

Summary
세포주기 단계의 진행과 속도론의 추적 미묘한 변화는 Propidium 요오드화물의를 통해 BrdU 및 전체 게놈의 DNA의 염색법과 핵산의 대사 라벨링의 조합을 사용하여 수행할 수 있습니다. 이 방법은이를 차례로 세포주기의 진행에 영향을 미치는가 아닌 특정의 DNA 손상의 도입을 방지, 자전거 전지의 화학 동기화의 필요성을 방지합니다.
Abstract
세포주기의 여러 단계를 거치 개시 및 이후 진행 정밀 제어는 세포를 proliferating의 파라마운트 중요하다. 세포주기 부문의 성장과 주요 세포주기의 구성 요소 복제 및 규제 완화가 carcinogenesis 1,2의 precipitating 사건에 연루되었을의 중요한 부분입니다. 안티 - 암 치료의 분자 에이전트는 자주 셀 사이클 부문 3 규제와 조정을 담당 생물 학적 경로를 타겟. 세포주기 운동은 세포 종류에 따라 다릅니다하는 경향이 있지만, 세포주기의 4 단계 사이 세포의 분포 mitogen과 성장 인자의 표현의 일관된 패턴으로 인해 특정 세포주 내에서 다소 일관성이다. Genotoxic 이벤트 및 기타 세포의 스트레스는 insti 가능하도록하기 위해서 체포 또는 특정 세포주기 단계에서 일시 정지가 발생, 세포주기 진행의 임시 블록이 발생할 수적절한 응답 메커니즘 gation.
실험적으로 자신의 세포주기의 진행 단계에 대한 참조가있는 세포 인구의 동작을 관찰하는 능력은 세포 생물학의 중요한 사전이다. 같은 mitotic와 같은 일반적인 절차는 세포주기 4-6의 특정 단계에서 세포를 분리하는 데 사용됩니다, 차등 원심 분리 또는 흐름 cytometry 기반 정렬을 뿌리 치다. 이러한 fractionated, 세포주기의 위상 강화 인구 그런 다음 실험적인 치료를 받게됩니다. 분리된 분수의 항복 순도 및 생존 능력은 종종 이러한 물리적인 분리 방법을 사용 위험 할 수 있습니다. 뿐만 아니라, 세포 인구의 분리 및 fractionated 세포가 선택한 셀 사이클 단계에서 발전할 수 의하여 실험적 치료의 시작 사이의 시간 경과는 성공적인 구현과 실험의 해석에 상당한 어려움을 초래할 수 있습니다.
성 이밖에 접근udy 세포주기 단계는 세포를 동기화하는 화학 물질의 사용을 포함합니다. 각각의 세포주기의 단계에 대한 주요 신진 대사 과정의 화학 억제제와 세포 치료는 다음 단계로 세포주기의 진행을 차단에 유용합니다. 예를 들어,하여 G1 / S 시점에서 ribonucleotide 환원 효소 억제제 hydroxyurea 중단 전지 deoxynucleotides, DNA의 빌딩 블록의 공급을 제한. 다른 주목할만한 화학 물질이 aphidicolin과 치료를 포함, mitotic 스핀들 형성에 방해가 둘 다 G1 연금을위한 중합 효소 알파 억제제, 콜히친 그리고 nocodazole과 치료, M 단계에서 세포를 중단하고 마지막으로, DNA 사슬 종결 5 fluorodeoxyridine과 치료 시작 S 위상 체포 7-9. 이러한 화학 물질로 치료는 특정 단계에서 세포의 전체 인구를 동기화의 효과적인 수단입니다. 화학 물질의 제거를 통해 세포 한마음으로 세포주기를 다시 가입. 테스트 에이전트 다음 릴리스의 치료세포주기를 차단하는 화학 물질로부터 elicited 마약 응답 유니폼, 세포주기 단계 - 특정 인구에서임을 보장합니다. 그러나, 화학 synchronizers의 많은 이래 도전하고 다양한 반응 경로 (테스트 에이전트 대 synchronizers까지)의 참여를 차별 괴롭히고, genotoxic 화합물을 알려져 있습니다.
