

Summary
시험관 내에서 액토미오신 다발의 형성 및 광학 핀셋을 사용한 미오신 앙상블 힘 생성 측정이 제시되고 논의됩니다.
Abstract
미오신은 ATP를 가수분해하여 액틴 필라멘트(AF) 트랙을 따라 이동하는 운동 단백질이며 운동성 및 근육 수축과 같은 세포 과정에 필수적입니다. 힘 생성 메커니즘을 이해하기 위해 미오신 II는 단일 분자 (SM) 수준과 광학 트래핑과 같은 생물 물리학 적 방법을 사용하는 시험관 내 모터 팀으로 조사되었습니다.
이 연구는 미오신 힘 생성 거동이 3 비드 배열의 단일 분자 수준에서 글라이딩 배열의 단단한 비드 또는 커버 슬립 표면에서 함께 작동하는 모터 그룹으로 이동할 때 크게 다를 수 있음을 보여주었습니다. 그러나 이러한 분석 구조는 세포 내에서와 같이 점탄성 구조 계층 내에서 미오신의 그룹 역학을 평가하는 것을 허용하지 않습니다. 우리는 여러 액틴 필라멘트와 상호 작용하는 미오신 앙상블에 의한 힘 생성 메커니즘을 조사하기 위해 광학 핀셋을 사용하는 방법을 개발했습니다.
이러한 액토미오신 번들은 모터 통신 및 앙상블 힘 출력을 캡처하는 계층적이고 규정을 준수하는 환경에서 조사를 용이하게 합니다. 분석의 사용자 정의 가능한 특성을 통해 실험 조건을 변경하여 미오신 앙상블, 액틴 필라멘트 번들 또는 주변 환경에 대한 수정이 어떻게 다른 힘 출력을 초래하는지 이해할 수 있습니다.
Introduction
운동 단백질은 생명에 필수적이며 화학 에너지를 기계적 작업으로 변환합니다 1,2,3. 미오신 모터는 트랙과 유사한 필라멘트를 따라 단계를 밟아 액틴 필라멘트와 상호 작용하며, 액틴-미오신 네트워크의 역학은 근육 수축, 세포 운동성, 세포질 분열 중 수축 고리 및 세포 내부의화물 이동을 수행합니다. 3,4,5,6,7,8 . 미오신은 많은 필수 역할을 하기 때문에 미오신 액틴 네트워크의 기능 장애는 비대성 심근병증(HCM)에서 심장 과수축을 유발하는 미오신 중쇄의 돌연변이와 같은 질병 발병으로 이어질 수 있습니다.9,10,11,12,13,14 . 근육 수축에서 개별 미오신 모터는 AF 4,15,16,17,18의 상대 슬라이딩을 수행하는 데 필요한 기계적 에너지를 제공하기 위해 앙상블로 작동하여 서로 협력합니다. 미오신 모터는 AF 사이에 교차 다리를 형성하고 기계 화학적 주기로 인한 구조적 변화를 사용하여 정렬된 필라멘트17,18,19,20,21의 가시 끝을 향해 집합적으로 이동합니다.
광학 트래핑과 같은 기술을 사용하여 SM 수준에서 정량적 체외 운동성 분석을 개발함으로써 SM 힘 생성 및 스텝 크기 측정을 포함하여 개별 미오신 모터가 어떻게 작동하는지에 대한 전례 없는 세부 정보를 쉽게 수집할 수 있었습니다. 22,23,24,25,26,27,28,29,30 . Finer et al.은 단일 미오신 II 모터23,31의 힘 발생 역학을 조사하기 위해 "3-비드" 또는 "덤벨" 광학 트래핑 분석을 개발했습니다. 근육 미오신 II는 AF를 수축시키기 위해 팀으로 작동하지만 SM 수준에서는 비과정적이기 때문에 광학 트래핑 분석 방향은 고전적인 모터 결합 비드 접근법(32)으로부터 재배열되어야 했습니다. 덤벨 분석을 형성하기 위해, 커버슬립-부착된 비드에 결합된 미오신 모터 위에 AF를 유지하기 위해 2개의 광학 트랩을 사용하였고, 단일 모터에 의해 출력된 힘은 트랩(23) 내의 AF의 이동을 통해 측정되었다.
그러나 SM 힘과 단일 모터 / 단일 필라멘트 분석 방향을 사용하면 myosin II를 포함한 많은 모터 단백질이 단독으로 작동하지 않고 종종 부분의 합으로 기능하지 않기 때문에 시스템 수준 힘 생성에 대한 전체 이미지를 제공하지 않습니다 15,16,17,32,33,34,35,36 . 하나 이상의 필라멘트와 상호 작용하는 하나 이상의 모터를 포함하는 더 복잡한 구조는 미오신 및 액틴 필라멘트의 네트워크(15,32)의 시너지 효과를 더 잘 이해하는 데 필요합니다. 덤벨 분석 방향은 비드에 여러 개의 미오신을 부착하거나 표면에 부착된 미오신 두꺼운 필라멘트를 사용하여 모터가 매달린 AF 4,23,34,37,38,39,40과 상호 작용할 수 있도록 함으로써 작은 앙상블 힘 생성을 조사하는 데 활용되었습니다.
다른 작은 앙상블 분석에는 미오신 모터가 커버슬립 표면에 코팅되고 AF에 결합된 비드가 모터 4,35,36,38,39,40,41,42,43 팀에 의해 생성된 힘을 프로브하는 시험관 내 필라멘트 글라이딩 분석이 포함됩니다. . 이 두 경우 모두 미오신은 단단한 표면(비드 또는 커버슬립)에 결합되어 하나의 AF를 사용합니다. 이러한 경우들에서, 모터들은 자유롭게 움직일 수 없거나 서로 통신할 수 없으며, 근육신이 단단히 결합된 것은 모터들이 sarcomere32에서 함께 동작할 수 있는 순응적이고 계층적인 환경을 반영하지 않는다. 이전 연구에서는 미오신 II가 환경을 감지하고 힘 생성 및 듀티 비율41,44,45와 같은 특성을 변경하여 변화하는 점탄성 또는 모터 집중 조건에 따라 적응할 수 있다고 제안했습니다. 따라서 미오신 II 앙상블 힘 생성의 기계론적 토대에 대한 보다 현실적인 그림을 그리기 위해 모터 통신 및 시스템 준수를 촉진하고 캡처하는 광학 트래핑 분석을 개발할 필요가 있습니다.
여기에서는 두 액틴 필라멘트 사이에 상호 작용하는 여러 개의 미오신 모터로 구성된 액토미오신 다발 또는 샌드위치를 형성하여 시험관 내 계층 구조와 광학 트래핑을 결합하는 방법을 개발했습니다. 이 모듈식 분석 기하학은 분자 및 환경 요인이 앙상블 미오신 힘 생성에 어떻게 영향을 미치는지 직접 조사할 수 있습니다. 또한 이러한 액틴-미오신 앙상블을 통한 힘 생성 메커니즘을 조사하면 근육 수축과 같은 대규모 세포 작업이 분자 수준 9,10,13에서 어떻게 전파되는지 모델링하고 이해하는 데 도움이 될 수 있습니다.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 에칭 커버슬립
- 1,000mL 비커에 100% 에탄올 300mL에 KOH 100g을 녹입니다. 대부분의 KOH가 녹을 때까지 교반 막대로 저어줍니다.
