

Summary
MITO permite a detecção de interações sensíveis transitória e estável entre as proteínas que são expressas no modelo de Saccharomyces cerevisiae organismo. Tem sido aplicado com sucesso para estudar exógenos e leveduras proteínas integrais da membrana, a fim de identificar seus parceiros interagem de uma forma de alto rendimento.
Abstract
A importância fundamental biológico e clínico de proteínas integrais da membrana solicitado o desenvolvimento de um sistema baseado em levedura para a identificação de alto rendimento de interações proteína-proteína (PPI) para as proteínas transmembrana full-length. Para este fim, nosso laboratório desenvolveu o split-ubiquitina Yeast membrana baseada Two-Hybrid (MITO) do sistema. Esta tecnologia permite a detecção sensível de interações proteína transiente e estável usando
Protocol
1. Informação de fundo
Interações proteína-proteína (IBP) são os blocos fundamentais que regem todos os envolvidos em processos celulares. Por conseguinte, é essencial que todas as interações são estritamente reguladas, a fim de manter a homeostase celular, como uma mudança neste equilíbrio biológico normalmente desempenha um papel na doença e transformação de células cancerígenas. Proteínas da membrana associados estão entre a classe mais importante de proteínas biologicamente como eles podem iniciar complexas cascatas de sinalização, e mediar a importação e exportação de várias moléculas, incluindo drogas, que tem sido de importância recente no domínio dos cuidados de saúde como problemas relacionados à resistência a drogas tornaram-se cada vez mais comum. Obter conhecimento sobre a complexidade desta classe de proteínas requer a identificação de seus parceiros de interação. Descobrir parceiros como provou desafiador, como muitas vezes requer condições adversas que devem ser otimizados para cada proteína ligada à membrana [1].
A importância fundamental biológico e clínico de proteínas integrais da membrana solicitado o desenvolvimento de um sistema baseado em levedura para a identificação de alto rendimento do PPI para as proteínas transmembrana full-length. Para este fim, desenvolvemos o split-ubiquitina Yeast membrana baseada Two-Hybrid sistema (MITO) [2-4]. Esta ferramenta permite a detecção sensível de interações proteína transiente e estável. Tem sido aplicado com sucesso para estudar proteínas exógenos e endógenos expressos no modelo Saccharomyces cerevisiae organismo [3-7]. MITO aproveita a observação de que a ubiquitina pode ser separado em duas metades:. Metade C-terminal (C ub) ea metade N-terminal (Nubi) em estudos in vivo têm mostrado que essas metades reconstituir espontaneamente, devido à sua elevada afinidade para um outro. (Figura. 1A) No entanto, a introdução de uma isoleucina 13 a glicina mutação de ponto na metade N-terminal da ubiquitina (produzindo um fragmento referido como N ub G) impede que esta re-associação espontânea [8] (Figura. 1b).
Usamos esse princípio no sistema MITO (Figura. 1c e 2). Resumidamente, a proteína isca integrante membrana é fundido a uma porção ub C, que é ligado a um fator de transcrição artificial consiste na Escherichia coli DNA-binding protein LexA eo domínio de ativação da VP16 de vírus herpes simplex. Presas são gerados pela fusão de fragmentos de DNA genômico ou cDNA derivada para a metade NubG. Uma interação entre iscas e proteínas presas em uma série de levedura leva a reconstituição de um full-length 'pseudo-ubiquitina "molécula, com subsequente reconhecimento por enzimas citosólicas deubiquitinating (camionete) e liberação proteolítica do fator de transcrição. O fator de transcrição pode então entrar no núcleo da célula, e ativar um sistema de gene repórter (geralmente envolvendo expressão da HIS3, lacZ e genes ADE2) permitindo o crescimento de cepas de leveduras em meios seletivos, o que é indicativo de isca e interação presa [ 2-4].
A identificação e caracterização das interações proteína integral de membrana irá fornecer informações para ajudar a compreender plenamente a sua função. À medida que mais precisamente entender e dissecar as funções das proteínas que interagem com proteínas integrais da membrana, podemos ganhar a introspecção na interação dinâmica envolvida na regulação destas proteínas e descobrir novos alvos que podem ter potencial terapêutico.
2. Seleção de Bait e Sistema mito apropriado
- Antes de realizar a análise MITO, verifique se sua proteína tem o seu N-e / ou C-terminal no citosol da célula. É essencial que o C tag ub-LexA-VP16 ser fundido para sua proteína em tal terminus a, uma vez que as enzimas deubiquitinating necessária para a liberação fator de transcrição estão localizadas no citosol [4].
- Em seguida, decidir qual das duas variantes principais do MITO é adequado. Para proteínas não-nativos de levedura, MITO tradicionais (tMYTH) pode ser usado, em que as iscas são overexpressed ectopicamente a partir de um plasmídeo. Para as proteínas de levedura nativa, MITO integrado (iMYTH) é o método de escolha. Em iMYTH iscas são endogenamente marcados com a tag C ub-LexA-VP16, deixando-os sob o controle de suas promotor nativo. Isto é vantajoso, pois o nível de expressão do tipo selvagem de iscas ajuda a eliminar problemas associados com a superexpressão da proteína, tais como um aumento do número de falsos positivos [3]. Com exceção da construção isca inicial e os meios de comunicação selectivas utilizadas, ambas as formas de MITO são realizadas de forma essencialmente idêntica. Para maior clareza, vamos nos concentrar principalmente sobre o uso de tMYTH neste relatório, já que esta variante pode, em princípio, ser usado com proteínas da membrana a partir de praticamente qualquer organismo e éportanto, mais amplamente aplicáveis.
