

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Hier wordt een protocol beschreven dat de colorimetrische kwantificering mogelijk maakt van de hoeveelheid voedsel die binnen een bepaald tijdsinterval wordt gegeten door Drosophila melanogasterlarven die worden blootgesteld aan diëten van verschillende macronutriëntenkwaliteit. Deze assays worden uitgevoerd in de context van een neuronaal thermogenetisch scherm.
Abstract
Foerageer- en voedingsgedrag stelt dieren in staat om toegang te krijgen tot energiebronnen en voedingsstoffen die essentieel zijn voor hun ontwikkeling, gezondheid en fitheid. Het onderzoeken van de neuronale regulatie van dit gedrag is essentieel voor het begrijpen van de fysiologische en moleculaire mechanismen die ten grondslag liggen aan nutritionele homeostase. Het gebruik van genetisch trekbare diermodellen zoals wormen, vliegen en vissen vergemakkelijkt dit soort studies aanzienlijk. In het afgelopen decennium is de fruitvlieg Drosophila melanogaster gebruikt als een krachtig diermodel door neurobiologen die de neuronale controle van voedings- en foerageergedrag onderzoeken. Hoewel ongetwijfeld waardevol, onderzoeken de meeste studies volwassen vliegen. Hier beschrijven we een protocol dat gebruik maakt van het eenvoudigere larvale zenuwstelsel om neuronale substraten te onderzoeken die het voedingsgedrag controleren wanneer larven worden blootgesteld aan diëten die verschillen in hun eiwit- en koolhydraatgehalte. Onze methoden zijn gebaseerd op een kwantitatieve colorimetrische no-choice voedingstest, uitgevoerd in de context van een neuronaal thermogenetisch activeringsscherm. Als uitlezing werd de hoeveelheid voedsel die door larven gedurende een interval van 1 uur werd gegeten, gebruikt bij blootstelling aan een van de drie diëten met een kleurstoflabel die verschillen in hun eiwit-koolhydratenverhoudingen (P:C). De werkzaamheid van dit protocol wordt aangetoond in de context van een neurogenetisch scherm in larvale Drosophila, door kandidaat-neuronale populaties te identificeren die de hoeveelheid voedsel reguleren die wordt gegeten in diëten van verschillende macronutriëntenkwaliteit. We waren ook in staat om de geteste genotypen te classificeren en te groeperen in fenotypische klassen. Naast een korte bespreking van de momenteel beschikbare methoden in de literatuur, worden de voor- en nadelen van deze methoden besproken en worden ook enkele suggesties gedaan over hoe dit protocol kan worden aangepast aan andere specifieke experimenten.
Introduction
Alle dieren zijn afhankelijk van een uitgebalanceerd dieet om de nodige hoeveelheden voedingsstoffen te verkrijgen voor overleving, groei en voortplanting1. De keuze van wat en hoeveel te eten wordt beïnvloed door een veelheid aan interactiefactoren die verband houden met de interne toestand van het dier, zoals het verzadigingsniveau en omgevingsomstandigheden, zoals voedselkwaliteit2,3,4,5. Eiwitten en koolhydraten zijn twee belangrijke macronutriënten en de evenwichtige inname ervan is essentieel om de fysiologische processen van dieren te ondersteunen. Daarom is het begrip van de neurale mechanismen die het voedingsgedrag controleren en een evenwichtige inname van deze macronutriënten ondersteunen uiterst relevant. Dit komt omdat levensgeschiedeniskenmerken zoals levensduur, vruchtbaarheid en metabole gezondheid direct worden beïnvloed door de niveaus van eiwitinname6,7,8,9,10.
Het gebruik van eenvoudigere meer tracteerbare organismen die evolutionair geconserveerde voedingsgewoonten vertonen bij complexe dieren, waaronder zoogdieren, is essentieel voor dit soort studies. Belangrijk is dat deze eenvoudigere diermodellen een goede gelegenheid bieden om complexe biologische vragen te ontleden in een dure, ethische en technisch effectievere context. In de afgelopen decennia is Drosophila, met zijn krachtige genetische toolkit, ingewikkeld en stereotiep gedrag en geconserveerde architectuur van perifere en nutriëntendetectiemechanismen met zoogdieren, een vruchtbaar model geweest voor gedragsneurobiologen11. Uiteindelijk is de hoop dat door te begrijpen hoe voedselinname bij dit dier wordt gereguleerd, met een eenvoudiger zenuwstelsel, we dan kunnen beginnen met het ontwarren van neuronale storingen die ten grondslag liggen aan menselijke eetstoornissen.
De studie van neuronale substraten voor voedingsgedrag is sterk afhankelijk van het tegelijkertijd kunnen meten van de voedselinname van dieren terwijl ze hun neuronale activiteit manipuleren. Vanwege de minimale hoeveelheden voedsel die worden ingenomen, is het kwantificeren van de hoeveelheid voedsel die door vliegen wordt gegeten uiterst uitdagend en alle methoden die momenteel beschikbaar zijn, vertonen aanzienlijke beperkingen. De gouden standaard is dus het gebruik van een combinatie van complementaire methodologieën12. Volwassen vliegen zijn historisch begunstigd als een genetisch en gedragsmodel. Niettemin bieden Drosophila-larven ook mogelijkheden om neuronale substraten te onderzoeken die coderen voor voedingsgedrag. Het larvale centrale zenuwstelsel (CZS), met ongeveer 12.000 neuronen, is aanzienlijk minder complex dan dat van de volwassene, die ongeveer 150.000 neuronen bevat. Deze lagere complexiteit is niet alleen numeriek, maar ook functioneel, omdat larvaal gedrag afhankelijk is van eenvoudigere locomotieffuncties en sensorische systemen. Ondanks de schijnbare eenvoud van hun zenuwstelsel vertonen larven nog steeds volledig voedingsgedrag, en sommige methoden om voedselinname in Drosophila-larven te kwantificeren zijn beschreven5,13,14,15. Door te paren met manipulaties van neuronale activiteit, kunnen Drosophila-larven een zeer tracteerbaar model vormen voor het begrijpen van de neurale regulatie van voedselinname.
Hier is een gedetailleerd protocol beschikbaar om de voedselinname te kwantificeren bij larven die worden blootgesteld aan diëten van verschillende macronutriëntenkwaliteit. De diëten, zogenaamde macronutriëntenbalanceringsdiëten, verschilden in de eiwit- en koolhydrateninhoud, met name met betrekking tot de eiwit-koolhydraatverhoudingen (P:C): 1:1 (eiwitrijk dieet), 1:4 (tussendieet) en 1:16 (eiwitarm dieet), zoals weergegeven in figuur 1A. Kortom, een kwantitatieve no-choice voedingstest werd vastgesteld met behulp van deze drie isocalorische sucrose-gist (SY) gebaseerde diëten geverfd met een blauwe voedselkleurstof. Omdat gistextract en sacharose werden gebruikt als eiwit- en koolhydraatbronnen en beide koolhydraten bevatten, werd variatie in de P:C-verhoudingen verkregen door de balans van deze twee componenten te veranderen, zoals eerder beschreven16 en zoals aangegeven in figuur 1B. Een schematisch overzicht van het protocol, met de belangrijkste experimentele stappen, is beschikbaar in figuur 2.
Dit protocol is opgesteld met als doel de rol van specifieke neuronale populaties te onderzoeken op de regulering van larvale voedingsniveaus in diëten met verschillende P:C-verhoudingen en in de context van een thermogenetisch neuronaal scherm. Een goed gekarakteriseerd neurogenetisch hulpmiddel werd gebruikt uit de Transient Receptor Potential (TRP) familie: Drosophila Transient Receptor Potential channel (dTRPA1), een temperatuur- en spanningsge gated cation kanaal, waardoor het afvuren van actiepotentiaal wanneer de omgevingstemperatuur boven 25 °C17stijgt. Om de dTRPA1 transgene uit te drukken, hebben we gebruik gemaakt van de Gal4-lijnen op basis van cis-regulerenderegio's uit het Drosophila-genoom, gevestigd in het Rubin-laboratorium, in het kader van het FlyLight-project op Janelia Research Campus18,19.
