

Summary
在这里,我们展示了单分子荧光显微镜,对生活的细菌细胞,使功能分子复合物进行检测,跟踪和量化的执行协议。
Abstract
全面洞察到活细胞的机制,可以实现只有通过调查的关键工序引出并直接在细胞水平上的事件。迄今为止的生物系统的剪切复杂性造成了精确的单分子实验过于苛刻,而不是单一系统使用相对粗糙的散装合奏平均测量的研究重点。然而,许多重要的过程发生在活细胞只是一个水平或几个分子;合奏测量一般掩盖这些事件的随机性和异质性。在这里,使用先进的光学显微镜和分析图像分析工具,我们演示了如何在一个单一的生活细菌细胞的单个分子的精确度,以及如何运作的生物机器,我们可以观察到分子复合物内的动态监测的蛋白质。该技术直接相关的生理。他们微创微扰和非侵入性正在研究的生物样品,并完全合拍,在物质生活的调查功能,没有现成的其他单分子生物物理的方法。此外,生物标本研究所有在未修改的细胞株(“基因组编码”),产生显着更多的蛋白质比自然发生的,而不是比较常见的,但不太理想的方法几乎相同的水平产生荧光标记的蛋白质(“质粒表达”)。因此,将调查的实际生物样品中有显着接近自然的有机体,因此观测真正的生理过程有关。
Protocol
- 要开始此过程中,50μL荧光蛋白表达大肠杆菌细菌细胞冷冻库存首次解冻和成长翻天覆地的5毫升LB生长介质一夜之间在37度C.在早上有氧,提取50μL这个饱和文化和子成最小的M63葡萄糖培养基,在30度4至6小时的彗星孵化,培养。在这里,我们展示了使用两种不同的细胞株,其中一个表示电子传输的细胞色素融合到绿色荧光蛋白,表示有关的蛋白质融合,以绿色荧光蛋白在细菌鞭毛电机的其他。
- 细胞可收获直接从不断增长的亚文化,如果他们作为固定样本观看,或他们可能会剪截断细菌鞭毛“拴”条件下观看。
- 剪切涉及配售的亚文化通常为1 -5毫升成两个无菌注射器,无菌管道加入的设备。剪切是通过在每个注射泵交替推推动通过狭窄的管文化。这是50-100倍,所需的剪切程度而定。文化是离心沉淀细胞,它是悬浮在最短的媒体删除鞭毛片段。
- 然后,我们准备在氢氧化钾饱和溶液浸泡20分钟乙醇清洗BK7玻璃盖玻片,然后在去离子水和乙醇和漂洗彻底离开干燥的空气中至少有1小时
- 我们构造一个简单的流细胞内部的细胞,在显微镜。这涉及到一个BK7玻璃显微镜幻灯片上绘制石蜡油脂线,然后建立一个隧道,夹层置于顶部的清洗coverlips之一,并与一双镊子轻轻按下,让流细胞体积的5-10μL 。
- 观察固定化细胞注射填充流动细胞与聚- L -赖氨酸0.1%W / V解决方案,并允许它在室温下孵育至少1分钟。然后,我们冲洗出来使用100μL最小的媒体,通过注入流通池的一端从其他媒体和排汗用纸巾。
- 然后,我们通过一个直径为200纳米乳胶微球(Polysciences)1:500稀释在最短的媒体,以纪念盖玻片表面的20μL灯芯。流通池倒盖玻片朝下,放置在一个平台上清晰的表面,在一个简单的湿度室,并在室温下孵育5 min。未绑定的珠子,然后通过100μL最小媒体排汗冲走。
- 如果我们想观察拴细胞,我们省略了聚- L -赖氨酸的一步,而不是填补流动细胞与5 mg / ml的抗鞭毛蛋白抗体。流通池是在10分钟的湿度室的地方,然后通过排汗刷新。
- 20细胞培养液,然后恶人通过流通池,可以使用剪切样品,如果观察拴细胞固定化细胞在观看或unsheared样品。
- 倒流通池,并放置在湿度为20分钟室。未绑定的细胞,然后通过100μL最小媒体排汗冲了出去。
- 浸油下降是摆在盖玻片的顶面中心和流通池,然后轻轻地放在定制荧光显微镜的样品架,使高数值孔径物镜的光学接触。
- 显微镜电子倍增接通相机和相机设置被冷却到-70摄氏度,该软件是掌握在一个典型的帧速率,帧传输模式,在25赫兹的图像,最初disenabling上的增益控制摄像头。
- 明照明开启和图像带入重点,选择一个合适的细胞或细胞组,他们被卡,其长轴平行的盖玻片表面的基础上成像。重点是微调,以确保200纳米乳胶珠坚持盖玻片表面,只是在重点。
- 图像序列是在明收购记录的细胞体的轮廓。明照明关闭和启用摄像机的增益为最大。
- 对于收购标准,使用全内反射荧光(TIRF)通过适当波长的激光束(在这里,绿色荧光蛋白的激发,我们使用了473纳米激光)是预先设定,以便集中后焦平面客观的镜头,但流离失所光轴横向到细胞样品的荧光激发产生渐逝场。
- 相机采集开始,和激光快门打开激发荧光蛋白的细菌内。收购的激光强度和速度的参数需要尝试不同的价值观,特别是在调查的生物系统进行了优化,而是一个典型的范围relevant研究细胞膜的移动蛋白复合物的1-10毫瓦激光功率超过一个直径1〜30微米的圆形激发面积,每帧5-40毫秒的曝光时间。样品被点亮,直到光漂白,通常为10秒
- 都拴细胞,在细胞体旋转的附着点之间的盖玻片和鞭毛存根和固定化细胞固定盖玻片表面,可以使用此协议。