주의 : 농축 KOH 용액은 화상을 입거나 의복을 손상시킬 수 있습니다. 장갑, 보안경 및 실험실 코트를 착용하십시오. - 커버슬립을 커버슬립 청소용 랙에 개별적으로 놓습니다.
알림: 랙은 커버슬립의 각 면, 바닥의 배수구에서 에칭 및 헹굼이 가능하도록 단일 커버슬립을 간격을 두고 유지하는 슬릿으로 설계되었으며 가혹한 에칭 조건을 견딜 수 있는 재료로 만들어졌습니다. 그들은 맞춤 제작되거나 상업적으로 구입할 수 있습니다. - 3개의 1,000mL 비커(300mL의 에탄올이 있는 비커와 300mL의 역삼투압(RO) 물이 있는 비커 2개를 준비하고 라벨을 붙입니다.
알림: 여기에서 RO 물은 실험실 정수기에서 공급되었지만 지역 정수기를 사용할 수 없는 경우 상업적으로 구입할 수도 있습니다. - 4 개의 비커 각각을 욕조 초음파 처리기에 넣어 5 분 동안 가스를 제거합니다.
- KOH와 에탄올의 비커에 커버 슬립 랙을 담그고 5 분 동안 초음파 처리합니다.
- 커버슬립 랙을 KOH/에탄올 비커에서 에탄올 전용 비커로 옮깁니다. 구슬이 생기지 않을 때까지 랙을 비커에 위아래로 담그십시오.
알림: 커버슬립을 방해하거나 랙을 비커에 강제로 떨어뜨리지 않도록 주의하십시오. 이로 인해 커버슬립이 랙에서 나오거나 화학 물질이 튀게 됩니다. - 커버 슬립 랙을 에탄올 비커에서 물 비커로 조심스럽게 옮기고 구슬이 없을 때까지 위아래로 담그십시오.
- 아직 사용하지 않은 물의 비커에 커버 슬립 랙을 담그고 5 분 동안 다시 초음파 처리합니다.
- 병을 사용하여 커버슬립 랙에 물이 커버슬립에서 부드럽게 흘러내릴 때까지 스프레이합니다. 에탄올로 반복하십시오.
- 건조할 랙을 90°C의 오븐에서 20분 동안 놓습니다. 에칭된 커버슬립 랙은 사용하기 전에 오염을 방지하기 위해 밀폐된 용기에 실온에서 보관하십시오.
2. 액틴 필라멘트 중합
- 솔루션 T 만들기
- 50mL 원추형 튜브에 3.94g의 트리스-HCl과 0.147g의CaCl2를 추가합니다. RO 물을 넣어 총 부피가 50mL가 되도록 하고 잘 섞습니다.
참고: 용액 T의 최종 농도는 각각 500mM Tris-HCl 및 20mMCaCl2 입니다. - 튜브 용액 T에 라벨을 붙이고 4 ° C에서 보관하십시오.
- 50mL 원추형 튜브에 3.94g의 트리스-HCl과 0.147g의CaCl2를 추가합니다. RO 물을 넣어 총 부피가 50mL가 되도록 하고 잘 섞습니다.
- TC 버퍼 만들기
- 40mL의 RO 물과 1.5mL의 용액 T를 50mL 원뿔형 튜브에 섞습니다. 소량의 농축 KOH를 첨가하여 pH를 8.0으로 변경합니다. 물을 넣어 용액 50mL를 만들고 pH를 확인합니다. 필요한 경우 pH를 조정하십시오.
참고: 최종 TC 완충액은 pH 8에서 5mM 트리스-HCl 및 0.2mMCaCl2 를 포함합니다. - 튜브 TC에 라벨을 붙이고 4 ° C에서 보관하십시오.
- 40mL의 RO 물과 1.5mL의 용액 T를 50mL 원뿔형 튜브에 섞습니다. 소량의 농축 KOH를 첨가하여 pH를 8.0으로 변경합니다. 물을 넣어 용액 50mL를 만들고 pH를 확인합니다. 필요한 경우 pH를 조정하십시오.
- FC 버퍼 만들기
- 85mL의 RO 물, 10mL의 용액 T, 3.73g의 KCl 및 0.041g의MgCl2 를 100mL 버퍼 병에 추가합니다. 소량의 농축 KOH를 첨가하여 pH를 7.5로 수정합니다. 물을 추가하여 최종 부피를 100mL로 만들고 pH를 확인합니다.
참고: 최종 FC 완충액은 pH 7.5에서 500mM 트리스-HCl, 500mM KCl, 2mM MgCl2 및 2mMCaCl2를 함유한다. - 튜브 FC에 라벨을 붙이고 4 ° C에서 보관하십시오.
- 85mL의 RO 물, 10mL의 용액 T, 3.73g의 KCl 및 0.041g의MgCl2 를 100mL 버퍼 병에 추가합니다. 소량의 농축 KOH를 첨가하여 pH를 7.5로 수정합니다. 물을 추가하여 최종 부피를 100mL로 만들고 pH를 확인합니다.
- 일반 액틴 완충액 (GAB)을 준비하십시오.
- 마이크로 원심분리 튜브에서 485μL의 TC 버퍼, 10μL의 10mM ATP 및 5μL의 50mM DTT를 혼합합니다.
참고: 최종 완충액 조건은 5 mM 트리스-HCl, 0.2 mMCaCl2, 0.5 mM DTT, 및 0.2 mM ATP입니다. - GAB로 라벨을 붙이고 4 ° C에서 보관하십시오.
- 마이크로 원심분리 튜브에서 485μL의 TC 버퍼, 10μL의 10mM ATP 및 5μL의 50mM DTT를 혼합합니다.
- 액틴 중합 완충액(APB)을 준비합니다.
- 마이크로 원심분리 튜브에서 455μL의 FC 완충액, 25μL의 100mM ATP 및 20μL의 50mM DTT를 혼합합니다.
참고: 최종 완충액 조건은 50 mM 트리스-HCl, 500 mM KCl, 2 mM MgCl2, 2 mMCaCl2 2 mM DTT, 및 5 mM ATP이다. - 튜브에 APB라고 표시하고 4 ° C에서 보관하십시오.
- 마이크로 원심분리 튜브에서 455μL의 FC 완충액, 25μL의 100mM ATP 및 20μL의 50mM DTT를 혼합합니다.
- 액틴 재구성
- 동결건조된 액틴 1mg 바이알에 100μL의 탈이온수를 추가하여 토끼 골격근 액틴을 재구성합니다. 위아래로 부드럽게 피펫팅하여 잘 섞습니다. 5 μL 샘플에 분취하고, 스냅 동결하고, -80 °C에서 10 mg/mL 액틴 분취량을 보관한다.