3. Geração de isca e Validação
- Meios necessários e Soluções
- Estéril DDH 2 O preparado em autoclave a 121 ° C, 15 psi durante 30 minutos.
- 3-amino-1 ,2,4-triazol (3-AT) solução preparada como uma solução 1M de ações em DDH 2 O. Esterilizar pela passagem através de um filtro de 0,2 mM.
- Mídia Crescimento YPAD consistindo de um extrato de levedura% w / v, 2% w / v de peptona, 2% de glicose w / v e 100 adenina mM preparada em DDH 2 O. Esterilizar em autoclave a 121 ° C, 15 psi durante 30 minutos.
- 10x de Aminoácidos / Mix de bases de nucleotídeos: A mistura completa contém 1,0 mM Adenina, Uracila 1,8 mM, 1,0 mM Arginina, Histidina 1,0 mM, 2,3 mM Isoleucina, Leucina 7,6 mM, 1,6 mM de lisina, 10,1 mM Metionina, Fenilalanina 3,0 mM, 16,8 mM treonina, triptofano 2,0 mM, 1,7 mM Tirosina e Valina 12,8 mM, preparado em DDH 2 O. Para drop-out omitir o ácido amino necessário (s) e / ou de bases de nucleotídeos (s). Esterilizar em autoclave a 121 ° C, 15 psi durante 30 minutos.
- Sintéticos Drop-Out (SD) Media Crescimento consistindo de 0,67% w / v base nitrogenada de levedura (sem aminoácidos mas com sulfato de amônio), 2% w / v de glicose, 2% w / v agar e 1x Mix Ácido / Nucleotide Amino, preparado em DDH 2 O. Prepare líquidos e sólidos (contendo agar 2%) SD-Leucina mídia. Também preparar sólidos SD-Triptofano-Leucina e SD-Triptofano-Leucina-Adenina-Histidina mídia. Esterilizar em autoclave a 121 ° C, 15 psi durante 30 minutos. Despeje em meios sólidos 100x15 milímetros pratos de Petri.
- Sintéticos Drop-Out Crescimento (SD) Media Contendo 3-AT. Prepare SD-Triptofano-Leucina-Adenina-Histidina mídia, como descrito, mas contendo 3-AT, em concentrações de 25, 50, 75 e 100 mM. Adicionar a quantidade adequada de 3-AT a partir da solução estoque 1M estéril para a mídia depois de ter sido autoclavado e resfriado (mas ainda não solidificado). Despeje em pratos de 100x15 milímetros Petri.
- PEG / acetato de lítio Mix composto por 40% w / v PEG-3350, 120 Acetato de Lítio e 167 mM mcg / mL de DNA de esperma de salmão (tipo de sal de sódio III), elaborado em DDH 2 O. Para garantir a esterilidade desta mistura prepará-lo da água estéril e soluções (ie autoclavado 50% PEG-3350, autoclavado Acetato de Lítio 1M e 2 mg / mL Salmon Sperm DNA tipo de sal de sódio III preparado em estéril DDH 2 O).
- Enzima e reagentes para a realização de PCR.
- Kit comercial miniprep.
- Cal sodada contas de vidro (0,5 mm).
- Escherichia coli competentes células adequado para a propagação do plasmídeo (por exemplo, DH5α, XL10 ouro) e mídia padrão adequado para a propagação de bactérias e seleção plasmídeo.
- Cepas de leveduras específicas, plasmídeos e primers, conforme descrito no protocolo.
- Geração de Iscas tMYTH por Repair Gap
- A isca deve ser clonado em um vetor apropriado para a marcação e expressão. Uma variedade de vetores tMYTH estão atualmente disponíveis para uso na construção isca. Vetores, como a pCMBV4, pAMBV4 pTMBV4 e permitir a construção de C-terminal iscas etiquetados (ISCA-C ub-LexA-VP16) sob o controle do CYC1 (fraca), ADH1 (forte) e TEF1 (muito forte) os promotores, respectivamente . Iscas N-terminal marcados (LexA-VP16-C ub ISCA) pode ser gerado através de vetores tais como pTLB-1 e pBT3-N, sob o controle do TEF1 e CYC1 promotores, respectivamente. A escolha do vetor depende da isca e deve ser determinada empiricamente. Em certos casos, a expressão maior isca é necessário para detectar interações, enquanto em outros casos superexpressão isca pode realmente ser prejudicial, levando a um aumento do número de falsos positivos.
- Restrição digerir o plasmídeo selecionado no sítio de restrição apropriada (s). Clivagem deve ocorrer apenas nas imediações da tag C ub-LexA-VP16 (upstream da tag para C-terminal de marcação ou a jusante de N-terminal tagging). Por exemplo, ao usar o vetor pAMBV4, SFII é uma escolha ideal. Armazenar o plasmídeo digerido a -20 ° C até que esteja pronto para uso.
