund af de minimale mængder mad, der indtages, er det ekstremt udfordrende at kvantificere mængden af mad, der spises af fluer, og alle tilgængelige metoder udgør i øjeblikket betydelige begrænsninger. Guldstandarden er således at bruge en kombination af komplementære metoder12. Voksne fluer er historisk set blevet foretrukket som en genetisk og adfærdsmæssig model. Ikke desto mindre tilbyder Drosophila larver også muligheder for at undersøge neuronale substrater, der kodper fodringsadfærd. Larve centralnervesystemet (CNS), med omkring 12.000 neuroner, er betydeligt mindre komplekst end den voksne, som indeholder ca. 150.000 neuroner. Denne lavere kompleksitet er ikke kun numerisk, men også funktionel, da larveadfærd er afhængig af enklere lokomotivfunktioner og sensoriske systemer. På trods af den tilsyneladende enkelhed i deres nervesystemer udviser larver stadig fuldstændig fodringsadfærd, og nogle metoder til at kvantificere madindtagelse i Drosophila larver er blevet beskrevet5,13,14,15. Ved at parre sig med manipulationer af neuronal aktivitet kan Drosophila larver udgøre en meget tractable model til forståelse af den neurale regulering af fødeindtagelse.
Forudsat her er en detaljeret protokol til at kvantificere fødeindtagelse i larver udsat for kost af forskellig makronæringsstoffer kvalitet. Kostvanerne, såkaldte makronæringsstoffer balancering kostvaner, afveg i protein og kulhydrater indhold, specielt med hensyn til protein til kulhydrat (P: C) nøgletal: 1:1 (protein-rige kost), 1:4 (mellemliggende kost), og 1:16 (protein-dårlig kost), som vist i figur 1A. Kort, en kvantitativ no-choice fodring assay blev etableret ved hjælp af disse tre isocaloric saccharose-gær (SY)-baserede kostvaner farvet med en blå fødevare farvestof. Da gærekstrakt og saccharose blev anvendt som protein- og kulhydratkilder, og begge indeholder kulhydrater, blev variationen i P:C-forholdet opnået ved at ændre balancen mellem disse to komponenter, som tidligere beskrevet16 og som angivet i figur 1B. En skematisk oversigt over protokollen, der viser de vigtigste eksperimentelle trin, findes i figur 2.
Denne protokol blev oprettet med det formål at undersøge specifikke neuronale populationers rolle i reguleringen af larvefodringsniveauer i kostvaner med forskellige P:C-forhold og i forbindelse med en termogentisk neuronal skærm. Et velkarakterisk neurogenetisk værktøj blev brugt fra Transient Receptor Potential (TRP) familien: Drosophila Transient Receptor Potential kanal (dTRPA1), som er en temperatur og spænding-gated kation kanal, så fyring af handling potentialer, når omgivelsestemperaturerne stiger over 25 ° C17. For at udtrykke dTRPA1 transgene, vi benyttede os af Gal4 linjer baseret på cis-regulerenderegioner fra Drosophila genom, der er etableret i Rubin laboratorium, i forbindelse med FlyLight projektet på Janelia Research Campus18,19.
Selvom protokollen, her beskrevet, er blevet etableret i forbindelse med en aktiveringsskærm, kan den let tilpasses af eksperimentatoren til andre specifikke behov eller interesser, nemlig at udføre en undertrykkelsesskærm ved hjælp af den temperaturfølsomme neuronale lyddæmper ShibireTS20, alternativt til dTRPA1. Denne og andre tilpasninger behandles i protokol- og diskussionsafsnittene.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Tilberedning af saccharosegær (SY)
- Afveje alle de tørre ingredienser (agar, gær, saccharose) til makronæringsstoffer balancering og L3 opdræt kostvaner. Mængderne i gram for hver af de ingredienser, der er nødvendige for at tilberede 1 L fødevarer, er angivet i figur 1B.
BEMÆRK: Tag højde for, at der er behov for ca. 13 mL mad for at fylde en 60 mm petriskål. - Opløs alle ingredienser i sterilt destilleret vand (brug ca. 50% af den samlede mængde vand, der er nødvendig for at forberede fødevaren) og rør mediet i 5-10 minutter.
- Autoklave i 50 min.
- Efter at mediumerne er blevet afkølet, tilsættes nipagin- og propionsyreopløsninger til kostvanerne ved en endelig koncentration (v/v) på henholdsvis 3% og 0,3%. Til makronæringsstoffer balancering kostvaner, tilføje blå mad farvestof til en endelig koncentration (v / v) på 1%. Udfyld de samlede mængder med destilleret vand.
- Hæld forsigtigt maddiætterne til 60 mm Petri-retter, så mængden af hældt mad er omtrent den samme i hver af pladerne. Mærk pladerne med P:C-forholdet mellem kostvanerne.
BEMÆRK: Forbered makronæringsstoffer balancering kostvaner på dagen for fodring assay. Hvis det ikke er muligt, opbevares de tilberedte diæter ved 4 °C i en forseglet beholder i en maksimal varighed på 3 dage. Længere opbevaringsperioder gør kosten for tør og hård, og larverne kan ikke grave sig ind i mediet.
2. Genetisk krydsning af forældrelinjer
BEMÆRK: Brug Gal4/UAS-systemet21 til at opsætte de genetiske kryds. I denne protokol blev kvindelige jomfruer af UAS dTRPA1-linjen17 for at aktivere neuronal funktion i specifikke neuronale populationer brugt og krydset til mænd fra Janelia Gal4-linjerne (Figur 2A). Den anvendte genetiske kontrol var afkom af en krydsning mellem dTRPA1-linjen og en "tom GAL4"-linje, som bærer Gal4 i den vektor, der bruges til at generere Rubin Gal4-samlingen, men uden regulatorisk fragment til stede (attP2)22. For at fremme den neuronale undertrykkelse kan en UAS-linje, der kodper ShibireTS20, bruges i stedet for dTRPA1.
- Opsæt 60 mm embryonopsamlingsbure med L3-opdrætsdiætplader, suppleret med en aktiv gærpasta.
- De voksne UAS dTRPA1-hunromaner og Janelia Gal4-hanner i alderen 5-8 dage overføres til embryonindsamlingsburene, og parringen kan forekomme i 24-48 timer ved 25 °C med 60 % fugtighed og en 12:12 lysmør cyklus (Figur 2A). For 60 mm embryonopsamlingsbure skal du bruge omkring 100 jomfruhunner og 30 hanner pr. Kryds.
- I slutningen af parringsperioden skal du fjerne og kassere L3-opdrætsdiætpladerne, der anvendes i de genetiske kryds. Erstat dem med friske L3 opdræt kost plader, for at udføre ægget lægger og larve iscenesættelse.
3. Fremstilling af tredje-instar larver (L3)
- De parrede voksne fluer overføres til friske L3-opdrætsplader, og æglægningen kan forekomme i 3-4 timer ved 25 °C (figur 2B). Sørg for, at alle pladerne er mærket med genotypen, P:C-forholdet for kosten og datoen for ægget lå.