여기에서 우리는 분열과 딸 세포의 분리에 이르기까지, DNA 복제 단계에서 자신의 진행을 통해 적극적으로 사이클링 세포의 subpopulation에 따라위한 신진 대사 라벨링 방법을 설명합니다. 유동세포계측법의 부량 결합이 프로토콜의 부재에서 세포주기의 운동 진행 중 측정 가능 중 기계적 또는 화학적으로 유도된 세포는 일반적으로 다른 세포주기의 동기화 방법 10와 관련된 강조합니다. 다음 섹션에서 우리는뿐만 아니라 생명 의학 연구 애플 리케이션의 일부로서, 방법론을 설명합니다.
Protocol
1. 셀 준비

- 약 60% confluency의 밀도를 달성하기위한 플레이트 세포. 세포는 콜렉션의 시간에 로그 단계에 있어야합니다. mcf7 세포의 경우이은 5 × 10 5 세포 / 적절한 매체 10cm 접시에 퍼뜨리고에 의해 수행됩니다. HT29 및 LS180 세포는 6 × 10 5 세포 / 6cm 판에 놓는됩니다. 우리는 DMEM 미디어 10 % FBS 및 1X 페니실린 / 스트렙토 마이신과 함께 보충 사용됩니다. 에서 올바른 긍정과 부정적인 컨트롤을위한 종자 접시해야테스트 샘플에 추가. 그들은 다음과 같습니다 :
| 나 | 긍정적인 제어 | 전용 BrdU |
| II | 긍정적인 제어 | PI 전용 |
| III | 부정적인 제어 | BrdU 부정, PI 부정적 |
(그것은 이와 프로토콜에 소개된 것과 같이 여러 timepoints와 예비 실험이 미래의 컬렉션을 수행하는 동안 기간의 범위를 좁힐하는 데 사용할 수있는 것을인지해야합니다.)
- 도금 세포는 37 incubated있다 ° C에서 5 % 24-48 H에 대한 CO 2,이를 통해 세포가 회복하고 연결할 수 있도록합니다.
- bromodeoxyuridine (BrdU)로 라벨 세포를 펄스 위해 DMEM는 WI를 대체됩니다10 μm의 BrdU을 포함 일까지 신선 미디어. 세포 ° C, 5 % CO 2가 DNA로 BrdU의 설립을 할 수 있도록 37에서 1 H 위해 incubated됩니다. 부정적인 제어 역할을 치료 한 접시를 두십시오.
- 펄스 라벨 미디어가 제거되고 세포는 1X PBS로 간단히 씻어서있다.
- 신선한 성장 매체는 추가됩니다 (마이너스 BrdU)과 세포 ° C에서 5 % CO 2 수확에 적합한 timepoint에 도달할 때까지 37 품어 계속해서 사용할 수 있습니다.
2. 수확 및 고정
- 1.3 단계에서 치료 세포는 제로 timepoint 간주됩니다. 이 샘플은 BrdU 치료 직후 수집되는 한 H timepoint 함께 수확 할 수 있습니다.
- 세포를 수확하기 위해 미디어가 제거되고 플레이트는 1X PBS로 한 번 씻어서있다.
- 셀은 5 분 먹어 볼래요 위해 1,500 rpm으로 원심 분리하여 trypsinized culturing 매체에서 수집하고 이후 pelleted있다ernatant은 삭제됩니다.
- 5 ML 얼음 차가운 1X PBS에 1-10 X 10 6 세포를 씻어. 5 분 뜨는 동안 1,500 rpm으로 원심은 삭제됩니다.
- 세포 펠렛 그런 다음 얼음처럼 차가운 PBS / 1% FBS를 100 μl에 resuspended있다. 셀 clumping을 예방 PBS 에이즈에 1퍼센트 FBS의 추가.
- -20 ° C, 세포를 해결하기 위해 70 % 에탄올 5 ML에 드롭하여 이러한 세포 드롭을 추가합니다.
- 4 ° C에서 하룻밤 사이에 30 분 또는 상점을 위해 얼음에 품어. 이것은 다양한 timepoints에서 샘플 수집을 중지되는 이상적인 단계입니다. 샘플은 그들이 프로토콜의 나머지 단계를 동시에 처리할 수 있도록 며칠 동안 에탄올로 남아있을 수 있습니다.
3. BrdU과 PI 염색법
- 1,500 rpm으로 5 분 원심 분리하여 펠렛 세포.
- 정착액을 제거했지만 ~ 50 μl vortexing하여 펠렛을 풀어야하는 둡니다.
- DNA를 변성하기 위해서 천천히 2N HCL / 트리톤 X-100의 1 ML 추가vortexing하면서 dropwise. 30 분 동안 상온에서 샘플을 품어.