Hoewel het protocol, hier beschreven, is vastgesteld in de context van een activeringsscherm, kan het door de experimenteerder gemakkelijk worden aangepast aan andere specifieke behoeften of interesses, namelijk om een onderdrukkingsscherm uit te voeren met behulp van de temperatuurgevoelige neuronale geluiddemper ShibireTS20, in alternatief voor dTRPA1. Deze en andere aanpassingen worden besproken in de protocol- en discussiesecties.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Bereiding van het sucrose-gist (SY) dieet
- Weeg alle droge ingrediënten (agar, gist, sacharose) voor de macronutriëntenbalancering en L3-opfokdiëten. De hoeveelheden in gram voor elk van de ingrediënten die nodig zijn om 1 L voedsel te bereiden , zijn aangegeven in figuur 1B.
OPMERKING: Houd er rekening mee dat er ongeveer 13 ml voedsel nodig is om een petrischaal van 60 mm te vullen. - Los alle ingrediënten op in steriel gedestilleerd water (gebruik ongeveer 50% van het totale volume water dat nodig is om het voedsel te bereiden) en roer het medium gedurende 5-10 minuten.
- Autoclaaf gedurende 50 minuten.
- Nadat u de mediums hebt laten afkoelen, voegt u nipagine- en propionzuuroplossingen toe aan de diëten, bij een eindconcentratie (v/v) van respectievelijk 3% en 0,3%. Voeg aan de macronutriëntenbalanceringsdiëten blauwe voedselkleurstof toe aan een eindconcentratie (v/v) van 1%. Voltooi de totale volumes met gedestilleerd water.
- Giet de voedseldiëten zorgvuldig naar 60 mm Petri-gerechten, zodat de hoeveelheid gegoten voedsel ongeveer hetzelfde is in elk van de borden. Label de borden met de P:C-verhoudingen van de diëten.
OPMERKING: Bereid de macronutriëntenbalanceringsdiëten voor op de dag van de voedingstest. Indien niet mogelijk, bewaar de bereide diëten bij 4 °C, in een verzegelde container, voor een maximale duur van 3 dagen. Langere opslagperioden maken het dieet te droog en hard en de larven kunnen zich niet in het medium graven.
2. Genetisch kruis van ouderlijke lijnen
OPMERKING: Gebruik het Gal4/UAS-systeem21 om de genetische kruisen in te stellen. In dit protocol, om neuronale functie in specifieke neuronale populaties te activeren, werden vrouwelijke maagden van de UAS dTRPA1 lijn17 gebruikt en gekruist naar mannen van de Janelia Gal4-lijnen (Figuur 2A). De gebruikte genetische controle was het nageslacht van een kruising tussen de dTRPA1-lijn en een "lege GAL4"-lijn, die Gal4 draagt in de vector die wordt gebruikt om de Rubin Gal4-collectie te genereren, maar zonder regelgevend fragment aanwezig (attP2)22. Om de neuronale onderdrukking te bevorderen, kan een UAS-lijn worden gebruikt die codeert voor ShibireTS20, in plaats van dTRPA1.
- Zet 60 mm embryo-verzamelkooien op met L3-opfokdieetplaten, aangevuld met wat actieve gistpasta.
- Breng de volwassen UAS dTRPA1 vrouwelijke maagden en Janelia Gal4-mannetjes van 5-8 dagen over naar de embryo-verzamelkooien en laat de paring gedurende 24-48 uur plaatsvinden, bij 25 °C, met een luchtvochtigheid van 60% en een licht-donkere cyclus van 12:12 uur (figuur 2A). Gebruik voor 60 mm embryo-verzamelkooien ongeveer 100 maagdelijke vrouwtjes en 30 mannetjes per kruis.
- Verwijder aan het einde van de paarperiode de L3-opfokdieetplaten die in de genetische kruisen worden gebruikt en gooi deze weg. Vervang ze door verse L3-opfokdieetplaten om de eier legt en larvale enscenering uit te voeren.
3. Bereiding van larven van derde instar (L3)
- Breng de gepaarde volwassen vliegen over op verse L3-opfokdieetplaten en laat het leggen van eieren gedurende 3-4 uur plaatsvinden bij 25 °C (figuur 2B). Zorg ervoor dat alle borden zijn gelabeld met het genotype, P: C-verhouding van het dieet en de datum van het legen van het ei.
OPMERKING: Om tijd te besparen, voert u het leggen van eieren rechtstreeks uit in het L3-opfokdieet, waardoor extra behandeling van de eieren wordt voorkomen. In het geval van kleinschalige genetische screenings kan optimalisatie van het leggen van eieren worden verkregen door gebruik te maken van appelsap agar platen. - Verwijder aan het einde van de legperiode de platen uit de kooien en bedek ze met plastic deksels. In het geval dat gistextract wordt gebruikt om de L3-opfokplaten aan te vullen, zorg er dan voor dat u alle resterende gist aan het einde van de eierlegging verwijdert. Dit is belangrijk om niet-uniforme voeding tijdens de larvale groei te voorkomen.
OPMERKING: De gedekte volwassenen kunnen worden overgebracht naar verse L3-opfokdieetplaten, zodat er meer eier legt en meer experimentele larven kunnen worden verkregen. Opeenvolgende eierlegt kan gedurende één hele werkweek met dezelfde volwassenen worden uitgevoerd. - Schat het aantal eieren per plaat en houd de larvale dichtheid op maximaal 200 embryo's per plaat. Deze schatting kan worden gedaan door het aantal embryo's in een kwart van de plaat te tellen.
OPMERKING: Een overvolle plaat vertraagt de ontwikkeling van larven en beïnvloedt het voedingsgedrag van larven. - Incubeer de L3-opfokplaten bij 18 °C (permissieve temperatuur), 60% vochtigheid en een licht-donkercyclus van 12:12 en laat de larven 9 dagen groeien (Figuur 2B).
- Verzamel op de negende dag na het leggen van eieren (AEL) drie groepen van 10 L3 van elk van de genotypen (en voor de duplo's) die moeten worden getest. Verzamel bovendien groepen van 10 L3 voor de controle van "zero-dye food". Zorg ervoor dat de larvenverzameling wordt gedaan tijdens gelijkwaardige perioden van de dag die worden gebruikt om het ei te leggen (bijv. als het leggen van het ei plaatsvond tussen 10.00 en 14.00 uur, verzamel de larven gedurende dezelfde periode van 9 dagen AEL) en wordt zo voorzichtig mogelijk uitgevoerd met behulp van een tang #5 of een vedergewicht tang. Breng de larven direct over zoals aangegeven in de volgende stap (3.6).
OPMERKING: De controledieren "zero-dye food" zijn larven die in de voedertest voedsel krijgen zonder blauwe kleurstof. Deze controle is noodzakelijk om de achtergrondabsorpance van de larvale extracten te verwijderen. - Breng de verzamelde experimentele larven over in plastic schaalgewicht boten met 1 ml water. Zorg ervoor dat L3 wordt verzameld en niet L2, door de aanwijzingen in figuur 3te volgen .
OPMERKING: Het verzamelen van L3 naar plastic boten met water of 1x Fosfaatgebufferde zoutoplossing (PBS) is belangrijk om larven goed gehydrateerd te houden voor het begin van de voedertest. Dit is vooral belangrijk als meerdere experimentele L3-groepen van verschillende genotypen tegelijkertijd worden verzameld. Houd de inzamelingsvolgorde voor elke groep bij, zodat verschillen in de duur van voedseltekort voor elke groep worden geminimaliseerd. Het gebruik van plastic boten in deze stap vergemakkelijkt de optionele stap 4.3 omdat het larven in staat stelt om direct in het waterbad te drijven.