BEMÆRK: For at spare tid skal æglægningen udføres direkte i L3-opdrætsdiæten, så du undgår ekstra håndtering af æggene. I tilfælde af små genetiske screeninger kan optimering af æglægningen opnås ved hjælp af æblesaft agarplader. - I slutningen af æglægningsperioden skal du fjerne pladerne fra burene og dække dem med plastlåg. Hvis gærekstrakt bruges til at supplere L3 opdræt plader, sørg for at fjerne alle de resterende gær i slutningen af ægget lægning. Dette er vigtigt for at undgå ikke-ensartet fodring under larvevæksten.
BEMÆRK: De parrede voksne kan overføres til friske L3-opdrætsdiætplader, så der udføres flere ægskure, og der kan opnås flere eksperimentelle larver. På hinanden følgende æg-lægger kan udføres med de samme voksne i løbet af en hel arbejdsuge. - Antallet af æg pr. plade skal estimeres, og larvetætheden må højst holdes på 200 embryoner pr. plade. Dette skøn kan gøres ved at tælle antallet af embryoner i en fjerdedel af pladen.
BEMÆRK: En overfyldt plade vil forsinke larveudviklingen og påvirke larvefodringsadfærden. - L3-opdrætspladerne inkuberes ved 18 °C (eftergivende temperatur), 60% fugtighed og en 12:12 lysmør cyklus, og larverne kan vokse i 9 dage (Figur 2B).
- På den niende dag efter æglægning (AEL) indsamles tre grupper på 10 L3 fra hver af genotyperne (og for replikaterne), der skal testes. Derudover indsamle grupper af 10 L3 for "nul-farvestof mad" kontrol. Sørg for, at larverne opsamling sker i tilsvarende perioder af dagen, der anvendes til at gøre ægget lå (f.eks, hvis ægget om fandt sted mellem 10:00 til 14:00, indsamle larverne i samme periode 9 dage AEL) og udføres, så forsigtigt som muligt, ved hjælp af tang #5 eller en fjervægt pincet. Overfør larverne direkte som angivet i næste trin (3.6).
BEMÆRK: Kontroldyrene "nulfarvet mad" er larver, der i foderanalysen får mad uden blåfarvet farvestof. Denne kontrol er afgørende for at fjerne baggrundsabsorpionen af larveekstrakterne. - Overfør de indsamlede eksperimentelle larver til plastiske skålvægtbåde, der indeholder 1 mL vand. Sørg for, at L3 indsamles, og ikke L2, ved at følge anvisningerne i figur 3.
BEMÆRK: Indsamling af L3 til plastbåde, der indeholder vand eller 1x fosfatbufferet saltvand (PBS), er vigtigt for at holde larverne godt hydreret inden fodringsanalysens start. Dette er især vigtigt, hvis flere eksperimentelle L3-grupper fra forskellige genotyper indsamles på samme tid. Hold styr på indsamlingsrækkefølgen for hver gruppe, så forskelle i varigheden af madmangel for hver gruppe minimeres. Brugen af plastbåde i dette trin letter det valgfrie trin 4.3, da det gør det muligt for larver at flyde direkte ind i vandbadet.
4. Termogentisk aktivering og ikke-valg fodring assay
BEMÆRK: Det anbefales at udføre fodringsanalyserne på omtrent samme tidspunkt af dagen for at minimere mulige variationer relateret til døgnrytmen. Kør også altid kontroleksperimenter (afkom af den "tomme Gal4" linje krydset til UAS dTRPA1 og "nul-farvestof mad" larver), parallelt med genotyper af interesse.
- Opsæt en inkubator til 30 °C (ikke-eftergivende temperatur), og hold høje fugtighedsniveauer (mindst 65 %) for at undgå dehydrering af larve under analysen.
- Inden foderanalysen påbegyndes, skal analysepladernes temperatur udredes ved opvarmning ved 30 °C i 30 min.
- (Valgfrit) Varm-chok de eksperimentelle larver i 2 minutter i et 37 ° C vandbad. Udfør dette trin med dyrene i plastvægtbådene, der indeholder noget vand.
BEMÆRK: Formålet med dette trin er at intensivere den neuronale aktivering ved at fremme affyringen af neuronerne siden begyndelsen af fodringsanalysen. - Hold flere timere klar til 1 time. Antallet af timere, der skal anvendes, afhænger af antallet af forsøgsgrupper, der testes, og af eksperimentatorens færdighedsniveau ved håndtering af larver.
BEMÆRK: Brugen af flere timere er afgørende for at holde analysens varighed konsistent for alle genotyper. - Dræn forsigtigt vandet fra plastbådene, og overfør forsigtigt L3-grupperne fra bådene til midten af analysepladerne ved hjælp af en fugtet blød børste. Sæt pladernes låg tilbage og start en timer for hver plade (eller gruppe af plader) for at opretholde nøjagtige 1-timers fodringssessioner.
- Lad larverne fodre i 1 time ved 30 °C i mørke (Figur 2C).
BEMÆRK: Analysens ydeevne i mørke er vigtig for at kontrollere for forskelle i de visuelle signaler på tværs af kostvaner, da kosten vil variere i toner, selvom de indeholder den samme farvestofkoncentration. - Stop fodringsanalysen ved at overføre pladerne til et isbad. Tryk isen så meget som muligt ned for at give en stabil overflade til pladerne.
BEMÆRK: Kolde temperaturer vil fremme slutningen af fodringen ved at hæmme grave- og graveadfærd. De fleste af larverne vil overflade madpladerne efter nogle minutter, hvilket letter deres genopretning i følgende trin.
5. Udvinding af farvestoffer til levnedsmidler
- Der fremstilles 2 mL mikrorør til hver gruppe på 10 L3, der er testet, og som indeholder omtrent de samme mængder 0,5 mm glasperler (nok til at fylde den nederste del af mikrorøret) og 300 μL iskold methanol. Hold mikrorørene i kulden ved hjælp af en bænkkøler.
ADVARSEL: Methanol er meget brandfarlig og giftig. Følg alle de sikkerhedsprocedurer, der anbefales til håndtering af dette reagens, herunder arbejde i et godt ventileret område og iført nitrilhandsker.
BEMÆRK: Brugen af methanol er vigtig for at fikse larveprøverne og undgå melaniseringsreaktioner i neglebåndet. - Ved hjælp af #5 eller fjervægts-pincet genvindes forsigtigt grupperne på 10 L3 fra foderanalysepladerne og overføres til lågene på analysepladerne, der indeholder noget vand. Skyl larverne for at fjerne madrester på deres kroppe, mens de forsigtigt håndterer larverne for at undgå skader. Hold styr på antallet af larver, der er genvundet for hver genotype pr. replikat, således at den gennemsnitlige mængde fødeindtagelse pr. larve kan kvantificeres.