- 1,500 rpm으로 5 분 동안 샘플을 centrifuging하여 펠렛 세포. 기음과 뜨는 버리고.
- 변성 단계를 무력화하기 위해 0.1M 나트륨 테트라 보 레이트, 산도 8.5의 1 ML에서 세포 펠렛을 Resuspend.
- 1,500 rpm으로 5 분 동안 세포를 원심 분리기. 기음과 뜨는 버리고.
- BrdU의 염색법 믹스의 75 μl (50 μl 0.5 % 십대 초반 1분의 20 % BSA / PBS + 20 μl FITC 복합 백신 BrdU + 5 μl 10 밀리그램 / ML RNase)에 세포 펠렛을 Resuspend.
- 빛으로부터 보호 45 분 동안 상온에서 품어.
- 펠렛 세포로, 1,500 rpm으로 5 분 동안 샘플을 원심 분리기. 기음과 뜨는 버리고.
- PBS는 5 μg / ML의 propidium의 요오드화물을 포함하는 1 ML에서 세포 펠렛을 Resuspend.
4. 유동세포계측법
- 세포를 분석하기 위해 유동 cytometer은 488 nm의 레이저 및 장착된적절한 필터가 필요합니다. 같은 CellQuest 같은 분석 소프트웨어는 아래에 설명된 플롯을 만들 필요가 있습니다.
- 흐름 cytometer를 통해 샘플을 실행 시킬때, 세포의 적절한 크기 분포 (그림 1A)을 보장하기 위해 전방 산란 (SSC-H) 대 측면 산란 (FSC-H) 플롯을 만듭니다. 이러한 매개 변수 중 모두 선형 스케일 꾸몄다됩니다.
- 동시에 FL3-H (PI는 얼룩) 대 FL2-W (그림 1B)의 음모를 볼 수 있습니다. 이 플롯은 세포주기의 정규 분포 패턴 내에있는 세포의 분율을 분리하는 게이트 (R3)를 만드는 데 사용됩니다. 일반적으로 이러한 분포 패턴은 각각 세포주기의 G1과 G2 단계를 대표하는 사립 탐정 염색법의 2N와 4N DNA 컨텐츠의 세포는 두 클러스터이 특징입니다. 이 두 클러스터 사이에 위치한 세포의 문자열은 세포주기의 S 단계에서 발생하는 지속적인 DNA 복제의 대표이다.
- 플롯의 displayi 만들기NG y 축은와 X-축 (그림 1C)에 PI (선형 계획)의 FL1-H와 단계 4.3 (FITC, 로그 음모)에서 문이 세포 (R3). 이 줄거리는 분석을위한 데이터를 수집하는 다양한 1.1에서 설명한 컨트롤뿐 아니라 사용하여 나머지 매개 변수를 설정하는 데 사용됩니다.
- cytometer쪽으로 PI 유일한 스테인드 샘플을 놓습니다. X 축에 ~ 200 G1을 배치하는 게인을 조정합니다. 이것은 얼룩 PI의 히스토그램 플롯의 시각화하기 쉬울 것입니다.
- 두 번째 제어 흐름 cytometer에 BrdU 전용 샘플을 실행하는 것입니다. 세포의 2 인구 (BrdU 양성 대 BrdU 음수)가 음모에 표시되도록 게인을 조정합니다. 이상적으로, BrdU 부정적인 세포는 단 10 -1 아래에 표시하기 위해 위치가 결정됩니다.
- 최종 제어는 BrdU / PI 부정적인 샘플입니다. 세포가 위쪽이나 오른쪽 quadrants의에 표시되지 않도록하려면이 부정적인 제어를 실행합니다.
- 다양한 플롯의 매개 변수가 설정되고 나면유동 cytometer는 BrdU과 PI 스테인드 세포 처리를 위해 보정하고 준비하고 있습니다. 올바른 PI 분율 (4.3 단계를 참조)에 문이있는 10,000 전지의 최소 모음에 따라 읽어야합니다.
- 수집된 데이터를 분석하려면 이러한 FlowJo 또는 FacsDiva과 같은 소프트웨어가 활용된다. 위에서 언급한 각각의 플롯은이 프로그램에서 재현하고 정량 및 통계 분석을 수행할 수 있습니다.