4. Thermogenetische activering en no-choice voedingstest
OPMERKING: Het wordt aanbevolen om de voedingstesten op ongeveer hetzelfde tijdstip van de dag uit te voeren om mogelijke variaties met betrekking tot de circadiaanse ritmes te minimaliseren. Voer ook altijd de controle-experimenten uit (het nageslacht van de "lege Gal4"-lijn die is overgestoken naar UAS dTRPA1 en de larven van "zero-dye food"), parallel aan de genotypen van belang.
- Zet een incubator op tot 30 °C (niet-permissieve temperatuur) en houd een hoge luchtvochtigheid (ten minste 65%) om uitdroging van larven tijdens de test te voorkomen.
- Voordat u met de voedertest begint, moet u de temperatuur van de testplaten gelijkmaken door ze gedurende 30 minuten op 30 °C te verwarmen.
- (Optioneel) Hitteschok de experimentele larven gedurende 2 minuten in een waterbad van 37 ° C. Voer deze stap uit met de dieren in de plastic gewichtsboten die wat water bevatten.
OPMERKING: Het doel van deze stap is om de neuronale activering te intensiveren door het afvuren van de neuronen sinds het begin van de voedingstest te bevorderen. - Houd meerdere timers ingesteld op 1 uur. Het aantal te gebruiken timers hangt af van het aantal experimentele groepen dat wordt getest en van het vaardigheidsniveau van de experimenteerder bij het hanteren van larven.
OPMERKING: Het gebruik van meerdere timers is van cruciaal belang om de duur van de test consistent te houden voor alle genotypen. - Giet het water voorzichtig af van de plastic boten en breng met behulp van een bevochtigde zachte borstel de L3-groepen voorzichtig over van de boten naar het midden van de testplaten. Plaats de deksels van de platen terug en start een timer voor elke plaat (of groep platen) om nauwkeurige 1-uurs voedingssessies te behouden.
- Laat de larven zich gedurende 1 uur, bij 30 °C, in het donker voeden (figuur 2C).
OPMERKING: De prestaties van de test in het donker zijn belangrijk om te controleren op verschillen in de visuele signalen tussen diëten, omdat de diëten in tonen zullen verschillen, ook al bevatten ze dezelfde kleurstofconcentratie. - Stop de voedingstest door de platen over te brengen naar een ijsbad. Druk het ijs zoveel mogelijk naar beneden om een stabiel oppervlak voor de platen te bieden.
OPMERKING: Koude temperaturen bevorderen het einde van de voeding door het graaf- en graafgedrag te remmen. De meeste larven zullen na enkele minuten de voedselplaten aan de oppervlakte brengen, waardoor hun herstel in de volgende stappen wordt vergemakkelijkt.
5. Extractie van voedselkleurstoffen
- Bereid 2 ml microtubes voor elke geteste groep van 10 L3, met ongeveer dezelfde hoeveelheden 0,5 mm glazen kralen (genoeg om het onderste gedeelte van de microbuis te vullen) en 300 μL ijskoude methanol. Houd de microtubes in de kou met behulp van een bankkoeler.
LET OP: Methanol is licht ontvlambaar en giftig. Volg alle veiligheidsprocedures die worden aanbevolen voor het gebruik van dit reagens, inclusief het werken in een goed geventileerde ruimte en het dragen van nitrilhandschoenen.
OPMERKING: Het gebruik van methanol is belangrijk om de larvale monsters te fixeren en melanisatiereacties in de nagelriem te voorkomen. - Met behulp van #5 of vedergewicht-tang, zorgvuldig herstellen van de groepen van 10 L3 van de voedertest platen en breng ze over naar de deksels van de assay platen met wat water. Spoel de larven af om voedselresten op hun lichaam te verwijderen terwijl ze voorzichtig met de larven omgaan om verwondingen te voorkomen. Houd het aantal teruggevonden larven per genotype per replica bij, zodat de gemiddelde hoeveelheid voedselinname per larve kan worden gekwantificeerd.
OPMERKING: Gewonde larven moeten worden weggegooid omdat ze een gemelaniseerde cuticula hebben, die niet geschikt zijn voor colorimetrische kwantificering. - Breng de L3-groepen over naar de microbuisjes van 2 ml die in 5.1 zijn bereid.
- Lyse de larvale weefsels om de voedselkleurstof uit de darmen te extraheren door middel van een mechanische lysismethode met behulp van een weefsellyzer en glasparels toegevoegd in stap 5.1. (als er geen tissue lyzer beschikbaar is, gebruik dan een homogeniserende stamper). Voer deze stap bij voorkeur uit bij 4 °C (figuur 2D).
OPMERKING: De duur van deze stap is afhankelijk van de gebruikte apparatuur. Met behulp van een conventionele weefsellyser is 1 min extractie voldoende. In geval van tijdsvernauwing kan het protocol aan het einde van deze stap worden onderbroken en later worden voortgezet. Bewaar de monsters bij -20 °C. - Breng de extracten over om 1,5 ml microtubes schoon te maken, door de 2 ml microtubes direct om te keren naar de nieuwe 1,5 ml microtubes. Indien voorzichtig uitgevoerd, blijven de meeste glazen kralen aan de onderkant van de 2 ml microbuis.
- Verwijder het celafval door de extracten bij een maximale snelheid van 10 minuten bij 4 °C te centrifugeren.
- Verzamel de supernatanten om 1,5 ml microtubes schoon te maken. Als er nog celresten zichtbaar zijn in de supernatanten, herhaalt u stap 5.6 en 5.7.
6. Colorimetrische kwantificering van voedselconsumptie
- Bereid standaardoplossingen voor, om een kalibratiecurve te genereren, door serieel 1:2 verdunningen uit te voeren in methanol van een beginnende blauwe kleurstofoplossing. Gebruik methanol alleen als blanco. De concentratie van de normen is afhankelijk van de voedselinname van de dieren.
OPMERKING: In het geval van het hier gepresenteerde proefscherm werd een standaardcurve gebruikt die werd verkregen door de absorpties van 8 seriële verdunningen van een blauwkleurstofoplossing van 2 μL/ml in methanol te meten, aangezien de voor de larvale extracten verkregen kleurstofconcentraties varieerden van 0,02 tot 1,93 μL/ml. Verhoog of verlaag indien nodig de concentratie van deze oplossingen, afhankelijk van de kleurstofconcentratie van de experimentele monsters. - Breng 100 μL van de experimentele monsters (verkregen in stap 5.7), normen en blanco (stap 6.1) over naar de putten van een 96-put microplaat en meet de absorptie bij 600 nm, met behulp van een plaatlezer (figuur 2E). Om de achtergrondabsorpance te verwijderen, meet u de absorptie van extracten verkregen uit larven die worden gevoed met voedsel zonder blauwe kleurstof als een "nul" voor de larvale extracten ("zero-dye food control").
- Genereer een standaardcurve en correleer de absorptiewaarden die zijn verkregen voor de monsters van elke experimentele larvale groep met de hoeveelheid voedselinname (volume in ml). Zoek de gemiddelde voedselconsumptie per larve door rekening te houden met het aantal larven dat voor elke groep is verzameld in stap 5.2
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Drosophila-larven reguleren hun eiwitinname ten koste van de inname van overtollige koolhydraten23 (schematisch plot in figuur 2E). Eigenlijk is deze prioritering van eiwitinname waargenomen bij veel andere dieren en wordt het eiwit24,25genoemd .
Gebruikmakend van deze robuuste voedingsgedragsrespons, werd een op gedrag gebaseerd scherm ontworpen om neuronale populaties te identificeren die betrokken zijn bij macronutriëntenbalancering. Er werd een no-choice voedingstest vastgesteld, die bestond uit het toestaan van groepen L3 (10 individuen per groep) om ad libitum gedurende 1 uur te voeden en onder neuronale thermogenetische activeringsomstandigheden met behulp van dTRPA1, in drie isocalorische (248 Cal/L) voedselgeverfde diëten met specifieke P:C-verhoudingen (1:1, 1:4 en 1:16) (Figuur 1 en Figuur 2). Als uitlezing werd de gemiddelde hoeveelheid voedsel gebruikt die werd gegeten in de macronutriëntendiëten van verschillende P:C-verhoudingen. Gebruikmakend van het Gal4/UAS-systeem21 en met behulp van enkele van de Janelia Gal4-lijnen uit het FlyLight Project18,19, werd de expressie van dTRPA1 geïnduceerd in specifieke neuronale populaties.