BEMÆRK: Skadede larver skal kasseres, da de vil have melaniseret neglebånd, der er uegnet til kolorimetrisk kvantificering. - L3-grupperne overføres til de 2 mL mikrorør, der er fremstillet i 5.1.
- Lys larvevævet for at udtrække madfarvet fra tarmene ved hjælp af en mekanisk lysismetode ved hjælp af en vævslyzer og glasperler, der tilsættes i trin 5.1. (hvis en vævslyzer ikke er tilgængelig, skal du bruge en homogeniserende støder). Fortrinsvis skal dette trin udføres ved 4 °C (figur 2D).
BEMÆRK: Varigheden af dette trin afhænger af det anvendte udstyr. Ved hjælp af en konventionel vævslyser er 1 min. ekstraktion tilstrækkelig. I tilfælde af tidssnævringer kan protokollen sættes på pause i slutningen af dette trin og fortsættes senere. Prøverne opbevares ved -20 °C. - Overfør ekstrakterne for at rengøre 1,5 mL mikrorør ved direkte at vende de 2 mL mikrorør på de nye 1,5 mL mikrorør. Hvis de udføres forsigtigt, forbliver de fleste glasperler i bunden af 2 mL mikrorøret.
- Cellulære vragrester ryddes ved at centrifugere ekstrakterne ved en maksimal hastighed i 10 min. ved 4 °C.
- Saml supernatanterne for at rengøre 1,5 mL mikrorør. Hvis cellulære vragrester stadig er synlige i supernatanterne, gentages trin 5.6 og 5.7.
6. Farvemetrisk kvantificering af fødevareforbruget
- Forbered standardløsninger til at generere en kalibreringskurve ved at udføre serielle 1:2 fortyndinger i methanol af en startblå farveopløsning. Som blank, brug kun methanol. Koncentrationen af normerne afhænger af dyrenes indhold af fødeindtagelse.
BEMÆRK: For den her præsenterede pilotskærm blev der anvendt en standardkurve opnået ved at måle absorbanserne af 8 serielle fortyndinger af en 2 μL/mL blåfarveopløsning i methanol, da farvestofkoncentrationerne for larveekstrakterne varierede fra 0,02 til 1,93 μL/mL. Hvis det er nødvendigt, øges eller mindskes koncentrationen af disse opløsninger afhængigt af farvekoncentrationen af forsøgsprøverne. - 100 μL af forsøgsprøverne overføres (opnået i trin 5.7), standarder og blindprøve (trin 6.1) til brøndene i en 96-brønds mikroplade, og absorbansen måles ved 600 nm ved hjælp af en pladelæser (Figur 2E). For at fjerne baggrundsabsorpmentet måles absorbansen af ekstrakter fra larver, der fodres med mad uden blåfarvet farvestof, som et "nul" for larveekstrakterne ("nulfarvet fødevarekontrol").
- Der genereres en standardkurve, og absorbansværdierne for prøverne fra hver forsøgs larvegruppe korreleres med mængden af fødeindtagelse (volumen i mL). Find det gennemsnitlige fødevareforbrug pr. larve ved at tage højde for antallet af larver, der er indsamlet for hver gruppe i trin 5.2
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Drosophila larver regulerer deres proteinindtag på bekostning af indtagelse af overskydende kulhydrater23 (skematisk plot i figur 2E). Faktisk er denne prioritering af proteinindtag blevet observeret hos mange andre dyr og kaldes proteinet, der udnytter24,25.
Ved at drage fordel af denne robuste fodring adfærdsmæssige reaktion, en adfærdsbaseret skærm blev designet til at identificere neuronale populationer involveret i makronæringsstoffer balancering. Der blev ikke etableret en foderanalyse uden valg. som bestod i at tillade grupper af L3 (10 personer pr. gruppe) at fodre ad libitum i 1 time og under neuronale termogentiske aktiveringsforhold ved hjælp af dTRPA1 i tre isokroniske (248 Cal/L) fødevarefarvede kostvaner, der indeholder specifikke P:C-nøgletal (1:1, 1:4 og 1:16)(figur 1 og figur 2C). Som en udlæsning blev den gennemsnitlige mængde mad, der blev spist i makronæringsstofferne i forskellige P:C-forhold, brugt. Drage fordel af Gal4/UAS system21 og ved hjælp af nogle af de Janelia Gal4 linjer fra FlyLight Project18,19, udtrykket af dTRPA1 blev induceret i specifikke neuronale populationer.
Med de metoder, der er beskrevet i denne protokol, var vi i stand til at kvantificere den relative mængde makronæringsstoffer, der forbruges, med hensyn til P: C-forhold, for dyr under termogentisk aktivering af specifikke neuronale populationer i larvenervesystemet. Denne eksperimentelle tilgang viste, at aktivering af forskellige populationer af neuroner i væsentlig grad påvirkede makronæringsstoffer i tredje-instar larver (Figur 4, Tabel 1). Det fodringsmønster, der er observeret for kontrollinjen (attP2), viser metodens effektivitet ved at vise en forventet kompenserende stigning i fødeindtagelsen hos larver, der er testet i diæter med lavere P:C-forhold (grå prikker og linje i figur 4). Desuden blev der fundet et betydeligt samspil mellem genotyperne og kosten, hvilket betyder, at den termogentiske aktivering af specifikke neuronale populationer ændrer den måde, larver regulerer deres fødeindtagelse på som reaktion på kostens makronæringsstofkvalitet.
Fodringsmønstrene for de genotyper, der testes i de tre makronæringsstoffers balanceringsdiæter (1:1, 1:4, 1:16), vises af de farvede prikker og linjer i figur 4, og den statistiske analyse er tilgængelig i tabel 1.
I aktiveringsskærmen blev i alt 36 Janelia Gal4-linjer, der vides at være sparsomt udtrykt i larvenervesystemet, testet. Ved hjælp af lineære regressionsmodeller bestemte vi, hvilke genotyper der udviste væsentligt forskellige fødeindtagelser med henvisning til de genetiske kontroldyr. Disse forskelle omfattede enten forskelle i den absolutte mængde mad, der spises på tværs af alle kostvaner, eller forskelle i makronæringsstoffer balancering svar (hældning af reaktionen på de forskellige P: C nøgletal af kostvaner).
På tværs af alle tre kostvaner spiste R12E06 betydeligt mere mad end kontroldyr. Desuden overkompenserede den stigningen i fødeindtagelsen på diæterne med mellemliggende og lavt proteinindhold, som det fremgår af en betydelig forskel i samspillet mellem fødeindtagelse og P:C-forholdet i kosten(tabel 1). R22H01 spiste betydeligt mere end kontrollerne, men var ikke forskellig i det makronæringsstofbalancerespons (tabel 1). R14B11, R19G11, R21B06, R29C02 og R48F09 larver spiste små mængder mad og mistede evnen til at kompensere for den dårlige makronæringsstoffer kvalitet af den tilgængelige kost (som angivet ved de betydelige interaktionsbetingelser mellem fødeindtagelse og P:C-forholdet i kosten, tabel 1). Endelig spiste R45D11 larver betydeligt mere i den proteinrige kost, der indeholder et P:C-forhold på 1:1 end i mellemprodukterne og i de proteinfattige kostvaner (1:4 og 1:16), hvilket er det modsatte af, hvad man kunne forvente på de lave proteindietter.