- 분석의 종점은 몇 연속적인 단계를 포함한다. PI 대 FL2-W 줄거리에서 긍정적인 PI와 같은 문이 세포와 FITC 대 PI의 줄거리를 생성하는 것은 하나가 자전거 인구를 구별할 수 있습니다. 두 번째 게이트는 BrdU 양성 인구를 분리하여이 음모에서 생성됩니다. X 축에 대한 PI와 히스토그램 플롯에서이 특유의 인구를 표현하는 한 세포주기 진행의 시간 과정을 추적할 수 있습니다. 이것은 G1 또는 시간의 함수로서 G2 콘텐츠로 BrdU 양성 세포의 수를하려하여 더욱 시각이 될 수 있습니다.
5. 대표 결과
일반적으로 propidium 요오드화물의 물들일 자전거 전지는 각각 2N와 4N DNA 내용을 포함하는 세포 (그림 2)에 해당하는 G1 및 G2의 독특한 봉우리을 보유하고 있습니다. BrdU 펄스 라벨 적극적으로 DNA를 (즉, S 단계)를 합성하고 세포의 하위 인구의 선택적 라벨링을 허용합니다. 짧게 BrdU 시약의 제거 후, 모든 레이블 세포는 S 단계 (그림 3)에 있습니다. 그들은 세포주기의 후속 단계 통과로 짧은 펄스로 라벨을 제한함으로써 한 많은 시간을 포인트 전반에 걸쳐 세포의이 이제 별개의 하위 인구를 따릅니다 수 있습니다. 이것은 G1의 독특한 부족과 G2 봉우리 그림 3의 1 H 시간 지점에서 시각이 될 수 있습니다.
추가 정보는 BrdU 라벨 단계에서 파생된 수 있습니다. 적극적으로 나누어 세포의 비율은 동부 표준시는 측정하지만 수있다뿐만 아니라샘플 두개 사이에 세포주기의 단계 분배 imate은 잘 결정하실 수 있습니다. 그들이 G2로 순환, 발전에 지속적으로 BrdU 시약의 제거에 따라 균등하게 간격 간격으로 세포를 수집하여 세포 추적할 수 있으며, 궁극적으로 결국 G1과 BrdU-긍정 딸 세포로 신흥, 원래 세포의 세포 분열까지 이동 DNA 내용 (그림 4). 세포주기의 위상 부량 및 운동 분석의 예제는 그림 5에서 제공됩니다.

그림 1. 대표 mcf7 세포 인구 (A) 앞으로 산포 대 사이드 분산형 플롯. 대표 mcf7 세포 인구 (B) PI 대 폭 (FL-W) 플롯. 게이팅은 최종 분석 (R3)에서 셀 doublets를 제외하도록 표시됩니다. 그들은 레이저 빔을 및 theref 통과 시간이 오래 걸릴으로 휴대 doublets는 단일 셀보다 큰 펄스 너비를해야합니다광석은 분석에서 제외시킬 수 있습니다. 대표 mcf7 세포 인구 (C) PI 대 FITC (BrdU) 플롯. 게이팅는 FITC (BrdU) 긍정적인 세포 (R4)를 포함하도록 표시됩니다.

그림 2. 정상적으로 순환 세포가 전체 인구의 대표 히스토그램 계획.

그림 3. FITC (BrdU) - 긍정적인 전지 게이팅 후 BrdU 펄스의 제거 후 1 시간을 수집되어 세포의 히스토그램 플롯. 라벨의 제거에 따라 조기 timepoint에서 BrdU 양성 세포는 DNA를 합성하는 동안 세포의 성공적인 라벨을 확인, 세포주기의 S-단계에있는 세포와 일치하는 DNA의 컨텐츠를 표시하는 샘플에 해당하는 PI 프로파일을 보여줍니다.