Met de methoden die in dit protocol worden beschreven, konden we de relatieve hoeveelheid macronutriënten kwantificeren die in termen van P:C-verhoudingen werden geconsumeerd voor dieren onder thermogenetische activering van specifieke neuronale populaties in het larvale zenuwstelsel. Deze experimentele benadering toonde aan dat het activeren van verschillende populaties neuronen de macronutriëntenbalancering in larven van derde instar aanzienlijk beïnvloedde (figuur 4, tabel 1). Het voedingspatroon dat is waargenomen voor de controlelijn (attP2) toont de effectiviteit van de methode aan door een verwachte compenserende toename van de voedselinname door larven die zijn getest in diëten met een lagere P:C-verhouding (grijze stippen en lijn in figuur 4) aan te tonen. Bovendien werd een significante interactie tussen de genotypen en het dieet gevonden, wat betekent dat de thermogenetische activering van specifieke neuronale populaties de manier verandert waarop larven hun voedselinname reguleren als reactie op de macronutriëntenkwaliteit van het dieet.
De voedingspatronen van de genotypen die zijn getest in de drie macronutriëntenbalanceringsdiëten (1:1, 1:4, 1:16) worden weergegeven door de gekleurde stippen en lijnen in figuur 4 en de statistische analyse is beschikbaar in tabel 1.
In het activeringsscherm werden in totaal 36 Janelia Gal4-lijnen getest waarvan bekend is dat ze schaars zijn uitgedrukt in het larvale zenuwstelsel. Met behulp van lineaire regressiemodellen hebben we bepaald welke genotypen significant verschillende voedselinname vertoonden ten opzichte van de genetische controledieren. Deze verschillen omvatten ofwel verschillen in de absolute hoeveelheid voedsel die in alle diëten wordt gegeten, ofwel verschillen in de macronutriëntenbalanceringsrespons (helling van de respons op de verschillende P:C-verhoudingen van de diëten).
In alle drie de diëten at R12E06 aanzienlijk meer voedsel dan controledieren. Bovendien overcompenseerde het de toename van de voedselinname op het tussenliggende en eiwitarme dieet, zoals blijkt uit een significant verschil in de interactietermijn tussen voedselinname en P:C-verhouding van het dieet (Tabel 1). R22H01 at aanzienlijk meer dan de controles, maar verschilde niet in de macronutriëntenbalanceringsrespons (Tabel 1). R14B11, R19G11, R21B06, R29C02 en R48F09 larven aten kleine hoeveelheden voedsel en verloren het vermogen om de slechte macronutriëntenkwaliteit van het beschikbare dieet te compenseren (zoals aangegeven door de significante interactietermen tussen voedselinname en P:C-verhouding van het dieet, tabel 1). Ten slotte aten R45D11-larven aanzienlijk meer in het eiwitrijke dieet met een P:C-verhouding van 1:1 dan in de tussenliggende en in de eiwitarme diëten (1:4 en 1:16), wat het tegenovergestelde is van wat men zou verwachten op de eiwitarme diëten.
Daarom stelden onze methoden ons in staat om de experimentele larven, van elk genotype, te classificeren in fenotypische klassen met betrekking tot de totale hoeveelheid gegeten voedsel en het vermogen om prioriteit te geven aan eiwitinname door te veel te nemen in de diëten met een lage P: C-verhouding. Voor de proefdieren werden vijf fenotypische klassen vastgesteld (figuur 5): 1 – "Eet veel" (meer dan de controledieren) en overcompenseren voor eiwitverdunning; 2 – "Eet veel, maar compenseer normaal"; 3 – "Eet weinig (minder dan de controle) maar compenseer"; 4 – "Eet weinig en compenseer niet"; 5 – "Eet afwijkend" (meer in eiwitrijke en tussenliggende diëten dan in het eiwitarme dieet). Bovendien tonen we voor elk van deze fenotypische klassen en genotypen de GFP-patronen in het centrale zenuwstelsel van derde-instar-larven. Deze informatie werd verkregen uit de openbaar beschikbare beeldvormingsgegevens in het online platform van FlyLight Project, waar men toegang kan krijgen tot de expressiepatronen van alle Rubin Gal4-lijnen van belang26.

Figuur 1: De sucrose-gist (SY) diëten die in ons protocol worden gebruikt. (A) De blauwe stippen vertegenwoordigen de isocalorische (248 calorieën/L) macronutriëntenbalancerende diëten die worden gebruikt bij de voedingstest, die verschillen in de eiwit-koolhydraatverhoudingen (P:C): 1:1, 1:4 en 1:16. De beige stip vertegenwoordigt het dieet dat wordt gebruikt om de experimentele derde-instarlarven (L3) op te wekken, die een P:C-verhouding van 1:2 en een calorische dichtheid van 495 calorieën/L. (B) bevatten Gedetailleerde samenstelling en voedingsinformatie van de op sacharosegist (SY) gebaseerde diëten. De componenten zijn hetzelfde voor alle diëten: agar, sucrose en gist. De hoeveelheid in gram van de componenten die nodig zijn om 1 L dieet te bereiden, wordt weergegeven. Merk op dat 1% (v/v) blauwe kleurstof moet worden toegevoegd aan de macronutriëntenbalanceringsdiëten en aan het L3-opfokdieet moeten nipagine- en propionzuuroplossingen worden toegevoegd aan een eindconcentratie (v/v) van respectievelijk 3% en 0,3%. Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 2: Schematische weergave van de belangrijkste stappen in ons protocol (A) Genetisch kruis van ouderlijke lijnen die gebruik maken van het Gal4/UAS-systeem. Het kruis tussen de Rubin Gal4-lijnen en de UAS-lijn die codeert voor dTRPA1, maakt de thermogenetische activering van specifieke neuronale populaties in het larvale centrale zenuwstelsel mogelijk. (B) Bereiding van de experimentele derde-instarlarven (L3). De vrouwtjes van de ouders mochten 3-4 uur eieren leggen en de larvale enscenering vindt plaats bij de permissieve temperatuur (18 °C) gedurende 9 dagen. Optioneel is de hitteschok bij 37 °C gedurende 2 minuten voor de voedertest. (C) Thermogenetische activering van de neuronale functie en no-choice voedingstest gedurende 1 uur bij de niet-permissieve temperatuur (30 °C). Drie groepen van 10 experimentele L3 van elk genotype mochten voeden in elk van de macronutriëntenbalancerende diëten die specifieke eiwit-koolhydraten (P:C) verhoudingen bevatten (1:1, 1:4 en 1:16). (D) Extractie van levensmiddelenkleurstoffen. Mechanische lysis van larven, met behulp van een weefsellyser, om de blauwe voedselkleurstof te extraheren. (E) Kwantificering van de voedselinname. Colorimetrische kwantificering van de gemiddelde hoeveelheid voedsel die per larve wordt gegeten door de concentratie van voedselkleurstoffen in de larvale extracten te kwantificeren. De absorptie van de experimentele monsters, normen en "nul" werd gemeten bij 600 nm (blauw), met behulp van een 96-put plaatlezer. Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 3: Verschillen tussen tweede (L2) en derde-instars Drosophila larven (L3). De L2 en L3 zijn gemakkelijk te onderscheiden door de observatie van spiracles onder een stereomicroscoop. De voorste spiracles van L2 zijn club-achtig, terwijl in L3 zijn vertakt. Andere kenmerken kunnen helpen om de twee instars te onderscheiden, maar zijn subjectief en minder betrouwbaar. De achterste spiracles van L3 hebben een donkeroranje ring aan hun punt, die ontbreekt of zwak aanwezig is in de L2. De luchtpijp is dikker in L3-larven. Illustratie door Marisa Oliveira. Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 4: Hoeveelheid voedsel gegeten per larve onder neuronale thermogenetische activeringsomstandigheden in drie macronutriëntenbalancerende diëten die specifieke eiwit-koolhydratenverhoudingen (P:C) bevatten. Gemiddelde hoeveelheid voedsel die per larve (ml) wordt gegeten in 3 macronutriëntenbalanceringsdiëten die de specifieke P:C-verhoudingen van 1:1, 1:4 en 1:16 bevatten. Groepen van 10 derde-instarlarven, van elk genotype, mochten gedurende 1 uur voeden, onder neuronale thermogenetische activeringsomstandigheden, met behulp van dTRPA1, bij 30 °C. De geteste genotypen (larvale nakomelingen van de genetische kruisingen tussen de Rubin Gal4-lijnen en de UAS dTRPA1-lijn) worden aangegeven door stippen en lijnen van verschillende kleuren. Als genetische controle (aangegeven in grijs) werd het larvale nageslacht van een kruising tussen de "lege Gal4"-lijn (attP2) en UAS dTRPA1 gebruikt. De namen die aan de genotypen werden gegeven, aangegeven in de legende, waren gerelateerd aan de gebruikte "Rubin GAL4"-lijnen. Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 5: Groepering van de geteste lijnen in 5 belangrijkste fenotypische klassen. De fenotypische klassen aangegeven door cijfers waren gebaseerd op de combinatie van de fenotypes waargenomen in termen van totale hoeveelheid gegeten voedsel en het vermogen om de prioriteringsrespons van de eiwitinname te behouden: 1 - veel eten (meer dan de controledieren) en in staat waren om eiwitverdunning te compenseren door te veel te eten; 2 – veel eten en niet in staat waren om te compenseren; 3 - eet weinig (minder dan de controle) maar compenseer; 4 – eet weinig en waren niet in staat om te compenseren; en 5 - een extra fenotypische klasse, die "afwijkend" werden genoemd, waarbij de larven zich niet gedroegen zoals verwacht als reactie op de macronutriëntenverdunning van eiwitgehalte in het dieet, waarbij ze meer aten in eiwitrijke en tussenliggende diëten dan in het eiwitarme dieet. Voor elk genotype wordt het GFP-expressiepatroon in het centrale zenuwstelsel van derde-instar-larven getoond. Deze beeldvormingsgegevens van de Rubin Gal4-lijnen die in deze test worden gebruikt, zijn afkomstig van het openbaar beschikbare FlyLight Project online platform26. Klik hier om een grotere versie van deze afbeelding te bekijken.
| Anova-tabel (tests van type II) | |||||
| Reactie: Concentratie/L3 | |||||
| Som Sq | Df | F-waarde | Pr(>F) | ||
| voedsel | 0.086832 | 1 | 113.5358 | < 2.2e-16 | *** |
| genotype | 0.078443 | 10 | 10.2567 | 9.762e-15 | *** |
| Eten : Genotype | 0.064038 | 10 | 8.3733 | 6.416e-12 | *** |
| Resten | 0.215673 | 282 | |||
| Betekeniscodes: 0 '***' 0,001 '**' 0,01 '*' 0,05 '.' 0.1 ' ' 1 | |||||
| Overzichtstabel (onderstaande coëfficiënten worden vergeleken met het attP-controlegenotype): | |||||
| schatten | Std. Fout | t waarde | Pr(>|t|) | ||
| (Onderscheppen) | 0.064245 | 0.004316 | 14.886 | < 2e-16 | *** |
| voedsel | -0.058117 | 0.007206 | -8.066 | 2.10e-14 | *** |
| Genotype R12E06 | 0.040243 | 0.008961 | 4.491 | 1.03e-05 | *** |
| Genotype R14B11 | -0.053347 | 0.014361 | -3.715 | 0.000245 | *** |
| Genotype R19G11 | -0.044880 | 0.010788 | -4.160 | 4.23e-05 | *** |
| Genotype R21B06 | -0.051912 | 0.009363 | -5.544 | 6.79e-08 | *** |
| Genotype R22H01 | 0.017682 | 0.007296 | 2.423 | 0.016004 | * |
| Genotype R29C02 | -0.043102 | 0.011113 | -3.879 | 0.000131 | *** |
| Genotype R40D06 | -0.005341 | 0.009876 | -0.541 | 0.589102 | |
| Genotype R45C03 | 0.004064 | 0.009876 | 0.412 | 0.680997 | |
| Genotype R45D11 | -0.052579 | 0.009876 | -5.324 | 2.08e-07 | *** |
| Genotype R48F09 | -0.044612 | 0.011362 | -3.926 | 0.000108 | *** |
| Voeding : Genotype R12E06 | -0.037763 | 0.015440 | -2.446 | 0.015067 | * |
| Voeding : Genotype R14B11 | 0.058054 | 0.027100 | 2.142 | 0.033031 | * |
| Voeding : Genotype R19G11 | 0.051532 | 0.017726 | 2.907 | 0.003937 | ** |
| Voeding : Genotype R21B06 | 0.054403 | 0.015689 | 3.467 | 0.000607 | *** |
| Voeding : Genotype R22H01 | -0.020863 | 0.012377 | -1.686 | 0.092979 | . |
| Voeding : Genotype R29C02 | 0.048996 | 0.018714 | 2.618 | 0.009317 | ** |
| Voeding : Genotype R40D06 | 0.003804 | 0.016550 | 0.230 | 0.818371 | |
| Voeding : Genotype R45C03 | 0.034117 | 0.016550 | 2.061 | 0.040177 | * |
| Voeding : Genotype R45D11 | 0.090661 | 0.016550 | 5.478 | 9.53e-08 | *** |
| Voeding : Genotype R48F09 | 0.051184 | 0.019045 | 2.688 | 0.007625 | ** |
| Betekeniscodes: 0 '***' 0,001 '**' 0,01 '*' 0,05 '.' 0.1 ' ' 1 | |||||
| Restnormfout: 0,02765 op 282 vrijheidsgraden | |||||
| Meerdere R-kwadraat: 0.516, Aangepast R-kwadraat: 0.4799 | |||||
| F-statistiek: 14,31 op 21 en 282 DF, p-waarde: < 2,2e-16 | |||||
Tabel 1: ANOVA-tabel voor het effect van neuronale thermogenetische activering en macronutriëntenkwaliteit van het dieet beschikbaar op de hoeveelheid voedselinname. Er werd een lineair model aangebracht om de genotypen te bepalen die een aanzienlijk ander voedingsgedrag vertonen dan de controledieren.
| genotype | Geassocieerd gen | oorsprong | BDSC Voorraadnummer |
| w[*] ; P{UAS-TrpA1(B). K}attP2 / TM6B, Tb[1] | Bloomington | 26264 | |
| w[1118] ; P{GAL4.1Uw}t.a.v. T.v. | Janelia | 68384 | |
| w[1118] ; P{GMR12E06-GAL4}t.a.v. | netto (CG11450) | Janelia | NA |
| w[1118] ; P{GMR14B11-GAL4}attP2 / TM3, Sb[1] | dnc (CG32498) | Janelia | 49255 |
| w[1118] ; P{GMR19G11-GAL4}t.a.v. | CG33696 | Janelia | 48864 |
| w[1118] ; P{GMR21B06-GAL4}t.n.v. | oa2 (CG6919) | Janelia | 49857 |
| w[1118] ; P{GMR22H01-GAL4}t.a.v. | fru (CG14307) | Janelia | 49001 |
| w[1118] ; P{GMR29C02-GAL4}t.a.v. T.v. | Ptp69D (CG10975) | Janelia | 48088 |
| w[1118] ; P{GMR40D06-GAL4}t.a.v. | cnc (CG17894) | Janelia | 48616 |
| w[1118] ; P{GMR45C03-GAL4}t.a.v. T.v. | kni (CG4717) | Janelia | 47936 |
| w[1118] ; P{GMR45D11-GAL4}t.a.v. | pnt (CG17077) | Janelia | 49563 |
| w[1118] ; P{GMR48F09-GAL4}t.a.v. | dpr8 (CG32600) | Janelia | 50377 |
Tabel 2: Drosophila-lijnen die in dit werk worden gebruikt. Gedetailleerde informatie van alle gebruikte regels: codenaam, genotype, geassocieerd gen, oorsprong en het Bloomington Drosophila Stock Center (BDSC) nummer.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Met dit protocol zou men het vermogen van larven onder thermogenetische activering van specifieke neuronale populaties kunnen testen om de innameniveaus van eiwitten en koolhydraten, twee belangrijke macronutriënten, te reguleren wanneer ze worden blootgesteld aan diëten met verschillende P:C-samenstelling. Deze methode werd getest in de context van een larvale voorlopige screening met als doel neuronale populaties te identificeren die verband houden met de controle van voedselinname in verschillende diëten van verschillende macronutriëntenkwaliteit. Dit werk draagt ook bij aan het aantonen dat Drosophila-larven waardevolle diermodellen zijn voor het onderzoeken van de neuronale basis van voedingsgedrag geassocieerd met nutriëntenhomeostase.
