

Summary
Xenopus embryonala ektoderm har blivit en attraktiv modell för studier av cellens polaritet. En analys beskrivs, där subcellulära distribution av fluorescerande proteiner bedöms i ektoderm celler. Detta protokoll kommer att hjälpa behandla frågor rörande fysisk kontroll av signalering.
Abstract
Cell polaritet är en grundläggande egenskap hos eukaryota celler som dynamiskt regleras av både endogena och exogena faktorer under fosterutvecklingen 1, 2. En av de signalvägar involverade i denna förordning är Wnt väg, som används många gånger under fosterutvecklingen och kritiska för mänskliga sjukdomar 3, 4, 5. Flera molekylära komponenter i denna väg reglera coordinately signalering i ett rumsligt fritt sätt, men de bakomliggande mekanismerna är inte helt klarlagda. Xenopus embryonala epitelceller är ett utmärkt system för att studera subcellulära lokalisering av olika signalsystem proteiner. Fluorescerande fusionsproteiner uttrycks i Xenopus embryon genom RNA mikroinjektion, är ektodermal explants förberedda och protein lokalisering utvärderas av epifluorescence. I denna experimentella protokoll beskriver vi hur subcellulära lokalisering av Diversin, en cytoplasmiska protein som har varit inblandad i signalering och cell polaritet beslutsamhet 6, 7 visualiseras i Xenopus ektodermal celler för att studera Wnt signaltransduktion 8. Coexpression av Wnt ligand eller en krulliga receptor förändrar fördelningen av Diversin smält med röda fluorescerande proteinet, RFP och rekryter till cellmembranet i ett polariserat sätt 8, 9. Detta ex vivo protokoll bör vara ett användbart komplement till in vitro-studier av odlade däggdjursceller, där rumsliga kontrollen av signalering skiljer sig från den intakta vävnaden och är mycket svårare att analysera.
Protocol
1. In vitro befruktning av Xenopus Ägg
- Skaffa ägg från kvinnliga grodor som injicerades med humant koriongonadotropin (400 enheter / groda) 12 timmar före försöket.
- Placera äggen i en liten mängd (0,5-1 ml) av en modifierad x Marcs Ringer-lösning (MMR) 10 till äggen och befruktar dem in vitro med en liten bit av dissekeras testiklarna. Efter 2-3 minuter, tillsätt 0,1 x MMR att täcka hela ytan av äggen. På 20 minuter är ägget gelé päls bort av 3% cystein - HCL (justerat till pH 8 med natriumhydroxid). Ägg är tvättas med 0,1 x MMR tre gånger och vänster i en kall inkubator (13 ° C) för injektionsvätskor.
- Befruktade äggen får utvecklas till 2-4 cell skede. För injektioner, är de embryon i lösningen innehåller 3% Ficoll, 0,5 x MMR.
2. RNA mikroinjektion
- RNA syntetiseras från linjäriserade DNA mallar med mMessage mMachine kit (Ambion) och utspätt med RNase-fritt vatten på lager koncentration av 0,1-1 mikrogram / l. Optimal doser av RNA för injektioner bestäms i pilotförsök. RNA för Diversin-RFP, membranet markör GFP-CAAX och krulliga 8 används vid 0,1-1 ng per injektion.
- Kanyler är förberedda med en nål avdragare från en kapillär och sedan med en nål kvarn. Före injektion är varje nål kalibreras med vatten för att mata ut 10 nl vätska per injektion.
- För injektioner, är embryon placeras på en plast skål i en stor droppe 3% Ficoll, 0,5 x MMR. En mikroliter av RNA lösningen sugs in i en injektionsnålen med Narishige microinjector. 10 nl av RNA injiceras i djur blastomerer av 8 cells embryon 2-3 gånger. Den injicerade embryona förs in i en brunn på 12-brunnar.
3. Förbereda Ectodermal explants
- När den injicerade embryona nå tidigt gastrulastadium, de överförs till 0,6 x MPR-lösning i en 3 cm plast fat täckt med 1% agaros. Vitelline membranet avlägsnas manuellt med en pincett. Ektodermal explants är exciderad från embryon med hjälp av en Tungsten nål och en hairloop.
- Ektodermal explants överförs till en injektionsflaska av glas och fixeras med 3,7% formaldehyd i fosfatbuffrad saltlösning (PBS) i 30 minuter. Fast explants tvättas med PBS tre gånger (10 minuter vardera). DAPI ingår i den tredje tvättningen att färga kärnor.
- Explants är monterade på en bild glas. Två remsor av tejp är knutna till bilden och explants placeras mellan de två remsorna. Eftersom utsidan av explants är pigmenterade, bör insidan av explants ansiktet mikroskop målet. Tillsätt 2-3 20 l droppar av monteringslösning (70% glycerol i PBS inklusive 25 mg / ml DABCO, anti-fading reagens) 11. Sätt på ett täckglas ovanpå.
4. Avbildning av explants Enligt ett fluorescerande mikroskop
- Prover ses under Zeiss Axioplan fluorescerande mikroskop med lämpliga filter.
- Bilderna är tagna med Apotome fastsättning att visualisera en specifik plan från flera oberoende explants.