Derfor tillod vores metoder os at klassificere de eksperimentelle larver, fra hver genotype, i fænotypiske klasser relateret til den samlede mængde mad, der blev spist og evnen til at prioritere proteinindtag ved at overberegne i kosten af lavt P: C-forhold. Der blev fastsat fem fænotypiske klasser for forsøgsdyrene (figur 5): 1 – "Spis meget" (mere end kontroldyrene) og overkompenserer for proteinfortynding; 2 - "Spis en masse, men kompensere normalt"; 3 – "Spis lidt (mindre end kontrollen), men kompenser"; 4 - "Spis lidt og ikke kompensere"; 5 – "Spis afvigende" (mere i proteinrige og mellemliggende kostvaner end i proteinfattig kost). Derudover viser vi for hver af disse fænotypiske klasser og genotyper GFP-mønstrene i centralnervesystemet i tredje instar larver. Disse oplysninger blev indhentet fra de offentligt tilgængelige billeddata i FlyLight Project online platform, hvor man kan få adgang til udtrykket mønstre af alle Rubin Gal4 linjer af interesse26.

Figur 1: Den saccharosegær (SY) kost, der anvendes i vores protokol. (A) De blå prikker repræsenterer den isocaloric (248 kalorier / L) makronæringsstoffer balancering kostvaner, der anvendes i fodring assay, som varierer i protein til kulhydrat (P:C) nøgletal: 1:1, 1:4 og 1:16. Den beige prik repræsenterer den kost, der anvendes til at opdrætte de eksperimentelle tredje-instar larver (L3), som indeholdt en P:C-forholdet på 1:2 og en kalorietæthed på 495 kalorier / L. (B) Detaljeret sammensætning og ernæringsmæssige oplysninger om saccharose-gær (SY) baseret kost. Komponenterne er de samme for alle kostvaner: agar, saccharose og gær. Mængden i gram af de komponenter, der er nødvendige for at forberede 1 L kost, vises. Det skal bemærkes, at der skal tilsættes 1 % (v/v) blåt farvestof til makronæringsstoffernes balanceringsdiæt, og at L3-opdrætsdiet nipagin- og propionsyreopløsningerne skal tilsættes til en endelig koncentration (v/v) på henholdsvis 3% og 0,3%. Klik her for at se en større version af dette tal.

Figur 2: Skematisk repræsentation af de vigtigste trin, der er involveret i vores protokol (A) Genetisk krydsning af forældrelinjer, der drager fordel af Gal4/UAS-systemet. Krydsningen mellem Rubin Gal4-linjerne og UAS-linjen, der koder dTRPA1, tillader termogentisk aktivering af specifikke neuronale populationer i larve centralnervesystemet. B)Forberedelse af de eksperimentelle tredje instar larver (L3). Forældrenes hunner fik lov til at lægge æg i 3-4 timer, og larve iscenesættelsen sker ved den eftergivende temperatur (18 °C) i 9 dage. Valgfrit er varmechokket ved 37 °C i 2 minutter før fodringsanalysen. (C) Termogentisk aktivering af den neuronale funktion og foderanalyse uden valg i 1 time ved den ikke-eftergivende temperatur (30 °C). Tre grupper på 10 eksperimentelle L3 fra hver genotype fik lov til at fodre i hver enkelt af makronæringsstoffer balancering kostvaner, der indeholder specifikke protein til kulhydrater (P:C) nøgletal (1:1, 1:4 og 1:16). D) Udvinding af farvestoffer til levnedsmidler. Mekanisk lysis af larver, ved hjælp af en vævslyser, for at udtrække det blå madfarvestof. (E)Kvantificering af fødeindtagelse. Farvemetrisk kvantificering af den gennemsnitlige mængde mad, der spises pr. larve ved at kvantificere koncentrationen af levnedsmiddelfarvestoffer i larveekstrakterne. Absorbansen af forsøgsprøverne, standarderne og "nul" blev målt ved 600 nm (blå) ved hjælp af en 96-brønds pladelæser. Klik her for at se en større version af dette tal.

Figur 3: Forskelle mellem anden (L2) og tredje-instars Drosophila larver (L3). L2 og L3 kan let skelnes ved observation af spirakler under et stereomikroskop. Den forreste spirakler af L2 er klub-lignende, mens der i L3 er forgrenet. Andre egenskaber kan bidrage til at skelne mellem de to instars, men er subjektive og mindre pålidelige. Den bageste spirakler af L3 har en mørk orange ring på deres spids, som mangler eller svagt til stede i L2. Luftrøret er tykkere i L3 larver. Illustration af Marisa Oliveira. Klik her for at se en større version af dette tal.

Figur 4: Mængden af mad, der spises pr. larve under neuronale termogentiske aktiveringsforhold i tre makronæringsstoffer, der afbalancerer kostvaner, der indeholder specifikt protein til kulhydrater (P:C) nøgletal. Gennemsnitlige niveauer af mad spist pr larve (mL) i 3 makronæringsstoffer balancering kostvaner, der indeholder de specifikke P: C nøgletal på 1:1, 1:4 og 1:16. Grupper på 10 tredje-instar larver, fra hver genotype, fik lov til at fodre i løbet af 1 time, under neuronale termogentiske aktiveringsforhold, ved hjælp af dTRPA1, ved 30 °C. De testede genotyper (larveafkom fra de genetiske krydsninger mellem Rubin Gal4-linjerne og UAS dTRPA1-linjen) er angivet med prikker og linjer i forskellige farver. Som en genetisk kontrol (angivet med gråt) blev larvegenet fra en krydsning mellem den "tomme Gal4" linje (attP2) og UAS dTRPA1 anvendt. De navne, der blev givet til genotyperne, angivet i legenden, var relateret til de anvendte "Rubin GAL4"-linjer. Klik her for at se en større version af dette tal.