 그건 = "그림 4"/>
그건 = "그림 4"/>
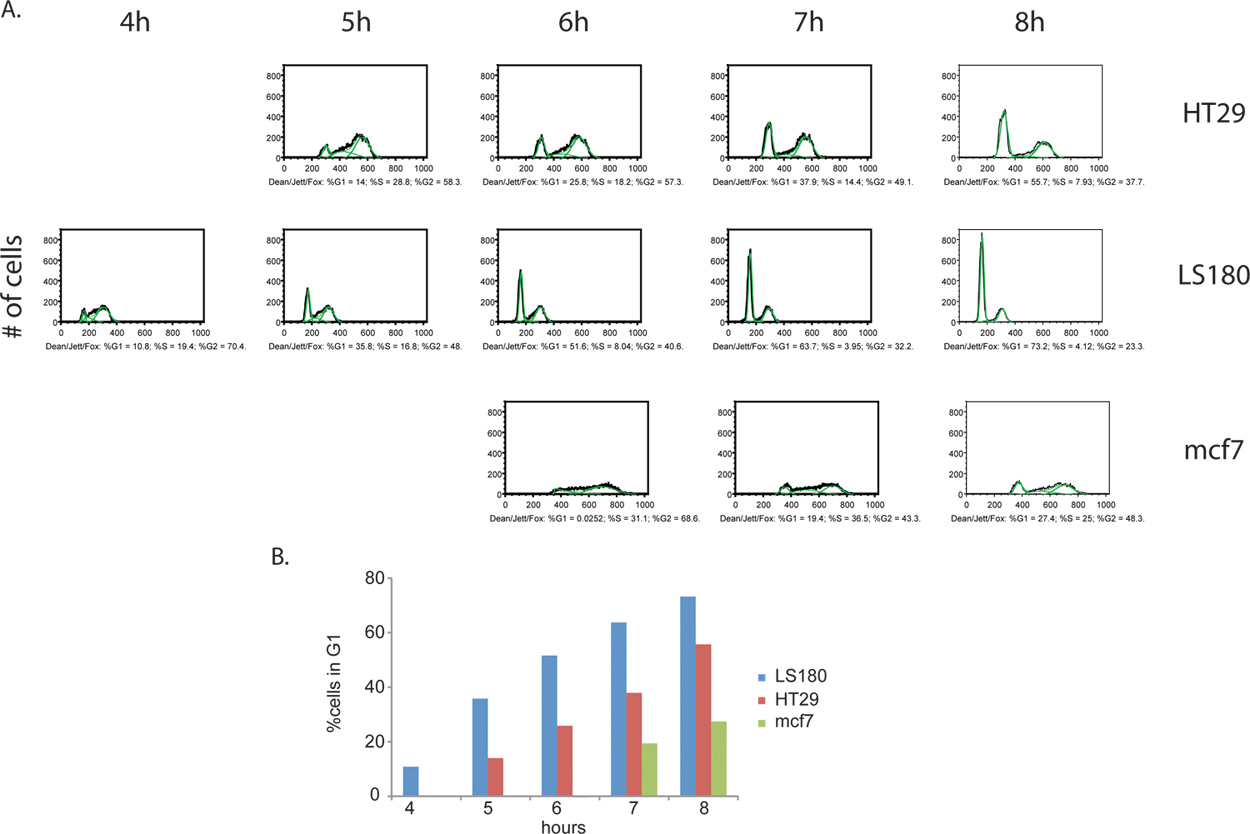
그림 4. 암 세포 라인, 대장암 암 HT29 () 및 LS180 (B)뿐만 아니라, 유방암 mcf7 (C) 사이의 세포주기 진행의 속도론의 비교. 전지는 BrdU 펄스의 제거에 따라, 8 시간 동안 매 시간을 수집했다. 본 실험에서는 대장암 세포 라인 LS180의 세포주기의 G2 단계를 통해 빠른 속도로 세포주기 진행의 명확한 프로파일을 관찰했다. 대장암 세포의 두 프로파일 사이에 운동을 비교, G1 피크의 출현이 정상이 부족 HT-29 세포주에 대한 해당 시점에 비해, LS180 세포에서 T = 4h 이후 BrdU에 분명하다. 두 대장암 세포 라인 중 하나에 비해 mcf7 전지는 크게 줄어 속도로 돌고있다. 큰 그림을 보려면 여기를 누르십시오 .
다시 5 "/>
그림 5. BrdU-라벨이 암 세포 () 양적 세포주기의 위상 분석. 딘 / 제트 / 여우 알고리즘 HT29, LS180과 mcf7 (녹색에서 그림)에 적용되었다. 각 샘플의 결과로 세포주기의 위상 배포판은 각 단계에 대한 % 총 BrdU-양성 세포로 표현됩니다. 이전 시점 점 중 일부에 대한 해석은 잘못된 결과를 생산으로 전용 각 세포주 시간 지점을 선택가 표시됩니다. 이러한 이전 시점 지점에서 모든 BrdU 양성 세포는 S-단계에 따라서 알고리즘 응용 프로그램에 필요한 독특한 G1과 G2 봉우리를 부족합니다. (B) 암 세포 Histograms은 세포주기의 G2 / M 단계를 통해 진행 속도론을 보여주고 있습니다. G2 / M 단계를 통해 진행이 HT29 및 mcf7 다음 LS180 세포를위한 가장 빠른 것입니다. 더 큰 그림을 보려면 여기를 누르십시오 .