- Primers projeto para a amplificação e clonagem do gene de interesse. Extremidade 5 'do seu Primer Encaminhar devem coincidir com aproximadamente 35-40 nucleotídeos upstream do sítio de restrição, enquanto que a extremidade 3' deve coincidir com o primeiro 18-20 nucleotídeos do gene alvo. Extremidade 5 'do primer reverso deve coincidir com o complemento reverso de aproximadamente 35-40 nucleotídeos a jusante do sítio de restrição, com a extremidade 3' correspondente ao complemento reverso da última 18-20 nucleotídeos do gene alvo (omitindo o códon de parada se o C ub-LexA-VP16 tag está sendo colocado no C-terminal). Dependendo se N-ou C-terminal de marcação está sendo realizado, selecione a 35-40 nucleotídeos do primer para a frente ou reversa de modo que o gene-alvo é clonado no quadro com a marca C ub-LexA-VP16.
- Amplificar o gene de interesse por PCR usando os primers acima. PCR parâmetros dependerá da enzima específica e primers específicos utilizados.
- Transformar o produto da PCR, juntamente com o plasmídeo digerido em uma cepa de levedura apropriadas laboratório tendo uma mutação leu2 (por exemplo, BY4741). Uma cepa repórter MITO (por exemplo, THY.AP4 ou L40) podem ser usados, mas não é necessário, como o propósito da levedura neste momento é simplesmente para servir como um ambiente em que a reparação gap recombinação homóloga podem ocorrer. Transformação deve ser realizada da seguinte forma:
- Inocular uma única colônia de sua cepa de levedura selecionada em 5 mL de mídia YPAD estéril e incubar durante a 30 ° C com agitação constante (200 rpm).
- Diluir a cultura durante a noite em 50 mL de mídia YPAD fresco para uma OD600 de ~ 0,15 e incubar a 30 ° C com agitação (200 rpm). Crescer por aproximadamente 3-4 horas até que um OD600 de ~ 0,6 é atingido.
- Centrifugar as células a 700xg por 5 minutos e remover o sobrenadante.
- Ressuspender o pellet celular em 25 mL estéreis DDH 2 O e centrifugar a 700xg por 5 minutos.
- Remover o sobrenadante e ressuspender o pellet celular em 1 mL estéreis DDH 2 O.
- Adicionar 100 mL de células, 300 mL de acetato mix PEG / Lithium, eo plasmídeo digerido (50 fmol) e PCR produto (250-500 fmol) para um tubo de microcentrífuga.
- Incubar a 30 ° C por 30 minutos.
- Choque térmico a 42 ° C por 1 hora.
- Centrifugar a 3000xg por 5 minutos e retire o sobrenadante.
- Ressuspender o pellet celular em 200 mL estéreis DDH 2 O e placa de todo o volume em sólido, SD-Leucina meios seletivos. Crescer a 30 ° C por 2-4 dias.
- Crescer uma única colônia da cepa transformada em 5 mL SD-Leucina meios líquidos a 30 ° C durante a noite.
- Centrifugar as células a 700xg por 5 minutos e remover o sobrenadante.
- Isolar DNA plasmídeo isca da pellets celulares usando qualquer kit miniprep comerciais. Seguir o protocolo padrão com uma modificação. A fim de garantir a lise celular suficiente fermento, adicionar um pequeno volume de 0,5 mm de refrigerante contas de vidro de cal para o pellet após ressuspensão inicial e vortex vigorosamente durante 5 minutos. Em seguida, proceder com o protocolo comercial como normal.
- Transformar DNA de levedura isolada em um E. competente coli cepa adequada para a propagação do plasmídeo (por exemplo, DH5α, XL10 Gold) com uma eficiência de transformação de pelo menos 1x10 7 células / DNA mg. Note-se que plasmídeos isca pode ser selecionado para a utilização de canamicina.
- DNA plasmídeo colheita da E. transformado coli através de um método padrão de isolamento de DNA ou kit comercial.
- Verificar a construção adequada da isca plasmídeo por seqüenciamento.
- Transformar a isca verificada em construir uma cepa adequada MITO repórter (por exemplo, THY.AP4, L40). O protocolo de transformação de levedura que acabamos de descrever pode ser utilizado, substituindo o DNA plasmídeo isca no lugar do produto digerido plasmídeo e PCR.
- Validação de isca - Localização adequada
- Antes do uso, cepas isca são analisados para assegurar que eles sejam devidamente localizado na membrana do fermento. Ao usar iMYTH, esta localização vai depender especificamente sobre as propriedades da isca marcados. Para tMYTH, plasmídeos isca geralmente incluem uma seqüência de sinais (por exemplo, Matα) dirigir a proteína expressa na membrana plasmática. Localização é determinada por microscopia de fluorescência. Inclusão de uma molécula YFP na seqüência tag isca (ie C ub-YFP-LexA-VP16) é a abordagem mais simples e direta, permitindo a visualização direta de células vivas, e é comumente usado em iMYTH. Alternativamente, uma abordagem padrão de imunofluorescência utilizando anticorpos contra o LexA ou VP16 componentes do tag pode ser usada.