- RO 물 20μL를 추가하여 비오티닐화된 토끼 골격근 액틴을 재구성합니다. 표본량을 5μL 샘플로 분취하고, 스냅 동결하고, -80°C에서 1mg/mL 비오티닐화된 액틴 분취량을 보관합니다.
- 로다민 팔로이딘 안정화를 통한 표지되지 않은 액틴 중합
- 액틴 10mg/mL 바이알 하나를 해동하고 얼음 위에 보관하십시오.
- 새로운 GAB 완충액을 준비하고 액틴 분취량에 100μL의 GAB를 추가하고 위아래로 부드럽게 피펫팅하여 혼합합니다. 용액을 얼음 위에서 1 시간 동안 배양하십시오.
- 배양 중에 신선한 APB를 준비하십시오. 배양 후, 액틴 용액에 11μL의 APB를 첨가하여 액틴을 필라멘트로 중합한다. 위아래로 부드럽게 피펫팅하여 잘 섞습니다. 얼음 위에 20분 동안 둡니다.
- 5 μL의 로다민 표지 팔로이딘을 새로 중합된 액틴 필라멘트 용액에 추가합니다. 어둠 속에서 얼음 위에 1 시간 동안 두십시오.
- 알루미늄 호일에 싸인 로다민 액틴 바이알을 4°C의 어두운 곳에 보관하십시오.
알림: 이 필라멘트는 최대 1주일 동안 사용하는 것이 좋습니다. AF만 포함된 플로우 셀의 빠른 이미징과 매일 일관된 필라멘트 보기를 통해 AF 품질을 매일 확인할 수 있습니다.
- Alexa Fluor 488 팔로이딘 안정화를 통한 비오틴화 액틴 중합
- 액틴 10mg/mL 바이알 1개와 비오티닐화 액틴 1바이알 1개를 해동하여 얼음 위에 보관하십시오.
- 새로운 GAB 버퍼를 만듭니다.
- 두 바이알(단계 2.8.1)을 10:1 액틴:비오티닐화 액틴 비율로 결합합니다. 액틴 혼합물에 100μL의 GAB를 넣고 위아래로 부드럽게 피펫팅하여 잘 섞습니다. 얼음 위에서 1 시간 동안 배양하십시오.
- 잠복기 동안 신선한 APB를 만드십시오.
- 배양 단계 후, 11 μL의 APB를 액틴 용액에 첨가하여 액틴을 중합한다. 위아래로 부드럽게 피펫팅하여 잘 섞습니다. 얼음 위에서 20분 동안 배양합니다.
- 5μL의 Alexa Fluor 488 표지 팔로이딘을 추가하고 어둠 속에서 얼음 위에서 1시간 동안 배양합니다.
- 알루미늄 호일에 싸인 비오티닐화된 액틴 바이알을 4°C의 어두운 곳에 보관한다.
알림: 이 필라멘트는 최대 1주일 동안 사용할 수 있습니다.
3. 미오신 및 비드 준비
- 미오신 II 재구성
- 동결 건조 된 골격 미오신 II를 간단히 스핀 다운 (~ 5 초)하여 표준 미니 원심 분리기를 사용하여 튜브 바닥에서 수집합니다.
- RO 물에서 준비된 1mM DTT 100μL를 추가하여 미오신을 10mg/mL로 재구성합니다.
- RO 물에서 1mM DTT의 90μL에 10mg/mL 미오신 10μL를 첨가하여 스톡 미오신 용액을 10배 희석합니다. 소량(1-5μL) 분취량을 만들고 스냅 동결하고 -80°C에서 보관하십시오.
참고: 미오신 활성은 이전에 발표된46,47에 게시된 표준 글라이딩 필라멘트 분석을 수행하여 확인할 수 있습니다. 간단한 설명은 토론을 참조하십시오.
- 스트렙타비딘 코팅 비드 청소
- 20μL의 1μm 스트렙타비딘 비드를 80μL의 RO 물에 희석합니다. 9,600 × g 에서 스핀다운하고 100 μL의 RO 물에서 재구성하여 4회 세척합니다.
- 40% 진폭에서 2분 동안 초음파 처리하고 세척된 비드를 4°C의 회전 장치에 보관합니다.
4. 플로우 셀 준비
- 50mL 튜브에 100% 에탄올 30mL를 넣고 물에 0.1% w/v 폴리-l-라이신 200μL를 첨가하여 폴리-l-라이신 용액(PLL)을 준비하고 잘 섞습니다.
- 에칭된 커버슬립을 PLL 용액에 추가하고 15분 동안 담가둡니다. 핀셋으로 커버슬립을 제거하고 커버슬립이 튜브에서 당겨질 때 커버슬립의 가장자리에만 닿도록 주의하십시오(그림 1A-C 참조). 장갑을 낀 손으로 커버슬립의 가장자리를 잡습니다.
- 에탄올이 남지 않고 커버슬립에 잔여물이 없을 때까지 여과된 에어라인으로 커버슬립을 건조시킵니다.
- 양면 접착 테이프 두 장을 현미경 슬라이드 중앙에 서로 3-4mm 간격으로 붙입니다. 슬라이드 가장자리에 매달려있는 여분의 테이프를 찢거나 잘라냅니다.
- 현미경 슬라이드의 장축에 수직인 테이프 위에 PLL 코팅 커버슬립을 추가하여(T를 형성함) 채널을 형성합니다.
- 작은 튜브를 사용하여 커버슬립을 테이프에 압축하고 테이프가 투명해질 때까지 현미경으로 완전히 밉니다(그림 1A). 유동 채널에서 누출이 발생할 수 있으므로 테이프에 기포가 없는지 확인하십시오.
참고: 플로우 셀은 10-15 μL의 부피를 유지할 수 있습니다.
5. 액토미오신 번들 준비
- 별도의 튜브에서 각 유형의 액틴 필라멘트(로다민 및 비오티닐화 488-표지)를 600μL의 정면, 표지된 액틴과 300μL의 APB를 혼합하여 600x 희석합니다. 각 튜브에 상응하는 라벨이 붙은 팔로이딘 5μL를 추가로 추가하고 어둠 속에서 15분 동안 얼음 위에서 배양합니다.
- 비오틴화 액틴 용액에 500mg/mL에서 베타-D-포도당 1μL, 25mg/mL에서 포도당 산화효소 1μL, 500단위/mL에서 카탈라아제 1μL의 산소 소거 시스템을 추가합니다. 1μL의 100mM ATP와 1μL의 100x 희석되고 세척된 스트렙타비딘 비드를 추가합니다. 피펫 팁으로 부드럽게 저어줍니다. 현탁액을 4°C에서 회전기에 놓고 나머지 액토미오신 다발이 조립되는 동안.
- 희석된 로다민 액틴 15μL를 PLL 플로우 셀에 추가합니다(그림 1D). 유동 셀을 통해 초과 용액을 흡수하되 유동 채널이 건조되지 않도록 하십시오. 습도 챔버에서 10 분 동안 배양하십시오.
알림: 습도 챔버는 바닥에 물을 추가하고 뚜껑을 알루미늄 호일로 덮어 빛을 차단하는 빈 피펫 팁 상자로 만들 수 있습니다. - APB에 1mg/mL 카제인 용액을 준비합니다.