Naast de informatie die als aantekeningen in het protocol wordt verstrekt, willen we enkele belangrijke aspecten verder bespreken. Zoals bij elke gedragstest, moeten de experimenteerders maatregelen nemen om de variatie in verband met dierlijk gedrag te minimaliseren. Een zeer belangrijk aspect dat men in gedachten moet houden, houdt verband met het belang van het verkrijgen van ontwikkelingssynchrone dieren. Het gebruik van vroege L3-larven die goed zijn gesynchroniseerd in hun ontwikkelingsfase, zal de gedragsvariaties verminderen die de dieren tijdens de voedertestvertonen 27. Het synchroniseren van larven wordt bereikt door korte eitjes en door de dichtheid van larven in de culturen te beheersen. Gebruik geen langere periodes van het leggen van eieren dan degene die we in het protocol aangeven (3-4 uur). Ook zal het beheersen van de larvale dichtheid tot een maximum aantal van 200 dieren per plaat ontwikkelingsachterstanden voorkomen en extra variatie in voedingsgedrag elimineren. Houd er rekening mee dat het eerste ei na de paring moet worden weggegooid om homogeniteit te behouden en een betere gesynchroniseerde larvale ontwikkeling te verkrijgen. Vrouwtjes hebben bevruchte eieren in eileider en leggen ze in verschillende stadia van ontwikkeling, waardoor het moeilijk is om uniformiteit tussen larvenverzameling te behouden. Het is absoluut noodzakelijk dat ten minste het eerste uur eieropvangplaat vóór de laatste inzameling wordt weggegooid. Houd er rekening mee dat de stress die bij de dieren wordt veroorzaakt tijdens het hanteren van larven ook een negatieve invloed kan hebben op het gedrag. Probeer zo zacht mogelijk te zijn, door een zachte en waterge bevochtigde borstel te gebruiken. Houd er ten slotte rekening mee dat een groot aantal replica's een betrouwbaardere gegevensset genereert.
Zoals in elk experimenteel protocol, vertonen onze methoden enkele beperkingen. Het gebruik van een colorimetrische methode om de voedselinname te kwantificeren op basis van de accumulatie van een voedselkleurstof in de darmen van dieren omvat enkele voorzorgsmaatregelen met betrekking tot de duur van de test. Voor volwassen vliegen werd aangetoond dat er een aanzienlijk risico bestaat op het bereiken van een stabiele toestand voor de accumulatie van kleurstoffen, waarbij de mate van egestie gelijk is aan de innamesnelheid, waardoor de nauwkeurigheid van de methode wordt verminderd28. Hoewel er geen bewijs is dat dit bij larven gebeurt, hebben we besloten om een voedingstest uit te voeren met een maximale duur van 60 minuten. Deze duur is handig en compatibel met een scherm met hoge doorvoer. Door de totale duur van het protocol zo kort mogelijk te houden, kunnen ook alle stappen in de secties 4, 5 en 6 in één werkdag worden voltooid. Indien de duur van de voedertest moet worden gewijzigd, moet de testduur van 60 tot 120 minuten een efficiënte kwantificering van de voedselinname tussen genotypen mogelijk maken, zoals eerder aangetoond29. De gevoeligheid van voedsel-stervende methoden is ook relatief laag wanneer kleine hoeveelheden voedsel worden geconsumeerd, wat de resolutie onder genotypen met zeer lage voedselinname aanzienlijk vermindert. We hebben onze voedingstesten opgezet met behulp van een no-choice paradigma. Er is slechts één dieettype beschikbaar voor elke experimentele groep larven, waardoor dieren de niveaus van de consumptie van eiwitten en koolhydraten niet onafhankelijk kunnen reguleren. Bovendien, omdat we chemisch ongedefinieerde diëten gebruiken, is het moeilijk om controle te houden over de concentraties voedingsstoffen die de patronen van larvale voeding direct kunnen beïnvloeden. Om deze problemen op te lossen, of om hits op een vooronderzoek te bevestigen en verder te ontleden, zou de experimenteerder de mogelijkheid willen overwegen om een nauwkeurige en gecontroleerde experimentele voedingscontext vast te stellen, door gedefinieerde synthetische (holidische) mediums10 te gebruiken en voedselkeuzetesten op te zetten zoals eerder beschreven30. Bij het gebruik van een protocol met thermogenetische neuronale modulatie is het belangrijk om te overwegen dat de noodzakelijke temperatuurverschuivingen de gedragsoutput van dieren direct kunnen beïnvloeden. Een aanvullend gebruik van optogenetische benaderingen zou interessant zijn om te controleren op temperatuurgeïnduceerde valse positieven, maar het gebruik van optogenetica in de context van larvale voedingstesten is technisch uitdagend, omdat voedende larven het grootste deel van de tijd in het voedselsubstraat doorbrengen.
Toch kunnen verschillende sterke punten van onze experimentele aanpak worden opgesomd. De eenvoud en relatief hoge doorvoer van onze methode maken het mogelijk om de voedselinname voor verschillende genotypen te kwantificeren wanneer ze worden blootgesteld aan verschillende voedingsomstandigheden. Voedingsgedrag in het larvale stadium is gemakkelijker kwantificeerbaar dan bij volwassen vliegen, waardoor het genereren van betere functionele uitlezingen mogelijk is. Het is ook minder uitdagend om voedingstesten vast te stellen die lijken op de natuurlijke omgeving bij larven dan voor volwassenen, zoals eerder is besproken31. Bovendien, in vergelijking met andere eerder vastgestelde methoden om het voeren in larven te kwantificeren, namelijk die gebaseerd op het handmatig tellen van het aantal mondhaakcontracties gedurende een bepaalde periode32, maakt onze colorimetrische methode genetische screeningstudies op grotere schaal mogelijk. Sommige andere methoden zijn eenvoudigweg gebaseerd op het scoren van het aandeel larven met geverfd voedsel in hun darmen , waardoor een nauwkeurige kwantificering van voedselinnameniveaus33,34niet mogelijk is . Met betrekking tot de neurogenetische controle van de neuronale functie zorgt het feit dat TRPA1-transgeen bij 18 °C inactief is ervoor dat neuronale activiteit niet wordt beïnvloed tijdens de ontwikkeling van larven. Dit zorgt ervoor dat de experimentele neuronale activering uitsluitend tijdens de voedertest wordt uitgevoerd en niet tijdens de larvale ontwikkeling. Daarnaast willen we nog een keer vermelden dat ons protocol eenvoudig kan worden aangepast aan specifieke behoeften en interesses van de experimenteerder. De onderdrukking van de neuronale functie, in plaats van activering, kan bijvoorbeeld gemakkelijk worden verkregen door de dTRPA1 te vervangen door een UAS-lijn die codeert voor de temperatuurgevoelige neuronale geluiddemper ShibireTS20. Ook als de voedingsniveaus van de experimentele larven erg laag zijn, waardoor het moeilijk is om de voedselinname te kwantificeren, is het mogelijk om een extra stap van 30 minuten larvale verhongering uit te voeren vóór de voedingstest (vóór de stappen in sectie 4 van het protocol), zoals eerder beschreven15. Deze voedseltekortstap kan bijzonder interessant zijn als u modulatoren van hongergestuurd gedrag onderzoekt. Ten slotte werd in eerdere studies, met behulp van kwantitatieve colorimetrische methoden, aangetoond dat het etiketteren van voedsel met blauwkleurstof geen invloed heeft op het voeren12. Niettemin zijn wij van mening dat het gebruik van complementaire, nauwkeurigere en gevoeligere methoden, zoals de radiolabeling van het levensmiddel12, in meer gevorderde stadia van een studie, gericht op het bevestigen of verder ontleden van hits die tijdens de voorbereidende stadia zijn gevonden, een goede aanvulling op onze methode zou zijn en door de experimenteerder zou moeten worden overwogen. Om al deze redenen geloven we in de aantrekkelijkheid van onze methoden om genetische schermen (vooral primaire schermen) uit te voeren, gericht op het identificeren van neuronale populaties die betrokken zijn bij het samenstellen van neuronale circuits die coderen voor voedingsgedrag.