JPG "고도 ="그림 S1 "/>
그림 S1. 유동세포계측법 제어합니다. mcf7 세포는 세포가 PI이나 BrdU의 스테인드도 있습니다 () 부정적인 컨트롤로 표시됩니다. (B) PI에만 묻다. (C) BrdU-FITC는 샘플을 분류.

PI 대 FITC (BrdU)의 형광 방출 스펙트럼 사이의 관계를 보여주는 S2. 도식 일러스트 그림. 형광 채널 1 (FL1, FITC) 및 형광 채널 3 (FL3, PI 용)에 대한 스펙트럼 회수 창이 해당 상자에 표시됩니다. FL1과 FL3 검출 사이 형광단 스펙트럼 중첩이 없습니다. PI와 FITC-BrdU의 실험 데이터를 공동으로 표시된 셀이 각각 FL1/FL3 채널에서 수집한 때 결국, 형광 보정이 필요하지 않습니다. 형광 spectrums는 BD BioSciences 웹사이트에서 얻은되었습니다대상 = "_blank"> http://www.bdbiosciences.com/research/multicolor/spectrum_viewer. 큰 그림을 보려면 여기를 누르십시오 .
Subscription Required. Please recommend JoVE to your librarian.
Discussion
BrdU의 설립과 함께 유동세포계측법를 결합함으로써, 우리는 세포주기의 속도론을 연구하는 데 필요한 도구를 가지고. thymidine의 아날로그로 작동하려면 BrdU의 독특한 속성은 자전거 세포의 DNA 컨텐츠 부량을 허용거야. 세포주기의 합성 단계에서 성장하는 딸의 DNA 가닥으로 BrdU의 법인이 하나가 G2에서 성장의 시대에 궁극적으로 세포 분열로, S 단계에서 DNA의 복제를 통해 자전거 세포의 subpopulation를 수행하는 것이 허용 뭔지 . BrdU 레이블은 신호가 배경 수준으로 감소 정도 세포 분열에 의해 희석되는 시간의 어느 시점까지 딸 BrdU-양성 세포가 계속 다음 수 있습니다.
총 DNA의 내용을 확인하기 위해 propidium 요오드화물을 사용하여, 우리는 S → G2 / M → G1에서 세포주기의 진행을 수행 할 수 있습니다. 세포의 순환 율을 추적할 수있는 능력은 특히 구경이 많은 응용 프로그램에서 중요하고 유용소설 화합물 11 세포주기의 단계 감도를 결정하는 세포주기의 특정의 DNA 손상 유도 약물 또는 약물 개발 과정과 관련된 R 사전 임상 연구. 신진 대사 레이블링 기법을 사용하는 가장 분명한 장점은 세포의 화학 동기화의 제거이다. 세포주기 정지를 유도 대부분의 화학 치료는 genotoxic 자체적으로 이러한 화학 물질의 면전에서 소설 화합물의 세포주기 감도 효과를 해부하는 것은 가능하지 않을 수 있습니다.
우리는 PI 공동 염색법이 BrdU의 신진 대사 라벨링 기법의 다양성을 보여주었다. 이 프로토콜을 사용, 우리는 성공적으로 세포주기의 G2 / M 단계를 통해 세포주기 진행의 증가가 효소 발현 10 강제 것으로 변형되어 telomerase - 부정적인 인간의 세포에서 관찰 될 수있다는 증명하고있다. 이 방법은 사이클 죽으로 세포의 동력학 측정에 강력한이지만여러 단계 헉, 한 제한은 전환시 타이밍 정보의 손실뿐만 아니라 G2와 M 단계 이내입니다. 세포가 M으로 G2를 통해 진행로서, 우리는 전적으로 DNA의 콘텐츠를 기반으로, 두 단계뿐만 아니라 구별할 수 없을뿐 아니라 하위 G1 인구. 이러한 분화가 필요한 어플 리케이션에서는 cyclins 또는 그의 표현 세포주기 12,13 중 원하는 단계와 관련된 다른 세포 단백질에 대한 항체와 공동 얼룩 BrdU 라벨이 세포에 잠재적으로 가능하다. 추가 최적화가 cyclin의 항체는 BrdU 레이블 및 immuno-염색법 프로토콜과 호환되도록하는 데 필요한 것입니다. 이상적으로, 적절히 세포주기의 다양한 단계를 차별화하기 위해 여러 cyclin의 항체는 하나의 샘플을 동시에 공동 라벨에 필요합니다. 우리 연구실은 세포를 구분하는 cyclin B와 phospho 히스톤 3 항체와 BrdU-펄스 표시된 셀의 이후 라벨에 노력하고 있습니다세포주기의 G2 대 M 위상 인치 PI와 FITC-BrdU 염색법과 더불어 cyclin의 항체와 공동 레이블은 크게 하드웨어와 데이터 분석 (형광 보상 문제) 관점 모두에서 유동세포계측법의 합병증을 증가시킵니다. 흐름 cytometers의 새로운 세대의 출현과 함께, 그들의 능력과 기능의 발전은 이러한 기술적인 문제를 완화해야합니다.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
우리는 공개 할게 없다.