- 후속 성분의 비특이적 결합을 방지하기 위해 1mg/mL 카제인 15μL를 추가합니다(그림 1E). 습도 챔버에서 5 분 동안 배양하십시오.
- 원하는 농도의 미오신 을 단계 5.2의 비오티닐화된 액틴 및 비드 현탁액에 첨가한다. 피펫 팁으로 부드럽게 저어준 다음 즉시 15μL의 5.2단계 현탁액 + 원하는 미오신 농도를 플로우 셀에 추가합니다(그림 1F,G). 20 분 동안 배양하십시오. 이미징 및 광학 트래핑 실험 중 증발을 방지하기 위해 플로우 셀의 열린 끝을 매니큐어로 밀봉하십시오.
참고: 1μM의 미오신 용액 농도는 강력한 번들링을 제공하며 원하는 분석 맞춤화를 위한 시작점으로 사용할 수 있습니다( 그림 2 참조).
6. 옵티컬 트랩(NT2 나노트래커2)을 사용한 힘 측정
참고: 아래 프로토콜은 NT2 시스템을 위한 것이지만 이 분석은 형광 기능이 있는 맞춤형 시스템을 포함하여 다른 광학 트래핑 시스템과 함께 사용할 수 있습니다. 일반적인 워크플로우는 슬라이드 표면에 초점을 맞추고, 비드 보정을 수행하고, 형광 액틴 번들을 찾아 데이터를 수집하는 것과 동일하게 유지됩니다. NT2 시스템의 경우 보충 그림 S1, 보충 그림 S2, 보충 그림 S3, 보충 그림 S4, 보충 그림 S5, 보충 그림 S6 및 보충 그림 S7 은 광학 트래핑 시스템 및 소프트웨어 인터페이스에 대한 세부 정보를 제공합니다.
- 컨트롤 박스와 레이저를 켭니다(보충 그림 S1).
- 바탕 화면에서 JPK 나노 트래커 아이콘을 클릭하여 광학 트랩 컴퓨터 소프트웨어를 시작하십시오.
- 중앙에 있는 Logitech 버튼을 클릭하여 조종기를 깨웁니다(보충 그림 S2).
- 켜기/끄기 스위치를 토글하여 형광 모듈을 켭니다(보충 그림 S3).
- 명시야 이미징을 위해 필터 큐브 터렛을 돌립니다(보충 그림 S4).
- 시스템이 준비되면 화면 왼쪽 하단 모서리에 있는 레이저 파워 버튼을 사용하여 레이저를 50mW로 켜고 30분 동안 안정화합니다(보충 그림 S5).
- 소프트웨어 내에서 조명, 카메라, 목표 및 스테이지 이동 버튼을 순차적으로 클릭하여 실험 중에 보고 조작할 수 있도록 해당 창을 불러옵니다. On/Off 버튼을 클릭하고 막대를 클릭하고 오른쪽 끝까지 드래그하여 최대 전력으로 설정하여 현미경 조명을 켭니다(보충 그림 S5).
- 샘플 영역을 열고 현미경 스테이지에서 샘플 홀더를 제거합니다. 플로우 셀을 추가하고 금속 샘플 홀더로 고정한 다음 커버슬립이 있는 슬라이드가 바닥에 있는지 확인합니다.
- RO 물 30μL를 하단 대물렌즈 중앙에 추가합니다. 피펫 팁이 렌즈에 닿지 않도록하십시오. 샘플 스테이지를 다시 삽입합니다.
알림: NT2 시스템은 수침 대물렌즈를 트래핑 대물렌즈로 사용하므로 사용자 설정의 트래핑 대물렌즈에 따라 침수 매체가 다를 수 있습니다. - 물 비드가 커버슬립에 닿을 때까지 화면의 제어 화살표 또는 조종기의 L2를 사용하여 하단 대물렌즈를 올립니다(보충 그림 S5).
- 화면 화살표 또는 조종기의 R2를 사용하여 플로우 셀까지의 거리의 약 절반에 도달할 때까지 상단 대물렌즈를 내립니다. 170μL의 RO 물을 상단 대물렌즈 바로 아래의 플로우 셀 상단에 추가합니다. 물의 표면 장력을 깨고 반월 상 연골을 형성 할 때까지 상단 대물렌즈를 내립니다.
- 조종기의 화살표 패드를 사용하여 흐름 채널에 인접한 테이프의 가장자리에 도달할 때까지 현미경 스테이지를 이동합니다. 샘플 도어를 닫습니다.
알림: 샘플 도어를 닫을 때 "딸깍" 소리가 나면 레이저 셔터가 열려 있음을 나타냅니다. 이것은 문이 닫힌 경우에만 셔터를 열 수 있는 안전 기능입니다. - 화면의 목표 창을 사용하여 화면 컨트롤을 사용하여 위쪽 화살표를 클릭하여 레이저 목표라는 아래쪽 목표를 위로 가져와 테이프의 가장자리에 초점을 맞춥니다. 하단 화살표를 클릭하여 상단 목표에 대해서도 동일한 작업을 수행합니다(보충 그림 S5).
참고: 이중 화살표는 목표나 스테이지를 더 빠르게 이동합니다. 테이프의 가장자리는 커버 슬립 표면에 가까운 크고 찾기 쉬운 물체이기 때문에 초점을 맞추는 데 사용됩니다. 테이프 내의 기포는 또 다른 옵션입니다. 그러나 사용자가 표면 초점을 찾기 위한 자동화된 루틴 또는 선호하는 사내 방법을 가지고 있는 경우에는 필요하지 않습니다. - 테이프에 초점이 맞춰지면 광학 트랩 상단의 조리개를 부분적으로 닫습니다. 홍채의 다각형 모양이 보일 때까지 상단 목표를 아래로 가져옵니다. 가장자리에 초점을 맞추고 조리개를 다시 연 다음 자물쇠 아이콘을 클릭하여 대물렌즈를 연결합니다(보충 그림 S5).
- 플로팅 비드를 찾아 트랩 셔터 버튼을 클릭하여 트래핑하면 셔터 가 열리고 트래핑 레이저가 샘플에 닿을 수 있습니다. 화면에서 트랩 커서를 클릭하고 드래그하여 트래핑 레이저의 위치를 이동합니다. 일단 갇히면 비드를 보정하여 전압 측정값을 힘 및 변위와 연관시킵니다.
- 보정 버튼을 클릭합니다. 전력 스펙트럼 분석을 기반으로 교정 루틴을 조정하고 X, Y 및 Z 방향에 대해 소프트웨어 내의 코너 주파수를 맞춥니다(보충 그림 S6).
- 설정을 클릭합니다. 비드의 직경(1,000nm)을 입력하고 소프트웨어 창의 왼쪽 하단에 있는 스테이지의 온도를 입력합니다. (보충 그림 S6 참조).