Figur 5: Gruppering af de testede linjer i 5 hovedphenotypiske klasser. De fænotypiske klasser, der er angivet med tal, var baseret på kombinationen af de fænotyper, der blev observeret med hensyn til den samlede mængde mad, der blev spist, og evnen til at opretholde proteinindtagsprioriteringsresponsen: 1 - spis meget (mere end kontroldyrene) og var i stand til at kompensere for proteinfortynding ved overspisning; 2 - spise en masse og var ikke i stand til at kompensere; 3 - spise lidt (mindre end kontrol), men kompensere; 4 – spise lidt og var ikke i stand til at kompensere; og 5 - en ekstra fænotypisk klasse, der blev kaldt "afvigende", hvor larverne ikke opførte sig som forventet som reaktion på makronæringsstoffer fortynding af proteinindholdet i kosten, spise mere i proteinrige og mellemliggende kostvaner end i proteinfattig kost. For hver genotype vises GFP-ekspressionsmønsteret i centralnervesystemet hos tredje-instar larver. Denne billeddata af Rubin Gal4 linjer, der anvendes i denne analyse blev udvundet fra den offentligt tilgængelige FlyLight Project online platform26. Klik her for at se en større version af dette tal.
| Anova tabel (type II-test) | |||||
| Reaktion: Koncentration/L3 | |||||
| Sum Sq | Df | F-værdi | Pr(>F) | ||
| mad | 0.086832 | 1 | 113.5358 | < 2.2e-16 | *** |
| genotype | 0.078443 | 10 | 10.2567 | 9.762e-15 | *** |
| Fødevarer : Genotype | 0.064038 | 10 | 8.3733 | 6.416e-12 | *** |
| Residualer | 0.215673 | 282 | |||
| Signifikanskoder: 0 '***' 0,001 '**' 0,01 '*' 0,05 '.' 0.1 ' ' 1 | |||||
| Sammenfattende tabel (nedenfor koefficienter sammenlignes med attP-kontrolgenotypen): | |||||
| estimat | Std. Fejl | t værdi | Pr(>|t|) | ||
| ( Skæring) | 0.064245 | 0.004316 | 14.886 | < 2e-16 | *** |
| mad | -0.058117 | 0.007206 | -8.066 | 2.10e-14 | *** |
| Genotype R12E06 | 0.040243 | 0.008961 | 4.491 | 1.03e-05 | *** |
| Genotype R14B11 | -0.053347 | 0.014361 | -3.715 | 0.000245 | *** |
| Genotype R19G11 | -0.044880 | 0.010788 | -4.160 | 4.23e-05 | *** |
| Genotype R21B06 | -0.051912 | 0.009363 | -5.544 | 6.79e-08 | *** |
| Genotype R22H01 | 0.017682 | 0.007296 | 2.423 | 0.016004 | * |
| Genotype R29C02 | -0.043102 | 0.011113 | -3.879 | 0.000131 | *** |
| Genotype R40D06 | -0.005341 | 0.009876 | -0.541 | 0.589102 | |
| Genotype R45C03 | 0.004064 | 0.009876 | 0.412 | 0.680997 | |
| Genotype R45D11 | -0.052579 | 0.009876 | -5.324 | 2.08e-07 | *** |
| Genotype R48F09 | -0.044612 | 0.011362 | -3.926 | 0.000108 | *** |
| Fødevarer : Genotype R12E06 | -0.037763 | 0.015440 | -2.446 | 0.015067 | * |
| Fødevarer : Genotype R14B11 | 0.058054 | 0.027100 | 2.142 | 0.033031 | * |
| Fødevarer : Genotype R19G11 | 0.051532 | 0.017726 | 2.907 | 0.003937 | ** |
| Fødevarer : Genotype R21B06 | 0.054403 | 0.015689 | 3.467 | 0.000607 | *** |
| Fødevarer : Genotype R22H01 | -0.020863 | 0.012377 | -1.686 | 0.092979 | . |
| Fødevarer : Genotype R29C02 | 0.048996 | 0.018714 | 2.618 | 0.009317 | ** |
| Fødevarer : Genotype R40D06 | 0.003804 | 0.016550 | 0.230 | 0.818371 | |
| Fødevarer : Genotype R45C03 | 0.034117 | 0.016550 | 2.061 | 0.040177 | * |
| Fødevarer : Genotype R45D11 | 0.090661 | 0.016550 | 5.478 | 9.53e-08 | *** |
| Fødevarer : Genotype R48F09 | 0.051184 | 0.019045 | 2.688 | 0.007625 | ** |
| Signifikanskoder: 0 '***' 0,001 '**' 0,01 '*' 0,05 '.' 0.1 ' ' 1 | |||||
| Resterende standardfejl: 0,02765 på 282 frihedsgrader | |||||
| Flere R-kvadrerede: 0,516, Justeret R-kvadreret: 0,4799 | |||||
| F-statistik: 14.31 på 21 og 282 DF, p-værdi: < 2.2e-16 | |||||
Tabel 1: ANOVA-tabel for effekten af neuronal termogentisk aktivering og makronæringsstoffer kvalitet af den tilgængelige kost på mængden af fødeindtagelse. Der blev monteret en lineær model til bestemmelse af de genotyper, der udviser en fodringsadfærd, der er væsentligt anderledes end kontroldyrene.
| genotype | Associeret gen | oprindelse | BDSC-aktienummer |
| w[*] ; P{UAS-TrpA1(B). K}attP2 / TM6B, Tb[1] | Bloomington | 26264 | |
| w[1118] ; P{GAL4.1Uw}attP2 | Janelia | 68384 | |
| w[1118] ; P{GMR12E06-GAL4}attP2 | netto (CG11450) | Janelia | Na |
| w[1118] ; P{GMR14B11-GAL4}attP2 / TM3, Sb[1] | DNC (CG32498) | Janelia | 49255 |
| w[1118] ; P{GMR19G11-GAL4}attP2 | CG33696 | Janelia | 48864 |
| w[1118] ; P{GMR21B06-GAL4}attP2 | oa2 (CG6919) | Janelia | 49857 |
| w[1118] ; P{GMR22H01-GAL4}attP2 | fru (CG14307) | Janelia | 49001 |
| w[1118] ; P{GMR29C02-GAL4}attP2 | Ptp69D (CG10975) | Janelia | 48088 |
| w[1118] ; P{GMR40D06-GAL4}attP2 | CNC (CG17894) | Janelia | 48616 |
| w[1118] ; P{GMR45C03-GAL4}attP2 | KNI (CG4717) | Janelia | 47936 |
| w[1118] ; P{GMR45D11-GAL4}attP2 | Pnt (CG17077) | Janelia | 49563 |
| w[1118] ; P{GMR48F09-GAL4}attP2 | dpr8 (CG32600) | Janelia | 50377 |
Tabel 2: Drosophila-linjer, der anvendes i dette arbejde. Detaljerede oplysninger om alle de anvendte linjer: kodenavn, genotype, tilknyttet gen, oprindelse og Bloomington Drosophila Stock Center (BDSC) nummer.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Med denne protokol kunne man teste larvernes evne under termogentisk aktivering af specifikke neuronale populationer til at regulere indtagelsesniveauerne for protein og kulhydrater, to store makronæringsstoffer, når de udsættes for kost af forskellig P:C-sammensætning. Denne metode blev testet i forbindelse med en larveforundersøgelse med det formål at identificere neuronale populationer forbundet med kontrol af fødeindtagelse på tværs af kostvaner af forskellig makronæringskvalitet. Dette arbejde bidrager også til at demonstrere, at Drosophila larver er værdifulde dyremodeller til at undersøge det neuronale grundlag for fodringsadfærd forbundet med næringsstofhomøostase.