5. Cryosectioning
Cryosectioning är ett alternativt sätt att visualisera fördelningen av fluorescerande proteiner i cellen och mer som gäller för immunfärgning. Vid stadium 10, embryon fastställs för 1-2 timmar med presidentens fixativ (20% DMSO, 80% metanol), tvättas med PBS, och inbäddade i 15% fisk gelatin/15% sackaroslösning 11. Den inbäddade embryona snabbt fryses på torris och kryosnitt genereras på Leica kryostat. Tvärsnitt skulle omfatta ektodermal celler som ärver injiceras RNA och deras översatta produkter protein. Avsnitten behålla fluorescens och kan immunostained med specifika antikroppar och sedan märkt med sekundära antikroppar konjugerade med fluorescens. Kärnor färgas med DAPI. Monteringen media är samma som beskrivits ovan. Imaging kan utföras som beskrivits ovan.
6. Representativa resultat:
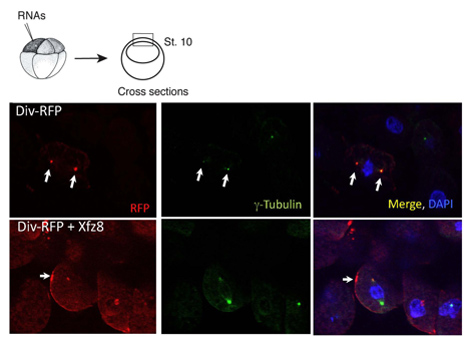
Figur 1. Krulliga receptorn rekryterar Diversin till cellmembranet. Ektoderm celler som uttrycker krulliga 8 (Xfz8) och div-RFP RNA avslöjar Div-RFP i cellmembranet, i stället för centrosome (som avslöjades av g-tubulin co-färgning). Systemet med experimentet är på toppen, ett typiskt tvärsnitt visas.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Vi har använt ovan nämnda protokollet för att karakterisera subcellulära lokalisering av Diversin. I explants djur pol, var Diversin-RFP upptäckt intill kärnan och colocalized med G-tubulin, ett centrosomal markör i kryosnitt (figur 1). När subcellulära lokalisering av ett protein som identifierats kan radering konstruktioner skapas för att fastställa vilket protein domäner är nödvändiga och tillräckliga för subcellulära lokalisering. Med hjälp av denna metod har centrosomal lokalisering domäner Diversin kartlagts i mitten och i karboxi-terminal domäner av proteinet, som båda innehåller den ihoprullade-coil motiv 8.
Samma protokoll kan användas i studier, i vilket protein lokalisering ändras som svar på signaler. Vi fann att Wnt utsöndrade proteiner agera för att flytta Div-RFP till punktat konstruktioner i närheten av cellmembranet, medan FZ8 rekryterar Div-RFP till fläckar cellmembranet (Figur 1). Vi upptäckte även att karboxi-terminal domän är inte nödvändigt för membran rekrytering i sig, men som krävs för polariserad membran rekrytering.
Sammanfattningsvis kommer ovanstående försöksprotokoll hjälp i olika studier av protein-protein interaktioner och protein lokalisering till olika cellulära avdelningar efter stimulering av celler med specifika tillväxtfaktorer eller signalsystem proteiner.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Inga intressekonflikter deklareras.
Acknowledgments
Forskning i Sokol laboratoriet är sponsrade av National Institues of Health.
References
- Gurdon, J. B. Embryonic induction --- molecular aspects. Development. 99, 285-306 (1987).
- Principles of Developmental Genetics. Moody, S. A. , Academic Press. (2007).
- Clevers, H.
- Klaus, A., Birchmeier, W. Wnt signalling and its impact on development and cancer. Nat Rev Cancer. 8, 387-398 (2008).
- Gordon, M. D., Nusse, R. Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors. J Biol Chem. 281, 22429-22433 (2006).
- Schwarz-Romond, T., Asbrand, C., Bakkers, J., Kuhl, M., Schaeffer, H. J., Huelsken, J., Behrens, J., Hammerschmidt, M., Birchmeier, W. The ankyrin repeat protein Diversin recruits Casein kinase Iepsilon to the beta-catenin degradation complex and acts in both canonical Wnt and Wnt/JNK signaling. Genes Dev. 16, 2073-2084 (2002).
- Moeller, H., Jenny, A., Schaeffer, H. J., Schwarz-Romond, T., Mlodzik, M., Hammerschmidt, M., Birchmeier, W. Diversin regulates heart formation and gastrulation movements in development. Proc Natl Acad Sci U S A. 103, 15900-15905 (2006).
- Itoh, K., Jenny, A., Mlodzik, M., Sokol, S. Y. Centrosomal localization of Diversin and its relevance to Wnt signaling. J. Cell Sci. 122, 3791-3798 (2009).
- Itoh, K., Jacob, J., Sokol, Y. S. A role for Xenopus Frizzled 8 in dorsal development. Mech Dev. 74, 145-157 (1998).
- Newport, J., Kirschner, M. A major developmental transition in early Xenopus embryos: I. characterization and timing of cellular changes at the midblastula stage. Cell. 30, 675-686 (1982).
- Itoh, K., Brott, B. K., Bae, G. U., Ratcliffe, M. J., Sokol, S. Y. Nuclear localization is required for Dishevelled function in Wnt/beta-catenin signaling. J Biol. 4, 3-3 (2005).