- 트랩 1을 클릭합니다. X 신호를 클릭합니다. 실행을 클릭하여 코너 주파수 피팅을 수행합니다. 창 내에서 클릭하고 드래그하여 기능 맞춤을 최적화합니다. 민감도 및 강성 값에 사용을 클릭합니다. 값 수락을 클릭합니다. Y 및 Z 신호에 대해 반복합니다. 창을 닫습니다. (보충 그림 S6 참조).
참고: 등분할 방법 또는 항력 방법과 같이 사용자가 강력하게 테스트한 다른 광학 트래핑 시스템 또는 맞춤형 시스템의 비드 교정 루틴도 허용됩니다57,58. - 커버슬립 표면에서 AF에 바인딩된 비드를 검색하여 액토미오신 번들을 찾습니다.
- 다른 플로팅 비드에 의해 붐비지 않은 비드가 감지되면 형광 이미징으로 비드 주변의 AF를 관찰하여 번들이 있는지 확인합니다.
- 두 형광 AF가 모두 공동 국소화되었는지 확인하여 번들이 있는지 확인합니다. 백색 광원을 켜고 적절한 필터 큐브를 사용하여 터렛을 돌려 각 액틴 필라멘트를 이미지화합니다(Alexa Fluor 488 및 로다민 여기용 각각 488nm 및 532nm 여기 필터 큐브). 보충 그림 S4를 참조하십시오.
참고: 단일 AF의 형광 강도를 확인하기 위한 대조 실험은 단일 488 및 단일 로다민 표지 필라멘트로 구성되거나 사용자가 사용하기로 선택한 형광단 세트에 적용할 수 있는 번들을 식별하는 데 유용할 수 있습니다. - 확인되면 트랩 셔터 버튼을 클릭하여 번들의 상단 필라멘트에 부착된 비드를 트랩 합니다.
- 화면 컨트롤을 사용하여 오실로스코프 버튼을 클릭하여 데이터를 기록하십시오 (보충 그림 S7). 데이터를 기록하지 않고 측정값을 시각화하려면 시작을 클릭합니다. 모든 데이터를 저장하려면 자동 저장을 클릭하십시오. 측정값을 기록하려면 기록 시작을 클릭합니다. 드롭 다운 메뉴에서 X 신호 또는 Y 신호를 선택하여 실시간으로 시각화 할 데이터 (위치, 힘, x 방향, y 방향)를 선택하십시오. x-방향은 왼쪽에서 오른쪽으로, y 방향은 화면에서 위아래입니다. 보충 그림 S7을 참조하십시오.
알림: 데이터는 .out 파일로 저장되며 각 방향에 대한 시간, 전압, 변위 및 힘을 포함합니다. 이러한 파일은 시각화 및 분석을 위해 다른 소프트웨어로 내보낼 수 있습니다.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
액토미오신 번들 시스템을 포함하는 플로우 셀은 현미경 슬라이드와 양면 접착 테이프로 만든 채널로 분리된 에칭된 커버슬립으로 구성된 표준 설계입니다(그림 1). 그런 다음 분석은 프로토콜에 설명된 대로 단계적 소개를 사용하여 커버슬립에서 구축됩니다. 최종 분석은 주형 로다민 표지 액틴 필라멘트로 구성됩니다. 원하는 미오신 농도 (1 μM이 도 2 및 도 3의 대표적인 결과를 위해 사용되었다); 비오티닐화, 알렉사 플루오르 488-표지된 액틴 필라멘트; 1 μm 스트렙타비딘 비드; 산소 소거 시스템; ATP; 및 APB 버퍼. 유동 셀당 여러 개의 다발이 형성될 것이며, 위에서 설명한 액틴 농도는 원치 않는 상호 작용을 방지하기 위해 다발 사이에 적절한 간격을 제공합니다. 이는 또한 플로우 셀당 여러 힘 측정을 쉽게 획득하여 데이터 수집 효율성을 높일 수 있습니다. 힘 프로파일은 플로우 셀 내에서 그리고 플로우 셀에서 플로우 셀로 재현할 수 있어야 합니다.
위의 프로토콜은 상용 광학 트래핑 설정의 사용에 맞춰져 있지만, 여기에 제시된 플로우 셀 및 분석은 현미경 또는 현미경 스테이지와 결합되고 형광 이미징 기능을 보유하는 다른 상용 기기 또는 맞춤형 광학 트래핑 설정에 쉽게 활용될 수 있습니다. 위의 프로토콜에 따라 모든 플로우 셀 추가가 완료되면 슬라이드의 액토미오신 번들(그림 1)을 즉시 측정할 준비가 됩니다. 플로우 셀이 광학 트랩 현미경 스테이지에 추가되고, 다중 비드 교정 측정이 획득되며, 번들 필라멘트의 형광 공동 국소화를 통해 번들이 식별됩니다. 번들에 바인딩된 비드가 트랩되고 변위 및 해당 힘 측정이 시작됩니다. 사용자는 컴퓨터 모니터에서 실시간으로 데이터 수집을 관찰 할 수 있습니다. 플로우 셀에 사용되는 미오신 농도에 따라 번들이 즉시 상당한 움직임을 보이기 시작하거나 변위/힘의 증가를 효과적으로 확인하는 데 30초-1분이 걸릴 수 있습니다.
대표적인 힘 추적은 그림 3A 에 나와 있으며, 여기서 미오신 모터는 안정적인 힘 램프와 고원을 나타냅니다. 이러한 유형의 추적은 2-5 분에 걸쳐 발생하는 것이 일반적입니다. 그러나 알짜 힘을 생성하지 않는 액토미오신 다발을 측정하는 것도 가능합니다(그림 3B). 이러한 트레이스는 기준선 노이즈로 나타나거나 90초 동안 힘이 크게 증가하지 않습니다. 이는 생산적인 슬라이딩을 허용하지 않는 모터의 국소 농도가 낮거나 번들이 필라멘트의 플러스와 마이너스 끝이 정렬되는 불리한 평행 방향에 있기 때문일 수 있습니다.
플로우 셀의 내용물은 입사 조명 및 트래핑 레이저로 인한 열화, 시간 경과에 따른 슬라이드의 국소 가열 및 라디칼 산소 종의 생성에 취약할 수 있으므로 동일한 플로우 셀을 1시간 이상 사용하지 않는 것이 좋습니다. 효율성을 극대화하려면 데이터를 수집하는 동안 다른 분석을 배양하는 것이 좋습니다. 변위/힘 추적은 추가 필터링 및 분석을 위해 광학 트래핑 소프트웨어에서 Excel, Matlab, Igor, 또는 기타 데이터 관리 프로그램으로 내보낼 수 있습니다. 이러한 광학 트래핑 앙상블/번들 실험에서 추출할 수 있는 데이터에는 다양한 분석 조건에서 다양한 유형의 힘 생성 프로파일(기준선, 램프/고원), 힘 생성 속도, 최대 힘 생성, 단계 크기 및 단계 또는 단계 팀 간의 체류 시간을 통한 앙상블 운동 및 스테핑 거동, 듀티 비율이 포함됩니다. 사용자는 또한 분석 조건을 변경하여 다양한 유형의 미오신 모터 추가, 액틴 결합 단백질 추가 또는 완충액 조건 변경이 이러한 앙상블 힘 생성 특성에 어떤 영향을 미치는지 비교할 수 있습니다.