- Validação de isca - N ub G / I Controle de Teste
- Localização, uma vez apropriada da isca foi estabelecido, é necessário garantir que a isca não ativar o sistema repórter sozinho ou na presença de não-interativas presas (isto é, verificar se a isca não é auto-ativação). Isto é realizado usando o N ub G / Teste I, onde a isca é transformada com a interação (positivo) e não-interativas presas (negativo), controle e crescimento é avaliado em meios seletivos. A isca deve crescer em meios seletivos in a presença do controlo positivo, e não crescer na presença do controle negativo, a fim de ser adequado para uso em MITO.
- Começar por transformar a tensão isca para ser testado com 100-200 ng de rapina controle plasmídeo. O protocolo de transformação descrito anteriormente levedura pode ser utilizado, substituindo SD-Leucina mídia no lugar de YPAD e usando SD-Triptofano-Leucina media para a etapa de revestimento final. As construções de controle presa seguintes são comumente usados:
- POST1-N ub I (Controle Positivo)
- POST1-N ub G (controle negativo)
- pFUR4-N ub I (Controle Positivo)
- pFUR4-N ub G (controle negativo)
- OST1 é um componente do complexo oligosaccharyl e está localizado na membrana retículo endoplasmático [9] enquanto FUR4 é uma permease uracil e está localizada na membrana plasmática [10]. Embora estas proteínas são geralmente a primeira escolha para o uso como não-interativas presas, a sua adequação irá variar caso a caso. No caso improvável de que a sua isca está previsto para realmente interagir com ambos os controles, presas alternativas terão de ser selecionado. Lembre-se que N ub I é a forma do tipo selvagem do N-terminal da ubiquitina, de forma espontânea e interage com C ub independente de uma interação entre as proteínas para o qual o ub ub C e N são fundidos. Assim presas N ub I constituem controles positivos, enquanto o N ub G presas (carregando a Isoleucina 13 a Glycine mutação que impede a associação espontânea de N ub ub e C) servir como controles negativos.
- Ressuspender colônias única de cada isca transformado em 100 mL estéreis DDH 2 O.
- Serialmente diluir as células ressuspendidas em DDH 2 O estéril para produzir diluições de 1 / 10, 1 / 100 e 1 / 1000.
- Spot 5 volumes mL de células não diluído e diluído em SD-Triptofano-Leucina e SD-Triptofano-Leucina-Adenina-Histidina mídia com e sem 3-AT em uma gama de concentrações. 3-AT age como um inibidor competitivo do gene repórter HIS3, e serve para aumentar o rigor do processo de seleção. Pode ser útil em alguns casos para inibir o crescimento não-específico de fraca a moderada auto-ativação iscas.
- Permitir que os pontos para secar e, em seguida, incubar as placas a 30 ° C por 2-4 dias.
- Todos os transformantes deve crescer nas placas SD-Triptofano-Leucina, indicando que eles foram transformado com sucesso com presas plasmídeo. Iscas que não ativar auto-vai crescer em SD-Triptofano-Leucina-Adenina-Histidina media apenas quando transformada com N ub eu presa constrói, e não com N ub G presas. Observe o que concentração de 3-AT é exigido na mídia (se houver), pois esta terá de ser usado durante a triagem.
4. Triagem
- Meios necessários e Soluções
- Estéril DDH 2 O preparado em autoclave a 121 ° C, 15 psi durante 30 minutos.
- Solução NaCl 0,9% preparado em DDH 2 O e esterilizados em autoclave a 121 ° C, 15 psi durante 30 minutos.
- Solução fosfato de sódio composto de 493 mM fosfato de sódio dibásico e 250 mM fosfato de sódio monobásico em DDH 2 O. Esterilizar em autoclave a 121 ° C, 15 psi durante 30 minutos.
- X-Gal (5-bromo-4-cloro-3-indolil-β-D-galactopiranosídeo) Solução preparada como uma de 100 mg / ml de solução em N, N-dimetil formamida.
- Mídia Crescimento 2xYPAD contendo extrato de levedura 2% w / v, 4% w / v de peptona, 4% de glicose w / v e 100 mM adenina, preparado em DDH 2 O. Esterilizar em autoclave a 121 ° C, 15 psi durante 30 minutos.
- Sintéticos Dropout (SD) Media Crescimento, preparado como descrito anteriormente. Prepare líquido SD-Leucina e sólido SD-Triptofano-Leucina. Despeje o meio sólido em ambos os pratos de 100x15 milímetros Petri. Prepare sólida SD-Triptofano-Leucina-Adenina-Histidina media em 150 milímetros rodada placas, 16 placas para cada tela, contendo 3-AT, se necessário, na concentração determinada a partir da ub N G / I teste de controlo.
- Sintéticos Dropout (SD) Media + 5-Bromo-4-cloro-3-β-Indoyl-D-galactopiranosídeo (X-Gal). Prepare SD-Triptofano-Leucina-Adenina-Histidina mídia contendo ágar, conforme descrito anteriormente. Após autoclavagem, deixe esfriar, acrescente 3-AT (se necessário), seguido de 1/10th o volume def solução de fosfato de sódio estéril. Em seguida, adicione X-Gal solução para uma concentração final de 80 mcg / mL. Misture bem e despeje em placas de 150 milímetros rodada.