Ud over de oplysninger, der gives som noter i protokollen, vil vi gerne yderligere drøfte nogle vigtige aspekter. Som i enhver adfærdsmæssig analyse skal der træffes foranstaltninger af eksperimentatoren for at minimere variationen forbundet med dyrs adfærd. Et meget vigtigt aspekt, som man skal huske på, er relateret til vigtigheden af at opnå udviklingssynkront synkroniserede dyr. Brugen af tidlige L3 larver, der er godt synkroniseret i deres udviklingsstadium, vil reducere de adfærdsmæssige variationer udstillet af dyrene under fodringsanalysen27. Synkronisering af larver opnås ved korte æglægninger og ved at kontrollere tætheden af larver i kulturerne. Brug ikke længere æglægningsperioder end dem, vi angiver i protokollen (3-4 timer). Også at kontrollere larvetætheden til et maksimalt antal på 200 dyr pr. Plade vil undgå udviklingsforsinkelser og eliminere yderligere variation i fodringsadfærd. Bemærk, at det første æg lå efter parring, skal kasseres for at opretholde homogenitet og opnå bedre synkroniseret larveudvikling. Hunnerne har befrugtede æg i oviduct og lægger dem i forskellige udviklingsstadier, hvilket gør det svært at opretholde ensartethed blandt larvesamlingen. Det er bydende nødvendigt, at mindst den første time æg indsamling plade kasseres før den endelige indsamling. Vær opmærksom på, at stress, der induceres til dyrene under larvehåndtering, også kan have en negativ indvirkning på adfærden. Prøv at være så blid som muligt ved hjælp af en blød og vand-fugtet børste. Endelig skal du huske, at et stort antal replikater genererer et mere pålideligt datasæt.
Som i enhver eksperimentel protokol, præsenterer vores metoder nogle begrænsninger. Ved hjælp af en kolorimetrisk metode til at kvantificere fødeindtagelse baseret på akkumulering af et levnedsmiddelfarvestof i dyrs tarme indebærer nogle forholdsregler i forbindelse med analysens varighed. For voksne fluer blev det påvist, at der er en betydelig risiko for at nå en stabil tilstand for farvestofakkumulering, hvor egestionshastigheden svarer til indtagshastigheden, hvilket reducerer nøjagtigheden af metoden28. Selvom der ikke er tegn på, at dette sker i larver, besluttede vi at udføre en fodringsanalyse med en maksimal varighed på 60 minutter. Denne varighed er praktisk og kompatibel med skærmen med høj overførselshastighed. Hvis protokollens samlede varighed er så kort som muligt, kan alle trin i afsnit 4, 5 og 6 også være afsluttet på én arbejdsdag. Hvis det er nødvendigt at ændre varigheden af foderanalysen, bør analysens varigheder fra 60 til 120 minutter muliggøre en effektiv kvantificering af fødeindtagelsen på tværs af genotyper, som tidligere påvist29. Følsomheden af fødevaredømte metoder er også relativt lav, når der forbruges små mængder mad, hvilket reducerer opløsningen blandt genotyper, der udviser meget lave niveauer af fødeindtagelse. Vi har oprettet vores fodring assays ved hjælp af en no-choice paradigme. Kun en kosttype er tilgængelig for hver eksperimentel gruppe larver, som ikke tillader dyr selvstændigt at regulere niveauet af proteiner og kulhydrater forbrug. Da vi desuden bruger kemisk udefinerede kostvaner, er det svært at holde styr på næringsstoffer koncentrationer, der direkte kan påvirke mønstrene for larvefodring. For at overvinde disse problemer, eller for at bekræfte og yderligere dissekere hits fundet på en foreløbig skærm, kan eksperimentatoren overveje muligheden for at etablere en præcis og kontrolleret eksperimentel ernæringsmæssig sammenhæng ved hjælp af definerede syntetiske (holidiske) medier10 og oprette fødevarevalgsanalyser som tidligere beskrevet30. Mens du bruger en protokol, der involverer termogentisk neuronal modulation, er det vigtigt at overveje, at de nødvendige temperaturændringer direkte kan påvirke dyrenes adfærdsmæssige output. En supplerende brug af optogenetiske tilgange ville være interessant at kontrollere for temperaturinducerede falske positiver, men brugen af optogenetics i forbindelse med larvefodringsanalyser er teknisk udfordrende, da fodring af larver bruger det meste af tiden gravet i fødevaresubstratet.
Ikke desto mindre kan flere styrker i vores eksperimentelle tilgang opregnes. Enkelheden og den relativt høje gennemløb af vores metode gør det muligt at kvantificere fødeindtagelsen for flere genotyper, når de udsættes for forskellige ernæringsmæssige forhold. Fodringsadfærd i larvestadiet er lettere kvantificerbare end i voksne fluer, hvilket gør det muligt at skabe bedre funktionelle udlæsninger. Det er også mindre udfordrende at etablere fodringsanalyser, der ligner det naturlige miljø i larver, end det er for voksne, som det tidligere er blevet diskuteret31. Sammenlignet med andre tidligere etablerede metoder til at kvantificere fodring i larver, nemlig dem, der er baseret på manuel optælling af antallet af mundkrogsammentrækninger i en vis periode32, muliggør vores kolorimetriske metode genetiske screeningsundersøgelser på større skalaer. Nogle andre metoder er simpelthen baseret på at score andelen af larver med farvet mad i deres tarme, hvilket ikke tillader en nøjagtig kvantificering af fødeindtagelsesniveauerne33,34. Med hensyn til neurogenetisk kontrol af neuronal funktion sikrer det faktum, at TRPA1 transgenet er inaktivt ved 18 °C, at neuronal aktivitet ikke påvirkes under larveudviklingen. Dette sikrer, at den eksperimentelle neuronale aktivering udelukkende udføres under fodringsanalysen og ikke under larveudviklingen. Derudover vil vi gerne endnu en gang nævne, at vores protokol let kan tilpasses eksperimentatorens specifikke behov og interesser. For eksempel kan undertrykkelsen af den neuronale funktion i stedet for aktivering let opnås ved at erstatte dTRPA1 med en UAS-linje, der koder den temperaturfølsomme neuronale lyddæmper ShibireTS20. Hvis fodringsniveauerne udstillet af de eksperimentelle larver er meget lave, hvilket gør det svært at kvantificere fødeindtagelsen, er det også muligt at udføre et ekstra trin på 30 min larve sult før fodringsanalysen (før trinene i protokollens afsnit 4), som tidligere beskrevet15. Denne mad-afsavn skridt kan være særligt interessant, hvis du undersøger modulatorer af sult-drevet adfærd. Endelig blev det i tidligere undersøgelser ved hjælp af kvantitative kolorimetriske metoder påvist, at mærkning af fødevarer med blåfarvet farvestof ikke har nogen indflydelse på fodring12. Ikke desto mindre mener vi, at brugen af komplementære, mere præcise og følsomme metoder, som radiomærkning af fødevarer12, i mere avancerede stadier af en undersøgelse, der har til formål at bekræfte eller yderligere dissekere hits fundet i de indledende faser ville være et godt supplement til vores metode og bør overvejes af eksperimentator. Af alle disse grunde tror vi på tiltrækningskraften af vores metoder til at udføre genetiske skærme (især primære skærme), der sigter mod at identificere neuronale populationer, der er involveret i samling af neuronale kredsløb, der kodper fodringsadfærd.