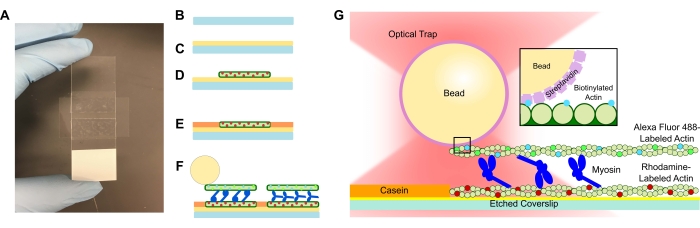
그림 1: 분석 개략도. (A-C) 에칭된 커버슬립은 폴리-L-라이신으로 코팅되고 양면 테이프와 현미경 슬라이드를 사용하여 플로우 셀을 형성하는 데 사용됩니다. 프로토콜에 설명된 시간 제한 도입 및 배양 단계를 수행하면 로다민 표지된 팔로이딘 안정화 액틴이 주형 또는 하단 필라멘트(D)로 생성되고, 이어서 비특이적 결합을 방지하기 위한 카제인 차단(E) 및 (F) Alexa Fluor 488 팔로이딘-안정화 비오티닐화 액틴을 화물 또는 상단 필라멘트로 사용하고, ATP가 도입될 때 필라멘트를 분리하고 힘을 생성하는 미오신 II 팀이 발생합니다. 모터의 기하학적 구조 및 번들 내의 가교결합의 특성은 염 농도(59)와 같은 상이한 조건 하에서 변할 수 있다. 이전의 연구는 미오신 꼬리 도메인이 액틴 필라멘트와 상호 작용하고 앙상블 운동성을 늦추는 능력을 가지고 있음을 입증했습니다46. 그러나, 무거운 메로미오신 실험에서 미오신 머리는 인접한 액틴 필라멘트(60)에 대한 각각의 머리의 결합을 입증한다. (G) 스트렙타비딘 비드는 트랩의 광학 핸들로 사용되며 화물 비오티닐화된 액틴 필라멘트에만 결합되어 슬라이드에 적절한 다발이 형성되었는지 확인하는 데 도움이 됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 2: 형광 액토미오신 다발. 그림 1에 제시된 번들 분석 내에서 액틴 필라멘트와 번들의 네 가지 다른 만남. Alexa Fluor 488 팔로이딘 채널이 있는 상단 화물 비오티닐화 액틴 필라멘트가 왼쪽에 표시되고 로다민 팔로이딘 채널이 있는 하단 템플릿 액틴 필라멘트가 오른쪽에 표시됩니다. 하단에는 눈을 안내하는 데 도움이 되도록 컬러 선이 겹쳐진 동일한 그림이 표시됩니다. (A) 상단 액틴 필라멘트는 하단 액틴 필라멘트 근처에서 발견되지만 불완전한 오버레이가 있습니다. 번들 실험에는 사용되지 않습니다. (B) 상단 및 하단 액틴 필라멘트는 공동 국소화되며 각 필라멘트의 강도는 번들 내의 각 단일 필라멘트임을 확인합니다. 이것은 번들 실험에 적합한 후보가 될 것입니다. (C) 자체 조립 된 로다민 필라멘트의 큰 묶음이 바닥에 있습니다. 공동 국소화 된 해당 상단 액틴 필라멘트가 있지만 하단 필라멘트가 너무 많습니다. 따라서 번들 실험에는 사용되지 않습니다. 이것은 또한 동일한 유형의 여러 액틴 필라멘트가 번들링될 때 형광 강도가 증가하는 방법의 예입니다. 사용자는 이것을 단일 필라멘트와 동일한 필라멘트 유형의 번들을 판단하는 게이지로 활용할 수 있습니다. (D) 하단 필라멘트에 해당하는 상단 필라멘트가 없으며 블리드 스루가 없음을 확인합니다. 번들 실험에는 사용되지 않습니다. Alexa Fluor 488 채널의 필라멘트 강도는 낮으며 사용중인 필터 세트 (Zeiss의 필터 세트 09) 때문이라고 생각합니다. 로다민 채널에 사용되는 필터 세트는 Zeiss의 필터 세트 43입니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.

그림 3: 미오신 II 앙상블 힘 생성. 골격 미오신 II 모터의 대표적인 흔적은 구성된 시험관 내 액틴 구조 계층 구조 내에서 힘을 생성합니다. 미오신 모터는 함께 작동하여 고원에 도달하고 힘이 유지되거나(A) 기준선 근처에서 길항을 경험할 때까지(B) 집단적이고 생산적으로 힘을 생성합니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
보충 그림 S1: 브루커/JPK 나노트래커2 광학 트랩. (A) 컴퓨터 모니터. (B) 컴퓨터 키보드. (C) 컴퓨터 타워. (D) 컨트롤러 상자. (E) 레이저 전원 공급 장치. (F) 광학 트랩 광학 상자. (G) 도립 현미경. (H) 현미경 스테이지에 대한 문. (I) 명시야와 차동 간섭 대비 이미징 사이를 전환하는 편광판 슬라이더. 이 파일을 다운로드하려면 여기를 클릭하십시오.
추가 그림 S2: 옵티컬 트랩용 원격 제어. (A) 전동 스테이지를 배치하는 키패드. (B-C) 트랩 위치를 조정합니다. (D) A, X, B는 각각 메인 셔터를 켜고 끄고, 트랩 1 셔터와 트랩 2 셔터를 켭니다. (E) 로지텍 버튼은 컨트롤러를 깨우는 데 사용됩니다. (F) 트래핑 대물렌즈를 배치하는 데 사용되는 위쪽 및 아래쪽 버튼입니다. (G) 감지 대물렌즈를 배치하는 데 사용되는 위쪽 및 아래쪽 버튼입니다. 리모컨이 필요하지 않으며 이러한 모든 조작은 소프트웨어에서 수행할 수 있습니다. 그러나 현미경 스테이지 환경을 들여다보면서 대물렌즈와 스테이지 위치를 제어할 수 있어 편리합니다. 이 파일을 다운로드하려면 여기를 클릭하십시오.
보충 그림 S3: 광학 트랩용 형광 모듈. 89North PhotoFluor 형광 백색 광원은 도립 현미경의 뒷면에 결합되어 있습니다. 토글 스위치(화살표)로 켜고 끌 수 있습니다. 이 파일을 다운로드하려면 여기를 클릭하십시오.
보충 그림 S4: 형광 필터 큐브 터렛. 터렛(화살표)은 DIC, 로다민 또는 Alexa Fluor 488 염료의 이미징에 필요한 필터 큐브를 사용하도록 돌릴 수 있습니다. 필터 큐브를 전환하여 다른 형광단을 사용하기 위한 설정을 사용자 지정할 수 있습니다. 이 파일을 다운로드하려면 여기를 클릭하십시오.