- PEG / acetato de lítio Solução II contendo 40% de PEG-3350, 100 Acetato de Lítio mM, EDTA 1 mM e 10 mM Tris pH 7.5. Preparar esta solução usando estéril DDH 2 O e soluções (por exemplo, autoclavado 50% PEG-3350, um acetato de lítio M, 100 mM Tris pH 7,5 e 500 mM EDTA pH 8,0).
- Acetato de lítio / Solução Tris EDTA contendo 110 mM de acetato de lítio, 11 mM Tris pH 7,5 e 1,1 mM EDTA. Preparar esta solução usando estéril DDH 2 O e soluções (por exemplo, acetato de lítio autoclavado 1M, 100 mM Tris pH 7,5 e 500 mM EDTA pH 8,0).
- 10x Tris EDTA Solução composta por 100 mM Tris pH 7,5 e 10 mM EDTA preparado em DDH 2 O. Esterilizar em autoclave a 121 ° C, 15 psi durante 30 minutos.
- Solução single-stranded DNA Carrier (ssDNA), contendo 2 mg / mL de esperma de salmão sal de sódio DNA Tipo III, preparado em estéril DDH 2 O.
- Kit comercial miniprep.
- Cal sodada contas de vidro (0,5 mm).
- Escherichia coli competentes células adequado para a propagação do plasmídeo (por exemplo, DH5α, XL10 ouro) e mídia padrão adequado para a propagação de bactérias e seleção plasmídeo.
- Cepas de leveduras específicas e plasmídeos, conforme descrito no protocolo
- Transformação de Grande Escala
- Inocular uma única colônia da cepa repórter MITO contendo sua isca em 5 mL de SD-Leucina mídia e incubar durante a 30 ° C com agitação (200 rpm).
- Diluir a cultura overnight em 200 mL SD-Leucina de mídia a uma OD600 = 0,15 e incubar a 30 ° C com agitação (200 rpm). Crescer até que o OD600 = 0,6-0,7 (cerca de 4-5 horas).
- Pouco antes do OD600-alvo é atingido, descongelar uma alíquota de solução de ssDNA. Ferver a 100 ° C por 5 minutos e depois esfriar no gelo. Repetir uma vez.
- Quando o alvo OD600 foi atingido, a colheita das células através de centrifugação a 700xg por 5 minutos (divide a cultura mL 200 mL entre 4x50 tubos de centrífuga com tampa de rosca).
- Lave cada pellet com 30 mL estéreis DDH 2 O e brevemente vórtice da amostra. Centrifugar a 700xg por 5 minutos.
- Descartar o sobrenadante e ressuspender cada pellet em 1 Acetato de Lítio mL / solução Tris EDTA. Transferir para um tubo de microcentrífuga estéril 1,5 ml e centrifugar a 700xg por 5 minutos.
- Descartar o sobrenadante e ressuspender cada pellet em 600 mL de Acetato de Lítio / solução Tris EDTA.
- Adicione o seguinte ao 4x15 mL tubos de tampa de rosca de centrífuga:
- 2,5 mL PEG / acetato de lítio Solução II
- 600 mL de células ressuspenso
- 100 mL de solução ssDNA
- 7 mg de DNA biblioteca presa
- Bibliotecas contendo tagged presas em uma ou outra o N-ou C-terminal com N ub G, e preparados a partir de uma variedade de fontes de cDNA ou genômica, estão disponíveis comercialmente ( www.dualsystems.com ). A biblioteca específico utilizado deve ser determinada numa base caso a caso, depende da isca selecionada e objetivos experimentais.
- Vortex por 1 minuto para assegurar a mistura completa e, em seguida, incubar em um banho-maria 30 ° C por 45 minutos. Mix brevemente a cada 15 minutos.
- Adicionar 160 mL dimetil sulfóxido (DMSO) a cada tubo e misturar imediatamente invertendo os tubos.
- Incubar em banho-maria 42 ° C por 20 minutos.
- Coletar transformantes por centrifugação a 700xg por 5 minutos.
- Desprezar o sobrenadante. Recuperar transformantes por ressuspensão de cada pellet em 3 2xYPAD mL. Piscina todas as amostras juntas em um único tubo de centrífuga de 50 mL com tampa de rosca.
- Incubar a 30 ° C por 90 minutos para a recuperação das células.
- Centrifugar a 700xg por 5 minutos e desprezar o sobrenadante.
- Ressuspender o pellets de células em 4,9 mL de NaCl 0,9% estéril.
- Utilizando 100 mL de células ressuspenso preparar 10 diluições de série em estéril de NaCl 0,9%, variando de 10x a 1000x.
- ML placa 100 das diluições 100x e 1000x para SD-Triptofano-Leucina mídia e incubar a 30 ° C por 2-3 dias. Estas placas servem como um controle e são usados para calcular a eficiência da transformação.