Acknowledgments
우리는 FACS 분석과 도움을 UBC의 바이오 메디컬 연구 센터의 앤디 존슨 감사드립니다. 왕 연구소에서 암 연구 기금은 캐나다 암 협회 연구소 (운영 기금 # 019250)와 약학, UBC 학부의 연구 재투자 자금에서 제공됩니다. JMYW는 캐나다 연구 의자 및 건강 연구 경력 개발 프로그램에 대한 마이클 스미스 재단이 지원됩니다.
Materials
| Name | Company | Catalog Number | Comments |
| bromodeoxyuridine | BD Biosciences | 55089 | |
| propidium iodide | Sigma-Aldrich | 287075 | 1mg/ml stock |
| FITC anti-BrdU | BD Biosciences | 347583 | |
| sodium tetraborate | Fisher Scientific | S80172 | 0.1M, pH 8.5 |
| FACS Caliber | BD Biosciences |
References
- Musgrove, E. A., Caldon, C. E., Barraclough, J., Stone, A., Sutherland, R. L. Cyclin D as a therapeutic target in cancer. Nat. Rev. Cancer. 11, 558-572 (2011).
- Molchadsky, A., Rivlin, N., Brosh, R., Rotter, V., Sarig, R. p53 is balancing development, differentiation and de-differentiation to assure cancer prevention. Carcinogenesis. 31, 1501-1508 (2010).
- Dickson, M. A., Schwartz, G. K. Development of cell-cycle inhibitors for cancer therapy. Curr. Oncol. 16, 36-43 (2009).
- Banfalvi, G. Cell cycle synchronization of animal cells and nuclei by centrifugal elutriation. Nat. Protoc. 3, 663-673 (2008).
- van Opstal, A., Boonstra, J. Inhibitors of phosphatidylinositol 3-kinase activity prevent cell cycle progression and induce apoptosis at the M/G1 transition in CHO cells. Cell Mol. Life Sci. 63, 220-228 (2006).
- Gasnereau, I. Flow cytometry to sort mammalian cells in cytokinesis. Cytometry. A. 71, 1-7 (2007).
- Harper, J. V. Synchronization of cell populations in G1/S and G2/M phases of the cell cycle. Methods Mol. Biol. 296, 157-166 (2005).
- Pedrali-Noy, G. Synchronization of HeLa cell cultures by inhibition of DNA polymerase alpha with aphidicolin. Nucleic Acids Res. 8, 377-387 (1980).
- Merrill, G. F.
- Fleisig, H. B., Wong, J. M. Telomerase promotes efficient cell cycle kinetics and confers growth advantage to telomerase-negative transformed human cells. Oncogene. , (2011).
- Cai, D., Byth, K. F., Shapiro, G. I. AZ703, an imidazo[1,2-a]pyridine inhibitor of cyclin-dependent kinases 1 and 2, induces E2F-1-dependent apoptosis enhanced by depletion of cyclin-dependent kinase 9. Cancer Res. 66, 435-444 (2006).
- Pozarowski, P., Darzynkiewicz, Z. Analysis of cell cycle by flow cytometry. Methods Mol. Biol. 281, 301-311 (2004).
- Sherwood, S. W., Rush, D. F., Kung, A. L., Schimke, R. T. Cyclin B1 expression in HeLa S3 cells studied by flow cytometry. Exp. Cell Res. 211, 275-281 (1994).

{kind=link}
{kind=link}