Als laatste opmerking willen we vermelden dat duizenden larvale Gal4-lijnen die zijn gevestigd in Janelia Research Campus openbaar beschikbaar zijn, in bloomington Drosophila Stock Center en een grote hoeveelheid informatie over larvale26 en volwassen19 CNS-expressiepatronen is ook openbaar toegankelijk in de FlyLight Image Database (http://www.janelia.org/gal4-gen1). Deze bronnen maken het mogelijk om putatieve structuur-functie neuronale kaarten uit te werken van de neuronen die het voedingsgedrag in Drosophila-larven reguleren. Dit is mogelijk door de fenotypische informatie gegenereerd in neuronale schermen te integreren met de expressiepatronen van de gebruikte stuurprogramma's. Wij geloven dat onze methoden een geldige benadering vormen om voorlopige neuronale kaarten te genereren voor voedingsgedrag geassocieerd met macronutriëntenbalancering in de Drosophila-hersenen.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs hebben niets bekend te maken.
Acknowledgments
Wij willen het Instituto Gulbenkian de Ciência (IGC) bedanken voor het verlenen van toegang tot een deel van de experimentele apparatuur die in dit protocol wordt beschreven. Dit werk werd ondersteund door portugese Stichting voor Wetenschap en Technologie (FCT), LISBOA-01-0145-FEDER-007660, PTDC/NEU- NMC/2459/2014, IF/00697/2014 en La Caixa HR17-00595 aan PMD en door een Australian Research Council Future Fellowship (FT170100259).
Materials
| Name | Company | Catalog Number | Comments |
| 1.5 mL microtubes | Sarstedt AG & Co. | 72.690.001 | |
| 10xPBS | Nytech | MB18201 | |
| 2.0 mL microtubes | Sarstedt AG & Co. | 72.695.500 | |
| 60 mm petri dishes | Greiner Bio-one, Austria | 628161 | |
| 96 well microplates | Santa Cruz Biotechnology | SC-204453 | |
| Agar | Pró-vida, Portugal | ||
| Bench cooler | Nalgene, USA | Labtop Cooler 5115-0032 | |
| Blue food dye | Rayner, Billingshurst, UK | ||
| Cell disruption media | Scientific Industries, Inc. | 888-850-6208 | (0.5 mm glass beads) |
| Dish weight boats | Santa Cruz Biotechnology | SC-201606 | |
| Embryo collection cage for 60 mm petri dishes | Flystuff, Scientific Laboratory Supplies, UK | FLY1212 (59-100) | |
| Featherweight forceps | BioQuip Products, USA | 4750 | |
| Fly food for stocks maintenance | 1 L food contains: 10 g Agar, 100 g Yeast Extract, 50 g Sucrose, 30 mL Nipagin, 3 mL propionic acid | ||
| Forceps #5 | Dumont | 0108-5-PS | Standard tips, INOX, 11cm |
| Incubator | LMS Ltd, UK | Series 2, Model 230 | For thermogenetic feeding assay (30?C) |
| Incubator | Percival Scientific, USA | DR36NL | To stage larvae (19?C) |
| Janelia lines | Janelia Research Campus | Detailed information in Table 2 | |
| Macronutrient balancing diets | Composition and nutritional information in Figure 1 | ||
| Methanol | VWR | CAS number: 67-56-1 | |
| Nipagin (Methyl 4-hydroxybenzoate) | Sigma-Aldrich | H5501 | |
| Nitrile gloves | VWR, USA | ||
| Refrigerated centrifuge | Eppendorf, Germany | 5804 R / Serial number: 5805CI364293 | |
| Rubin Gal4 ines | Janelia Research Campus | Stoks available at Bloomington Drosophila Stock Center | |
| ShibireTS UAS line | Bloomington Drosophila Stock Center | BDSC number: 66600 | Provided by Carlos Ribeiro Group |
| Soft brushes | For sorting anaesthetised fruit flies | ||
| Spectrophotometer plate reader | Thermo Fisher Scientific | Multiskan Go 51119300 | |
| Stereo microscope | Nikon | 1016625 | |
| Sucrose | Sidul, Portugal | ||
| Third-instar larvae (L3) rearing diet | Composition and nutritional information in Figure 1 | ||
| Timer | |||
| Tissue lyzer / bead beater | MP Biomedicals, USA | FastPrep-24 6004500 | |
| TRPA1 UAS line | Bloomington Drosophila Stock Center | BDSC number: 26264 | Expresses TrpA1 under UAS control; may be used to activate neurons experimentally at 25 ?C |
| Water bath | Sheldon Manufacturing Inc., USA | W20M-2 / 03068308 / 9021195 | |
| Yeast extract | Pró-vida, Portugal | 51% Protein, 15% Carbohydrate |
References
- Raubenheimer, D. Nature of nutrition - a unifying framework from animal adaptation to human. , (2012).
- Carvahlo, M. J. a, Mirth, C. K. Coordinating morphology with behavior during development: an integrative approach from a fly perspective. Frontiers in Ecology and Evolution. , (2015).
- Steck, K., et al. Internal amino acid state modulates yeast taste neurons to support protein homeostasis in Drosophila. Elife. 7, 31625 (2018).
- Itskov, P. M., Ribeiro, C. The dilemmas of the gourmet fly: the molecular and neuronal mechanisms of feeding and nutrient decision making in Drosophila. Frontiers in Neuroscience. 7, 12 (2013).
- Bjordal, M., Arquier, N., Kniazeff, J., Pin, J. P., Leopold, P. Sensing of amino acids in a dopaminergic circuitry promotes rejection of an incomplete diet in Drosophila. Cell. 156 (3), 510-521 (2014).
- Grandison, R. C., Piper, M. D., Partridge, L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature. 462 (7276), 1061-1064 (2009).
- Lee, K. P., et al. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proceedings of the National Academy of Sciences of the United States of America. 105 (7), 2498-2503 (2008).
- Levine, M. E., et al. Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population. Cell Metabolism. 19 (3), 407-417 (2014).
- Solon-Biet, S. M., et al. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metabolism. 19 (3), 418-430 (2014).
- Piper, M. D., et al. A holidic medium for Drosophila melanogaster. Nature Methods. 11 (1), 100-105 (2014).
- Jones, W. D. The expanding reach of the GAL4/UAS system into the behavioral neurobiology of Drosophila. BMB Reports. 42 (11), 705-712 (2009).
- Deshpande, S. A., et al. Quantifying Drosophila food intake: comparative analysis of current methodology. Nature Methods. 11 (5), 535-540 (2014).
- Neckameyer, W. S. A trophic role for serotonin in the development of a simple feeding circuit. Developmental Neuroscience. 32 (3), 217-237 (2010).