- Dividir igualmente os restantes 4,8 mL de células ressuspenso e placa em grandes (150 mm) SD-Triptofano-Leucina-Adenina-Histidina placas,contendo a quantidade necessária de 3-AT, conforme determinado na ub N G / I de teste, e incubar a 30 ° C por 3-4 dias.
- Ressuspender colônias individuais (cada uma representando as células que contêm um potencial de interação par isca presa) em 100 mL de NaCl 0,9% e placa de 5 mL em alíquotas SD-Triptofano-Leucina-Adenina-Histidina + X-Gal mídia (incluindo 3-AT se necessário). Permitir a crescer por 2-4 dias. Esta etapa serve como uma segunda rodada de análise selectiva, e ajuda na remoção de falsos positivos obtidos na rodada inicial. Colônias só que exibem um crescimento robusto e uma cor azul são selecionados para análise posterior.
- Isolamento e sequenciamento de DNA presa
- Isolar DNA plasmidial das colônias de leveduras azul usando um protocolo miniprep com as modificações descritas anteriormente. Certifique-se de crescer as células em SD-Triptofano somente mídia, para selecionar para a retenção de rapina, mas não isca, plasmídeos. Para telas que produzem um número muito grande de visitas, um disponível comercialmente de alto rendimento kit miniprep pode ser vantajosa neste momento.
- Transformar DNA de levedura isolada do plasmídeo em um E. competente coli cepa adequada para a propagação do plasmídeo (por exemplo, DH5α, XL10 Gold) com uma eficiência de transformação de pelo menos 1x10 7 células / DNA mg. Note-se que plasmídeos rapina pode ser selecionado para a utilização de ampicilina.
- Isolar o DNA do plasmídeo E. transformado coli através de um método padrão de isolamento de DNA ou kit comercial. Mais uma vez, um alto rendimento kit miniprep pode ser útil se o número da amostra é grande. A amplificação do DNA em E. coli aumenta o rendimento plasmídeo e garante que uma quantidade suficiente de DNA está presente tanto para seqüenciamento e análise posterior.
- Seqüência os plasmídeos isolados usando um primer complementar à seqüência dentro do ub N G.
- Compilar e analisar todos os dados de seqüenciamento de montar sua lista preliminar dos interagentes. Isso pode ser feito manualmente ou de forma automatizada usando software apropriado.
- Teste de Dependência Bait
- Após a montagem da lista preliminar de interatores é importante verificar novamente as interações e eliminar presas promíscua que interagem / ativar o sistema repórter de uma maneira independente da identidade isca. Isto é realizado utilizando o Teste de Dependência Bait. Neste teste, todos os interatores identificados são transformados de volta para a cepa isca original, bem como uma cepa de abrigar uma isca artificial de controle consiste em um único domínio transmembrana fundidos para a marca C ub-LexA-VP16. Transformação é realizada conforme o protocolo padrão descrito anteriormente, utilizando SD-Leucina mídia no lugar de YPAD e SD-Triptofano-Leucina meios sólidos para a etapa de revestimento final.
- Ressuspender colônias única das transformações acima em 100 mL de estéril DDH 2 O 5 volumes e local mL para SD-Triptofano-Leucina-Adenina-Histidina + X-Gal mídia (incluindo 3-AT se necessário). Placas são então incubadas por 2-4 dias a 30 ° C. Idealmente transformantes múltiplos devem ser selecionados para cada presa, e ambas as iscas o original e isca artificial deve ser visto sobre o mesmo prato.
- Levedura levando a isca artificial e presas que a ativação causa do sistema de repórter (ou seja, o crescimento ea cor azul) são consideradas promíscuas e que presa específico é removido da lista de interatores.
- Presas que causam o crescimento e coloração azul na levedura com a isca de interesse, mas não a isco artificial, confirma essa interação específica. Se, no entanto, fermento abrigar as presas e sua isca de interesse não crescem, esta presa é removido da lista de interatores.
- As presas restantes constituem a lista completa de interatores identificado na tela MITO.
5. Estudos mais
Uma vez triagem MITO foi concluída, novas análises devem ser realizadas a fim de validar e determinar o significado biológico das interações detectados. Os estudos específicos a realizar irá variar em uma base caso a caso, e deve ser determinada pelo pesquisador individual. Alguns exemplos comuns de trabalho de acompanhamento incluem co-imunoprecipitação experiências e estudos de eliminação no organismo nativa. Além disso, análise computacional dos dados obtidos podem ser úteis para a detecção de padrões, e ajudando a identificar a relevância potencial e papel diferentes interações pode jogar. Assim, a tecnologia MITO serve como um "passo-a primeiro poderosa para a identificação e compreensão das interacções críticas funcionais de proteínas da membrana. Juntamente com detalhados estudos de acompanhamento e outros te recentemente desenvolvidos e emergenteschnologies, que promete ser uma ferramenta valiosa para desvendar os mistérios da célula.

Figura 1. Princípio da separação ubiquitina. A. Ubiquitina pode ser separado em duas metades: a metade C-terminal (C ub) ea metade N-terminal (N ub I). Estas metades reconstituir espontaneamente por causa de sua alta afinidade por um outro. b. AN ub mutação pontual na posição 13 I de uma isoleucina a glicina (N ub G) impede que esta espontânea re-associação. c. No sistema MITO, o ub C é fundido para a isca de interesse (B) ea presa é fundida ao ub N G (A). AB interação proteína reconstitui pseudo-ubiquitina.