Som en sidste bemærkning vil vi gerne nævne det faktum, at tusindvis af larve Gal4 linjer etableret i Janelia Research Campus er offentligt tilgængelige, på Bloomington Drosophila Stock Center og en stor mængde information om larve26 og voksne19 CNS udtryk mønstre er også offentligt tilgængelige på FlyLight Image Database (http://www.janelia.org/gal4-gen1). Disse ressourcer gør det muligt at udarbejde formodede strukturfunktions neuronale kort over neuronerne, der regulerer fodringsadfærd i Drosophila larver. Dette er muligt ved at integrere de fænotypiske oplysninger, der genereres i neuronale skærme, med udtryksmønstrene for de anvendte drivere. Vi mener, at vores metoder udgør en gyldig tilgang til at generere foreløbige neuronale kort til fodringsadfærd forbundet med makronæringsstofbalance i Drosophila-hjernen.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har intet at afsløre.
Acknowledgments
Vi vil gerne takke Instituto Gulbenkian de Ciência (regeringskonferencen) for at give os adgang til en del af det eksperimentelle udstyr, der er beskrevet i denne protokol. Dette arbejde blev støttet af Det Portugisiske Institut for Videnskab og Teknologi (FCT), LISBOA-01-0145-FEDER-007660, PTDC/NEU- NMC/2459/2014, IF/00697/2014 og La Caixa HR17-00595 til PMD og af et australsk forskningsråds fremtidige stipendium (FT170100259) til CKM.
Materials
| Name | Company | Catalog Number | Comments |
| 1.5 mL microtubes | Sarstedt AG & Co. | 72.690.001 | |
| 10xPBS | Nytech | MB18201 | |
| 2.0 mL microtubes | Sarstedt AG & Co. | 72.695.500 | |
| 60 mm petri dishes | Greiner Bio-one, Austria | 628161 | |
| 96 well microplates | Santa Cruz Biotechnology | SC-204453 | |
| Agar | Pró-vida, Portugal | ||
| Bench cooler | Nalgene, USA | Labtop Cooler 5115-0032 | |
| Blue food dye | Rayner, Billingshurst, UK | ||
| Cell disruption media | Scientific Industries, Inc. | 888-850-6208 | (0.5 mm glass beads) |
| Dish weight boats | Santa Cruz Biotechnology | SC-201606 | |
| Embryo collection cage for 60 mm petri dishes | Flystuff, Scientific Laboratory Supplies, UK | FLY1212 (59-100) | |
| Featherweight forceps | BioQuip Products, USA | 4750 | |
| Fly food for stocks maintenance | 1 L food contains: 10 g Agar, 100 g Yeast Extract, 50 g Sucrose, 30 mL Nipagin, 3 mL propionic acid | ||
| Forceps #5 | Dumont | 0108-5-PS | Standard tips, INOX, 11cm |
| Incubator | LMS Ltd, UK | Series 2, Model 230 | For thermogenetic feeding assay (30?C) |
| Incubator | Percival Scientific, USA | DR36NL | To stage larvae (19?C) |
| Janelia lines | Janelia Research Campus | Detailed information in Table 2 | |
| Macronutrient balancing diets | Composition and nutritional information in Figure 1 | ||
| Methanol | VWR | CAS number: 67-56-1 | |
| Nipagin (Methyl 4-hydroxybenzoate) | Sigma-Aldrich | H5501 | |
| Nitrile gloves | VWR, USA | ||
| Refrigerated centrifuge | Eppendorf, Germany | 5804 R / Serial number: 5805CI364293 | |
| Rubin Gal4 ines | Janelia Research Campus | Stoks available at Bloomington Drosophila Stock Center | |
| ShibireTS UAS line | Bloomington Drosophila Stock Center | BDSC number: 66600 | Provided by Carlos Ribeiro Group |
| Soft brushes | For sorting anaesthetised fruit flies | ||
| Spectrophotometer plate reader | Thermo Fisher Scientific | Multiskan Go 51119300 | |
| Stereo microscope | Nikon | 1016625 | |
| Sucrose | Sidul, Portugal | ||
| Third-instar larvae (L3) rearing diet | Composition and nutritional information in Figure 1 | ||
| Timer | |||
| Tissue lyzer / bead beater | MP Biomedicals, USA | FastPrep-24 6004500 | |
| TRPA1 UAS line | Bloomington Drosophila Stock Center | BDSC number: 26264 | Expresses TrpA1 under UAS control; may be used to activate neurons experimentally at 25 ?C |
| Water bath | Sheldon Manufacturing Inc., USA | W20M-2 / 03068308 / 9021195 | |
| Yeast extract | Pró-vida, Portugal | 51% Protein, 15% Carbohydrate |
References
- Raubenheimer, D. Nature of nutrition - a unifying framework from animal adaptation to human. , (2012).
- Carvahlo, M. J. a, Mirth, C. K. Coordinating morphology with behavior during development: an integrative approach from a fly perspective. Frontiers in Ecology and Evolution. , (2015).
- Steck, K., et al. Internal amino acid state modulates yeast taste neurons to support protein homeostasis in Drosophila. Elife. 7, 31625 (2018).
- Itskov, P. M., Ribeiro, C. The dilemmas of the gourmet fly: the molecular and neuronal mechanisms of feeding and nutrient decision making in Drosophila. Frontiers in Neuroscience. 7, 12 (2013).
- Bjordal, M., Arquier, N., Kniazeff, J., Pin, J. P., Leopold, P. Sensing of amino acids in a dopaminergic circuitry promotes rejection of an incomplete diet in Drosophila. Cell. 156 (3), 510-521 (2014).
- Grandison, R. C., Piper, M. D., Partridge, L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature. 462 (7276), 1061-1064 (2009).
- Lee, K. P., et al. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proceedings of the National Academy of Sciences of the United States of America. 105 (7), 2498-2503 (2008).
- Levine, M. E., et al. Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population. Cell Metabolism. 19 (3), 407-417 (2014).
- Solon-Biet, S. M., et al. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metabolism. 19 (3), 418-430 (2014).
- Piper, M. D., et al. A holidic medium for Drosophila melanogaster. Nature Methods. 11 (1), 100-105 (2014).
- Jones, W. D. The expanding reach of the GAL4/UAS system into the behavioral neurobiology of Drosophila. BMB Reports. 42 (11), 705-712 (2009).
- Deshpande, S. A., et al. Quantifying Drosophila food intake: comparative analysis of current methodology. Nature Methods. 11 (5), 535-540 (2014).
- Neckameyer, W. S. A trophic role for serotonin in the development of a simple feeding circuit. Developmental Neuroscience. 32 (3), 217-237 (2010).