- Gasque, G., Conway, S., Huang, J., Rao, Y., Vosshall, L. B. Small molecule drug screening in Drosophila identifies the 5HT2A receptor as a feeding modulation target. Scientific Reports. 3, (2013).
- Schoofs, A., et al. Selection of motor programs for suppressing food intake and inducing locomotion in the Drosophila brain. PLoS Biology. 12 (6), 1001893 (2014).
- Pocas, G. M., Crosbie, A. E., Mirth, C. K. When does diet matter? The roles of larval and adult nutrition in regulating adult size traits in Drosophila melanogaster. Journal of Insect Physiology. , 104051 (2020).
- Hamada, F. N., et al. An internal thermal sensor controlling temperature preference in Drosophila. Nature. 454 (7201), 217-220 (2008).
- Pfeiffer, B. D., et al. Tools for neuroanatomy and neurogenetics in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 105 (28), 9715-9720 (2008).
- Jenett, A., et al. A GAL4-driver line resource for Drosophila neurobiology. Cell Reports. 2 (4), 991-1001 (2012).
- Kitamoto, T. Conditional modification of behavior in Drosophila by targeted expression of a temperature-sensitive shibire allele in defined neurons. Journal of Neurobiology. 47 (2), 81-92 (2001).
- Brand, A. H., Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118 (2), 401-415 (1993).
- Shirangi, T. R., Stern, D. L., Truman, J. W. Motor control of Drosophila courtship song. Cell Reports. 5 (3), 678-686 (2013).
- Mirth, C. M. J. Food intake and food choice are altered by the developmental transition at critical weight in Drosophila melanogaster. Animal Behaviour. 126, 195-208 (2017).
- Simpson, S. J., Raubenheimer, D.
- Raubenheimer, D., Simpson, S. J. Integrative models of nutrient balancing: application to insects and vertebrates. Nutrition Research Reviews. 10 (1), 151-179 (1997).
- Li, H. H., et al. A GAL4 driver resource for developmental and behavioral studies on the larval CNS of Drosophila. Cell Reports. 8 (3), 897-908 (2014).
- Bhatt, P. K., Neckameyer, W. S. Functional analysis of the larval feeding circuit in Drosophila. Journal of Visualized Experiments. (81), e51062 (2013).
- Wong, R., Piper, M. D. W., Blanc, E., Partridge, L. Pitfalls of measuring feeding rate in the fruit fly Drosophila melanogaster. Nature Methods. 5 (3), 214-215 (2008).
- Almeida-Carvalho, M. J., et al. The Ol1mpiad: concordance of behavioural faculties of stage 1 and stage 3 Drosophila larvae. Journal of Experimental Biology. 220, Pt 13 2452-2475 (2017).
- Rodrigues, M. A., et al. Drosophila melanogaster larvae make nutritional choices that minimize developmental time. Journal of Insect Physiology. 81, 69-80 (2015).
- Wong, R., Piper, M. D., Wertheim, B., Partridge, L.
- Wu, Q., et al. Developmental control of foraging and social behavior by the Drosophila neuropeptide Y-like system. Neuron. 39 (1), 147-161 (2003).
- Wu, Q., Zhang, Y., Xu, J., Shen, P. Regulation of hunger-driven behaviors by neural ribosomal S6 kinase in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 102 (37), 13289-13294 (2005).
- Lingo, P. R., Zhao, Z., Shen, P. Co-regulation of cold-resistant food acquisition by insulin- and neuropeptide Y-like systems in Drosophila melanogaster. Neuroscience. 148 (2), 371-374 (2007).
Tags
Neurowetenschappen Drosophila larven thermogenetisch neuronaal scherm larvaal voedingsgedrag macronutriëntenbalancering eiwit koolhydraten voedselinnametest voedselkleurstof colorimetrische kwantificeringErratum
Formal Correction: Erratum: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae
Posted by JoVE Editors on 10/06/2020.
Citeable Link.
An erratum was issued for: Quantification of Macronutrients Intake in a Thermogenetic Neuronal Screen using Drosophila Larvae. A figure was updated.
Figure 1 was updated from:
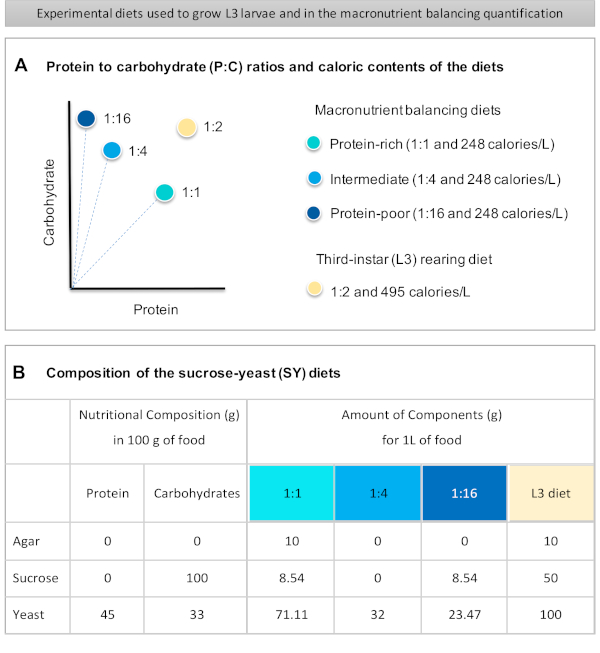
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
to:
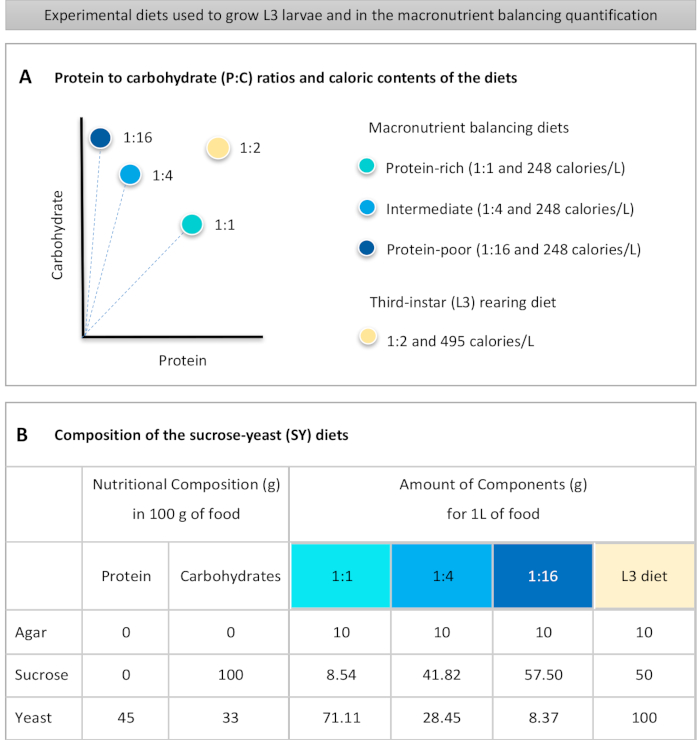
Figure 1: The sucrose-yeast (SY) diets used in our protocol. (A) The blue dots represent the isocaloric (248 calories/L) macronutrient balancing diets used in the feeding assay, which differ in the protein to carbohydrate (P:C) ratios: 1:1, 1:4 and 1:16. The beige dot represents the diet used to rear the experimental third-instar larvae (L3), which contained a P:C ratio of 1:2 and a caloric density of 495 calories/L. (B) Detailed composition and nutritional information of the sucrose-yeast (SY) based diets. The components are the same for all the diets: agar, sucrose and yeast. The amount in grams of the components needed to prepare 1 L of diet is shown. Note that 1% (v/v) of blue dye must be added to the macronutrient balancing diets and to the L3 rearing diet nipagin and propionic acid solutions must be added to a final concentration (v/v) of 3% and 0.3%, respectively. Please click here to view a larger version of this figure.