보충 그림 S5: 나노트래커2 소프트웨어. (A) 레이저 전원 버튼 및 제어. (B) 목표 포지셔닝 창. 방향 화살표는 탐지(위쪽) 및 트래핑(아래쪽) 목표를 이동하는 데 사용됩니다. 이중 화살표는 목표를 더 빠른 속도로 이동합니다. 왼쪽 하단의 파란색과 빨간색 버튼은 목표를 분리하고 원래 위치로 되돌립니다. 이는 현미경 스테이지 안팎에서 샘플을 채취할 때 필요합니다. 대물렌즈와 자물쇠 아이콘이 있는 왼쪽의 세 번째 버튼은 대물렌즈를 "결합"하여 초점이 맞춰지고 Kohler 조명을 달성할 때 사용자가 트래핑 및 감지 대물렌즈를 z축에서 위아래로 이동할 수 있도록 합니다. (C) x축 및 y축에서 현미경 스테이지를 이동하는 데 사용되는 샘플 위치 결정 창. 이중 화살표는 스테이지를 더 빠른 속도로 이동합니다. 이 창은 상단 메뉴에서 위쪽/아래쪽 및 왼쪽/오른쪽 화살표 아이콘을 클릭하여 활성화됩니다. (D) 카메라 시각화 창. 렌치 아이콘을 사용하여 사용자 정의 이미징 조건을 설정할 수 있습니다. 이 창은 상단 메뉴에서 카메라 아이콘을 클릭하여 활성화됩니다. (E) 현미경 조명 창. 이 창은 상단 메뉴에서 전구 아이콘을 클릭하여 활성화됩니다. 이 파일을 다운로드하려면 여기를 클릭하십시오.
보충 그림 S6: 보정 창. (ᅡ) 이 창은 비드 캘리브레이션에 사용되며 상단 메뉴에서 Cal 아이콘을 클릭하여 활성화됩니다. 비드를 교정하기 위해 x, y 및 z 신호에서 코너 주파수의 최적 피팅이 달성됩니다. (B) 각 신호에 대해 왼쪽 상단의 적절한 신호 버튼을 선택합니다. (C) 실행을 클릭하고 녹색 창 (D) 내에서 클릭하고 드래그하여 맞춤을 최적화합니다. (E) 착용감에 만족하면 감도와 강성을 위해 사용을 클릭합니다. 이렇게 하면 나노미터 단위의 변위와 피코네우톤의 힘을 기록할 수 있습니다. (F) 그런 다음 왼쪽 하단의 값 수락 을 클릭합니다. y 및 z 방향에 대해 반복합니다. 이 파일을 다운로드하려면 여기를 클릭하십시오.
보충 그림 S7: 데이터 수집 창. 이 창은 위치 및 힘 데이터를 수집하는 데 사용되며 사용자가 실시간으로 측정값을 볼 수 있도록 합니다. (ᅡ) 이 창은 상단 메뉴의 x,t 아이콘을 클릭하면 활성화됩니다. (b) 사용자는 x 신호와 y 신호 보기 사이를 전환할 수 있습니다. (C) 시작을 클릭하여 데이터 시각화를 시작합니다. 자동 저장을 클릭하여 데이터를 저장합니다. 클릭 기록 시작 데이터 기록 및 저장을 시작합니다. 이 파일을 다운로드하려면 여기를 클릭하십시오.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
액틴 필라멘트와 상호 작용하는 미오신 앙상블의 역학을 조사하기 위해 형광 이미징과 결합된 광학 핀셋을 사용한 시험관 내 연구가 수행되었습니다. 악틴-미오신 - 액틴 다발은 근육 미오신 II, 번들의 하단 및 커버 슬립 표면의 로다민 액틴, 번들 상단의 488 표지 된 비오틴 액틴 필라멘트를 사용하여 조립되었습니다. 토끼 근육으로부터의 액틴 단백질은 일반 액틴 완충액 (GAB) 및 액틴 중합 완충액 (APB)을 사용하여 중합 및 안정화되었다. GAB 및 APB는 ATP, FC 버퍼 및 TC 버퍼를 사용하여 실험실에서 매일 새로 준비해야 합니다. 근육 미오신 II는 액틴-미오신 - 액틴 샌드위치를 형성하는데 사용되었다. Phalloidin은 액틴 필라멘트의 형광 염색 및 시험관 내 안정화에 사용되었습니다.
미오신 활성은 이전에 공개된46,47과 같은 표준 글라이딩 필라멘트 분석을 수행함으로써 확인할 수 있다. Myosin II 및 그의 하위단편은 다양한 배향으로 커버슬립 표면에 결합할 수 있고, 꼬리 도메인의 존재는 무거운 meromyosin46,48,49를 사용하는 분석과 비교하여 필라멘트 슬라이딩을 늦출 수 있다. 그러나 활공 및 표면 이동은 여전히 관찰 할 수 있습니다. 미오신 활성의 보다 명백한 입증은 활성 액틴 필라멘트 파괴로, 더 긴 액틴 필라멘트가 더 작은 조각으로 부서진 다음 여러 방향으로 미끄러지는 곳에서 관찰할 수 있습니다. 이것은 표면에 활성 모터의 농도가 높기 때문에 발생하며 여러 실험실에서 관찰되었으며 활성 미오신 모터가 42,50,51,52,53,54 없이는 발생하지 않습니다. 또한, 여기에 제시된 번들 분석은 번들 분석이 유리 표면의 카제인 차단을 포함하기 때문에 유리 커버슬립 상의 다양한 모터 결합 방향과 같이 주로 글라이딩 필라멘트 분석과 관련된 운동성 문제를 완화하는 데 도움이 되는데, 이는 번들 분석이 유리 표면의 카제인 차단을 포함하기 때문에 모터가 번들(47,55,56) 내에서 결합하도록 하기 때문입니다.
첫 번째 단계는 로다민 액틴 필라멘트를 플로우 셀의 폴리-L-라이신 코팅 커버슬립에 바닥 또는 템플릿 필라멘트로 추가하는 것입니다. 폴리-L-라이신은 폴리-라이신이 양전하를 띠는 반면 액틴은 음전하를 띠고 이전의 세포골격계 시험관 내 분석 제제61,62,63에 사용되었기 때문에 액틴 결합을 촉진하는 데 사용됩니다. 번들 형성 전에 액틴 농도를 최적화하기 위해 다양한 액틴 희석액을 유동 세포에 첨가했습니다. 이 경우 재고에서 600x가 번들 형성을 위해 충분한 수의 템플릿 필라멘트를 생성하지만 번들이 개별화되도록 적절한 간격을 갖는 최적의 희석이었습니다. 희석은 APB 완충액을 사용하여 수행하였다. 로다민 액틴을 첨가한 후 카제인 층을 추가하여 표면을 차단하고 비특이적 결합을 피했습니다. 유동 세포를 30분 동안 인큐베이션하고, 완충액으로 인큐베이션한 후 임의의 결합되지 않은 액틴 필라멘트를 씻어내도록 세척하였다. 마지막으로, 미오신 488/비오틴 액틴 및 스트렙타비딘 코팅 비드의 조합을 유동 세포에 추가하여 액틴-미오신 다발 형성을 촉진했습니다. 비드 농도는 표면 결합 번들을 결합하기에 충분하고 교정을 용이하게 하기에 충분한 현탁액이 있어야 합니다. 그러나 비드 농도가 너무 높으면 인접한 비드가 레이저 트랩으로 떨어지고 측정을 방해하여 트래핑 실험 중에 어려움을 겪을 수 있습니다. 미오신 모터는 슬라이드에 주입하기 직전에 조합에 추가되어 미오신 모터가화물 또는 상부 비오틴화 액틴 필라멘트와 선제적으로 응집되지 않도록 하여 하부 로다민을 결합하여 비오틴화 액틴 필라멘트를 묶습니다.