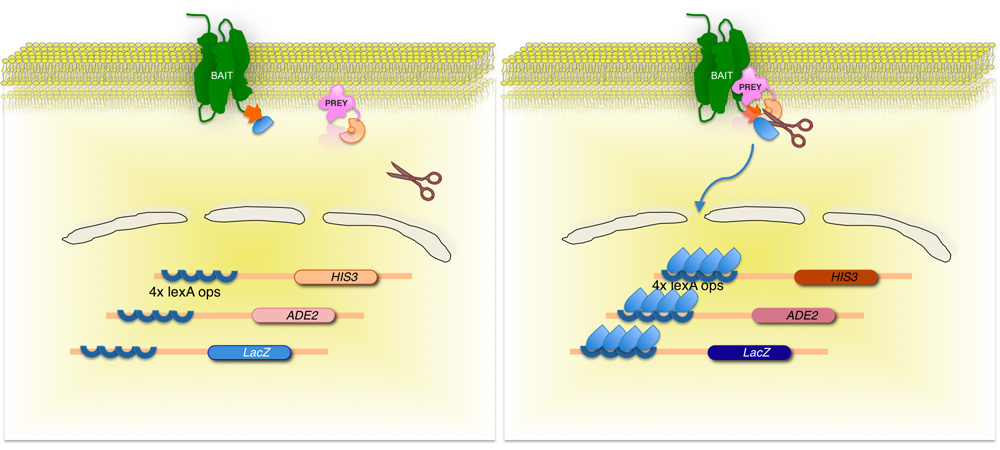
Figura 2. Split-ubiquitina levedura membrana baseada dois híbridos (MITO) do sistema.
A membrana de proteína de interesse (isca) é fundido para a metade C-terminal de levedura ubiquitina (Ub C), conjugado com um fator de transcrição. Usando uma biblioteca de cDNA ou gDNA, cada proteína codificada pela biblioteca (presa) é fundida ao correspondente N-terminal da molécula de ubiquitina (Ub N G). Se as duas proteínas não interagem, o fator de transcrição permanece na interface membrana (painel esquerdo). No entanto, se as proteínas interagem, as duas metades da ubiquitina participar, resultando em clivagem pela ubiquitina proteases específicas. Clivagem libera o fator de transcrição, resultando na expressão de genes repórter (painel direito)

Figura 3. Pipeline MITO.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
MITO é o sistema de débito primeira alta que permite a identificação de interações entre proteínas de longa-metragem de membrana e citosólica ou ligada à membrana parceiros. Ela tem sido usada para estudar proteínas de membrana de uma série de organismos [3-7]. Há, no entanto, detalhes específicos que podem precisar de ser examinado para assegurar a proteína de interesse é passível de estudo com MITO.
Muitos ligada à membrana proteínas são direcionadas para a membrana plasmática através de uma seqüência de sinal que é posteriormente clivado para produzir a proteína madura. Esta sequência é-organismo específico e é possível que esta seqüência de sinal nativo causar um mal-localização da proteína de interesse, quando expressa em levedura porque a seqüência de sinal permanece desconhecida. Para contornar esse problema, nós engenharia estas proteínas específicas para ser fundido com a seqüência de sinal de levedura, derivado do fator alfa de acoplamento (MATα). Esta seqüência de peptídeos (os chamados MFα-ss) re-localiza a proteína para a membrana plasmática de levedura e importante, é clivada por peptidases sinal de levedura. Esta seqüência peptídeo é encontrado em plasmídeos pTMBV-MFα e pAMBV-MFα.
Outro parâmetro importante que precisa ser enfatizado é níveis de expressão isca. O promotor que dirige a expressão da isca regula este parâmetro. Pode ser necessário para otimizar os níveis de isca expressão usando o teste NubG / Nubi ea quantidade de 3-AT necessárias para eliminar os artefatos superexpressão, onde a isca é "auto-ativação" (ou seja, promiscuamente interage com muitas proteínas não-específicas presas) . Examinar proteínas de levedura produz as concentrações isca mais fisiologicamente relevante quando iMYTH é aplicada. Neste caso, o gene de interesse é marcado com o Cub-TF dentro do gDNA. Alternativamente, proteínas exógenas pode ser expressa de pBT3-STE e plasmídeos pCMBV que carregam o promotor CYC1 resultando na expressão bait baixo. O pTMBV plasmídeos e pTLB1 porto promotor TEF1 enquanto pAMBV tem o promotor ADH1, tanto a expressão que forte impulso da proteína isca. Se os níveis de proteína isca requerem maior optimização, pode ser necessário usar o plasmídeo pTLB-1, que transporta o promotor TEF1, no entanto, o domínio DNA LexA ligação é mutante em R156G a diminuir a afinidade para com os promotores de genes exógenos repórter, em última análise, diminuindo o probabilidade de auto-ativação [5].