- Gasque, G., Conway, S., Huang, J., Rao, Y., Vosshall, L. B. Small molecule drug screening in Drosophila identifies the 5HT2A receptor as a feeding modulation target. Scientific Reports. 3, (2013).
- Schoofs, A., et al. Selection of motor programs for suppressing food intake and inducing locomotion in the Drosophila brain. PLoS Biology. 12 (6), 1001893 (2014).
- Pocas, G. M., Crosbie, A. E., Mirth, C. K. When does diet matter? The roles of larval and adult nutrition in regulating adult size traits in Drosophila melanogaster. Journal of Insect Physiology. , 104051 (2020).
- Hamada, F. N., et al. An internal thermal sensor controlling temperature preference in Drosophila. Nature. 454 (7201), 217-220 (2008).
- Pfeiffer, B. D., et al. Tools for neuroanatomy and neurogenetics in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 105 (28), 9715-9720 (2008).
- Jenett, A., et al. A GAL4-driver line resource for Drosophila neurobiology. Cell Reports. 2 (4), 991-1001 (2012).
- Kitamoto, T. Conditional modification of behavior in Drosophila by targeted expression of a temperature-sensitive shibire allele in defined neurons. Journal of Neurobiology. 47 (2), 81-92 (2001).
- Brand, A. H., Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118 (2), 401-415 (1993).
- Shirangi, T. R., Stern, D. L., Truman, J. W. Motor control of Drosophila courtship song. Cell Reports. 5 (3), 678-686 (2013).
- Mirth, C. M. J. Food intake and food choice are altered by the developmental transition at critical weight in Drosophila melanogaster. Animal Behaviour. 126, 195-208 (2017).
- Simpson, S. J., Raubenheimer, D.
- Raubenheimer, D., Simpson, S. J. Integrative models of nutrient balancing: application to insects and vertebrates. Nutrition Research Reviews. 10 (1), 151-179 (1997).
- Li, H. H., et al. A GAL4 driver resource for developmental and behavioral studies on the larval CNS of Drosophila. Cell Reports. 8 (3), 897-908 (2014).
- Bhatt, P. K., Neckameyer, W. S. Functional analysis of the larval feeding circuit in Drosophila. Journal of Visualized Experiments. (81), e51062 (2013).
- Wong, R., Piper, M. D. W., Blanc, E., Partridge, L. Pitfalls of measuring feeding rate in the fruit fly Drosophila melanogaster. Nature Methods. 5 (3), 214-215 (2008).
- Almeida-Carvalho, M. J., et al. The Ol1mpiad: concordance of behavioural faculties of stage 1 and stage 3 Drosophila larvae. Journal of Experimental Biology. 220, Pt 13 2452-2475 (2017).
- Rodrigues, M. A., et al. Drosophila melanogaster larvae make nutritional choices that minimize developmental time. Journal of Insect Physiology. 81, 69-80 (2015).
- Wong, R., Piper, M. D., Wertheim, B., Partridge, L.
- Wu, Q., et al. Developmental control of foraging and social behavior by the Drosophila neuropeptide Y-like system. Neuron. 39 (1), 147-161 (2003).
- Wu, Q., Zhang, Y., Xu, J., Shen, P. Regulation of hunger-driven behaviors by neural ribosomal S6 kinase in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 102 (37), 13289-13294 (2005).
- Lingo, P. R., Zhao, Z., Shen, P. Co-regulation of cold-resistant food acquisition by insulin- and neuropeptide Y-like systems in Drosophila melanogaster. Neuroscience. 148 (2), 371-374 (2007).
Tags
Neurovidenskab Problem 160 Drosophila larver termogentisk neuronal skærm larvefodringsadfærd makronæringsstofbalance protein kulhydrater fødeindtagelsesanalyse madfarvestof kolorometrisk kvantificeringErratum
Formal Correction: Erratum: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae
Posted by JoVE Editors on 10/06/2020.
Citeable Link.
An erratum was issued for: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae. A figure was updated.
Figure 1 was updated from:
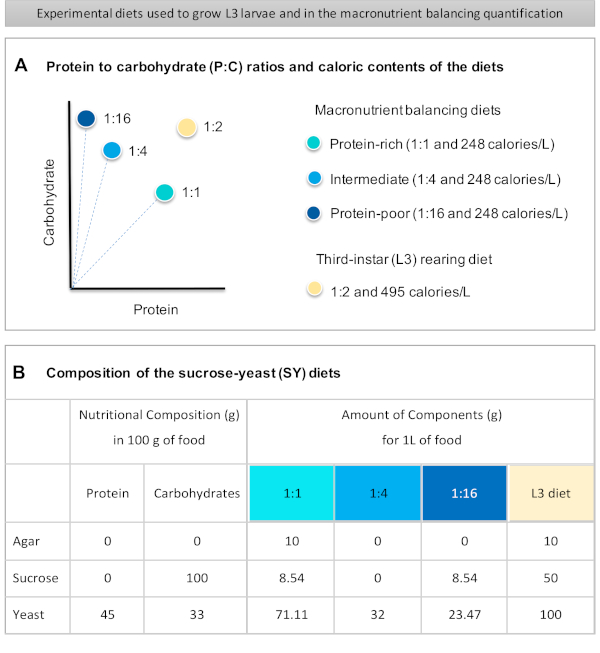
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
to:
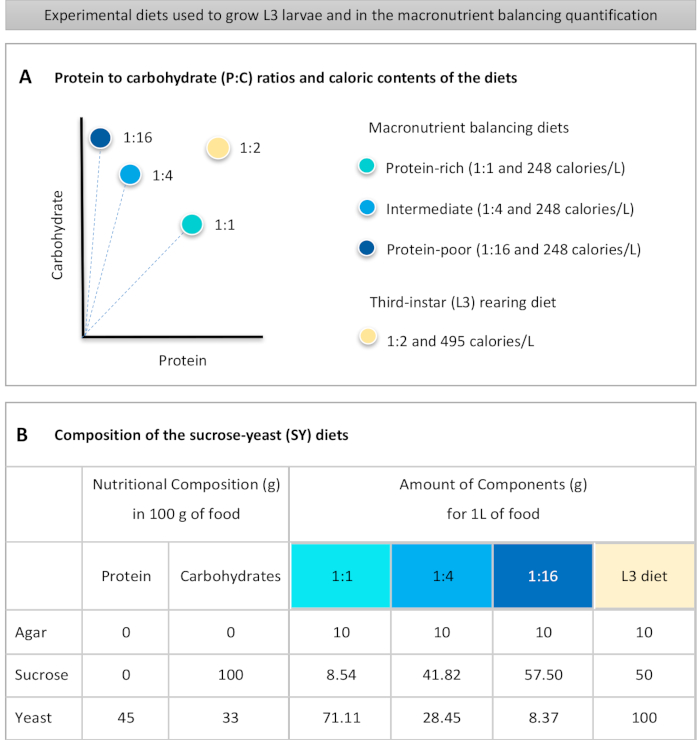
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