NT2 광학 트래핑 시스템은 명시야, 미분 간섭 대비(DIC) 및 에피형광 이미징 방식이 결합된 상용 광학 트랩입니다. Zeiss AxioObserver 3 도립 현미경과 결합되어 100x/NA 1.46 및 63x/NA 1.0 침수 트랩핑 및 검출 대물렌즈가 있습니다. 이 시스템에는 하나의 레이저 트랩의 클릭 및 드래그 트래핑 기능이 장착되어 있으며 이전에 나열된 모든 방식으로 이미징하는 동안 사용할 수 있습니다. 형성된 다발은 형광 이미징을 사용하여 검출되고 확인됩니다. 적절한 필터 큐브(GFP/FITC 및 TRITC/CY3)가 있는 백색 광원을 사용하면 필라멘트 이미징 간에 빠르게 전환할 수 있습니다. 광학 핀셋을 사용하여 각 힘을 측정하기 전에 서로 다른 여기 파장에서 AF를 시각화하여 공동 국소화 AF를 검증했습니다. 필라멘트는 산소 제거 시약을 사용하더라도 빠르게 포토 표백 할 수 있으므로 연구원은 번들 실험을 수행하기 전에 강도 및 노출 시간과 같은 시각화 매개 변수를 최적화하는 것이 좋습니다.
광학 트래핑은 ATP의 존재하에 스트렙타비딘 비드를 사용하여 비오틴화된 액틴 필라멘트를 결합시키고 힘 변환기로서 미오신 힘 생성을 활성화하기 위해 힘 측정을 수행하기 위해 사용되었다. 광학 트래핑으로 얻은 변위 및 힘 대 시간 데이터는 분석을 위해 트래핑 소프트웨어에서 추출되었습니다. 그러나, 상용 트래핑 소프트웨어는 또한 활용될 수 있는 분석 루틴을 제공하거나, 다른 프로그램의 커스텀 알고리즘이 트래핑 데이터를 시각화하고 분석하도록 사용자에 의해 프로그래밍될 수 있다. 맞춤형 광학 트래핑 시스템에서 사용자는 필터가 있는 백색 광원 대신 여기 레이저를 사용할 수 있으며 이 경우에도 사용할 수 있습니다. 또한, 형광 염료는 발광 스펙트럼이 겹치지 않고 블리드스루를 유발하지 않는 경우 사용자가 가질 수 있는 기존 장비에 적합하도록 변경될 수 있다.
제시된 분석은 액토미오신 앙상블 역학 영역 내에서 연구 질문에 따라 사용자가 추가로 맞춤화할 수 있는 기준 분석입니다. 일반적인 워크플로우는 유사분열 방추 32,61,63,64,65,66 의 최소 모델을 형성하는 미세소관 번들 분석과 같이 관심의 대상이 될 수 있는 다른 시험관내 세포골격 앙상블 시스템에도 적용될 수 있다. 수정에는 사용자의 기존 설정에 적합한 형광단 라벨을 변경하는 것이 포함될 수 있지만 이에 국한되지는 않습니다. 미오신 농도, 구조물 또는 이소타입 변경; 및 완충 조건 적정, 다른 측면 중에서.
이 분석을 수행할 때 잠재적인 문제가 발생할 수 있습니다. 액틴-미오신 다발을 형성할 때, 액틴 다발 내의 미오신 농도는 슬라이드 전체에 걸쳐 균질하지 않을 수 있다. 이를 수용하기 위해 전체 슬라이드에 걸쳐 여러 번들을 측정하여 모터 분포 및 힘 생성 프로파일이 적절하게 샘플링되었는지 확인합니다. 힘 데이터의 해석에 필요한 경우 번들 방향을 아는 것도 어렵습니다. 따라서 각 번들에 대해 여러 번 시도해야 합니다. 형광 겔솔린 또는 광학 트래핑 핸들보다 작은 크기의 겔솔린 코팅 비드를 통해 액틴 필라멘트 끝단 라벨링을 통합할 수도 있습니다. 형광 이미징은 또한 번들 배향을 추론하기 위해 x 및 y 구성 요소 힘을 관찰하는 데 사용할 수 있습니다. 더욱이, 미오신 응집 상태는 KCl의 빠른 희석시 발생하는 두꺼운 필라멘트의 형성과 함께 완충액의 이온 강도에 의해 크게 영향을 받기 때문에 완충액 염 농도를 적절하게 모니터링해야합니다67,68.
글라이딩 분석과 같은 다른 시험관 내 방법을 사용한 이전 연구는 미오신 도메인의 역할을 식별하고 미오신 및 다른 액틴 결합 단백질 간의 구성 및 상호 작용을 연구하는 데 도움이 되었습니다. 그러나, 이들 방법은 미오신을 단단한 표면에 결합시키는 것이 미오신 모터 사이의 배위 가능성을 제한하고, 따라서 모터 앙상블이 고듀티 비 모드(33,35,41,69)에 있는지를 결정하기 위해 발생하는 메카노센싱 피드백을 제한한다는 단점이 있다. 또한, 단일 미오신 모터 네트워크를 사용한 광학 트래핑은 미오신 모터가 서로 및 액틴 필라멘트와 상호 작용하는 방식에 대한 명확한 이해를 제공하지 않습니다. 여기에서 개발된 프로토콜은 규정을 준수하는 계층적 액틴 네트워크 내에서 미오신 모터 앙상블 역학을 조사할 수 있도록 합니다. 또한 농도, 이소 폼 및 완충 환경과 같은 모터 필라멘트 앙상블 특성 측면에서 사용자 정의가 가능하여 체계적인 조사가 가능합니다. 제시된 프로토콜은 보다 복잡한 액토미오신 네트워크의 향후 연구를 위한 플랫폼이며 전통적으로 단일 분자 연구에 사용되었던 광학 트래핑에 의해 촉진되는 변위 및 힘 생성 측정의 정밀도를 유지합니다.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
저자는 선언 할 이해 상충이 없습니다.
Acknowledgments
이 작업은 미시시피 대학교 대학원 학생위원회 연구 펠로우십 (OA), 미시시피 대학교 샐리 맥도넬-바크스데일 아너스 칼리지 (JCW, JER), 보조금 번호 NNX15AH78H (JCW, DNR)의 미시시피 우주 보조금 컨소시엄 및 보조금 번호 848586 (DNR)의 미국 심장 협회.
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