Outro fator que desempenha um papel importante para o sucesso MITO é a biblioteca de escolha utilizado para o processo de triagem. Isso vai depender de os perfis de expressão endógena isca. Por exemplo, a isca pode ser expressa em tecidos específicos, e por isso é importante o uso de uma biblioteca que é construído a partir deste tecido específico. Isso irá garantir fisiologicamente interações relevantes são detectados.
O sistema MITO é uma ferramenta simples e rápida, que oferece uma abundância de informações sobre uma classe de proteínas que têm sido difícil de estudar. Essas interações identificadas podem auxiliar na elucidação da função biológica completa de proteínas da membrana.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Igor Stagljar é um co-fundador da DualSystems Biotech, Suíça.
Acknowledgments
Gostaríamos de agradecer Edmonds Alvorada para uma leitura crítica deste manuscrito. O laboratório Stagljar é apoiado por fundos da Fundação Canadense para Inovação (CFI), o Instituto Canadense de Pesquisa em Saúde (CIHR), a Fundação do Coração e Derrame, da Sociedade Canadense do Câncer, e Novartis.
Materials
| Name | Company | Catalog Number | Comments |
| Polyethenlene Glycol (PEG3350) | BioShop Canada | PEG335 | |
| Lithium Acetate Bihydrate | BioShop Canada | LIA001 | |
| X-Gal (5-Bromo-4-Chloro-3-Indolyl-b-D-galactopyranoside) | BioShop Canada | XGA001 | |
| N`,N-dimethyl formamide | BioShop Canada | DMF 451 | |
| 3-amino-1,2,4-triazole (3-AT) | BioShop Canada | ATT124 | |
| Sodium phosphate dibasic | BioShop Canada | SPD307 | |
| Sodium phosphate monobasic | Fisher Scientific | BP329-500 | |
| Salmon Sperm DNA | VWR international | CA80601-120 | |
| D-Glucose | BioShop Canada | GLU501 | |
| LB Broth LENOX | BioShop Canada | LBL405 | |
| Yeast Nitrogen Base | BioShop Canada | YNB406 | |
| Yeast Extract | BioShop Canada | YEX401 | |
| Peptone | BD Biosciences | 211677 | |
| Bio-Tryptone | BioShop Canada | TRP402 | |
| Adenine Sulphate | BioShop Canada | ADS201 | |
| L-Uracil | BioShop Canada | URA241 | |
| L-Threonine | BioShop Canada | THR002 | |
| L-Histidine | BioShop Canada | HIS200 | |
| L-Methionine | BioShop Canada | MET222 | |
| L-Valine | BioShop Canada | VAL201 | |
| L-Phenylalanine | BioShop Canada | PHA302 | |
| L-Isoleucine | BioShop Canada | ISO910 | |
| L-Tyrosine | BioShop Canada | TYR333 | |
| L-Leucine | BioShop Canada | LEU222 | |
| L-Arginine | BioShop Canada | ARG006 | |
| L-Tryptophane | Fisher Scientific | BP395-100 | |
| L-Lysine | BioShop Canada | LYS101 | |
| L-Alanine | Fisher Scientific | BP369-100 | |
| Agar | BioShop Canada | AGR001 | |
| Soda Lime Galss Beads | Biospec Products | 11079105 | |
| Sodium Chloride | BioShop Canada | SLD002 |
References
- Stagljar, I., Fields, S. Analysis of membrane protein interactions using yeast-based technologies. Trends Biochem Sci. 27 (11), 559-563 (2002).
- Iyer, K. Utilizing the split-ubiquitin membrane yeast two-hybrid system to identify protein-protein interactions of integral membrane proteins. Sci STKE. 275, pl3-pl3 (2005).
- Paumi, C. M. Mapping protein-protein interactions for the yeast ABC transporter Ycf1p by integrated split-ubiquitin membrane yeast two-hybrid analysis. Mol Cell. 26 (1), 15-25 (2007).
- Stagljar, I. A genetic system based on split-ubiquitin for the analysis of interactions between membrane proteins in vivo. Proc Natl Acad Sci U S A. 95 (9), 5187-5192 (1998).
- Gisler, S. M. Monitoring protein-protein interactions between the mammalian integral membrane transporters and PDZ-interacting partners using a modified split-ubiquitin membrane yeast two-hybrid system. Mol Cell Proteomics. 7 (7), 1362-1377 (2008).
- Scheper, W. Coordination of N-glycosylation and protein translocation across the endoplasmic reticulum membrane by Sss1 protein. J Biol Chem. 278 (39), 37998-38003 (2003).
- Thaminy, S. Identification of novel ErbB3-interacting factors using the split-ubiquitin membrane yeast two-hybrid system. Genome Res. 13 (7), 1744-1753 (2003).
- Johnsson, N., Varshavsky, A. Split ubiquitin as a sensor of protein interactions in vivo. Proc Natl Acad Sci U S A. 91 (22), 10340-10344 (1994).
- Kelleher, D. J., Gilmore, R. The Saccharomyces cerevisiae oligosaccharyltransferase is a protein complex composed of Wbp1p, Swp1p, and four additional polypeptides. J Biol Chem. 269 (17), 12908-12917 (1994).
- Chevallier, M. R. Cloning and transcriptional control of a eucaryotic permease gene. Mol Cell Biol. 2 (8), 977-984 (1982).