- 有一个复合物的高拷贝数的一些细胞株细胞TIRF成像之前的一极的重点从最初的衍射极限的激光漂白受益。这种漂白后荧光恢复使用漂白(FRAP),相当于。使用我们的自定义显微镜激发激光灯可喂到第二个独立使用的路径为FRAP型漂白。通常情况下使用1 - 10mW的激光功率,一个典型的漂白时间范围在10-300毫秒。这样的结果要高得多,在允许个别的复合物,随后进入该地区扩散到更容易地可视化细胞漂白区造影。
- 复合物在细胞浆内扩散速度远远比在细胞膜上采用不同的照明模式,所谓的“slimfield”的可视化。这里聚焦的激光是横向扩展,只包括一个单细胞。这将产生一个非常激烈的领域,允许采取风险通常毫秒快得多。
- 经过数据采集,图像送入自定义编写的软件(在LabVIEW 8.5中的编码)。这会自动检测在一个典型的几个纳米的精度细胞的荧光斑点的位置,并提取它们的大小和亮度。 brightnessof漂白跟踪跟踪的分子复合物的时间是用来估计化学计量的,即弥补了单分子复杂多少个人的荧光蛋白质。
代表性的成果:
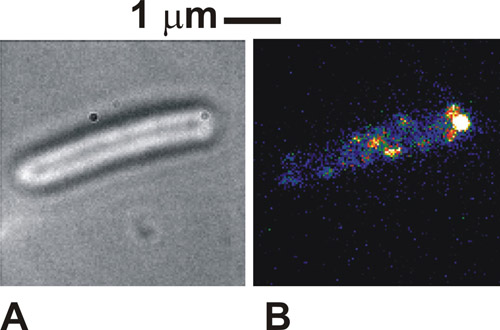
当该协议是正确的细胞在明看的图像是非常明显的,对一个白色/灰色的细胞体(图1A)黑暗的细胞体的周长。在荧光采用固定化细胞,我们可以看到踩点的强度,一般宽度在250-300纳米(图1b)。将被视为健康,拴细胞绕明图像点的系绳附件。在我们的例子中的一些分子络合物在荧光激发下也可能看到的附着点,说明标签蛋白与鞭毛马达的本地化。这些斑点是个别分子络合物,他们看到的数量将取决于所使用的照明模式,以及如何配合物的许多实际上存在于细胞内,在任何一个时间。点的流动性取决于下的具体研究生物系统。如果斑点的密度最初非常高,因为这里使用的标记细胞色素的情况下,然后执行初始FRAP还原漂白剂可以提高成像的对比度。

图1。 (一)明场和(二)TIRF图像(假色)为一个固定的大肠杆菌细胞表达的一种蛋白质融合,这是众所周知的细菌鞭毛马达的绿色荧光蛋白(GFP )的。请点击这里看到图1的放大版本。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
必须小心不要“剪切”拴细菌细胞,因为这可能会损害鞭毛马达的功能。重要的是使用细胞的显微镜玻片上一次更长的时间不到一个小时,因为他们可能成为氧气耗尽。可能需要相当大的优化,正在调查中找到最好的显微镜成像条件满足您特定的生物系统。这可能是明智的做法是企图用纯化GFP的单独确定正确的强度激光激发您的特殊的显微镜系统所需的成像。
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
我们承认,从教授朱迪思阿米蒂奇(英国牛津大学)和康拉德Mullineaux教授(英国伦敦大学玛丽皇后学院)组菌株的实物捐赠。 IMD是共同出资的生物化学部(牛津大学)和OCISB; AR是由工程和物理科学研究理事会(EPSRC)的DTC的助学金的资助; ND是从生物技术和生物科学研究理事会(BBSRC)资助; MCL由英国皇家学会大学研究奖学金。
References
- Leake, M. C., Chandler, J. H., Wadhams, G. H., Bai, F., Berry, R. M., Armitage, J. P. Stoichiometry and turnover in single, functioning membrane protein complexes. Nature. 443, 355-358 (2006).
- Leake, M. C., Greene, N. P., Godun, R. M., Granjon, T., Buchanan, G., Chen, S., Berry, R. M., Palmer, T., Berks, B. C. Variable stoichiometry of the TatA component of the twin-arginine protein transport system observed by in vivo single-molecule imaging. Proc Natl Acad Sci U S A. 105, 15376-15381 (2008).
- Leake, M. C., Wilson, D., Gautel, M., Simmons, R. M., M, R. The elasticity of single titin molecules using a two-bead optical tweezers assay. Biophys. J. 87, 1112-1135 (2004).
- Leake, M. C., Wilson, D., Bullard, B., Simmons, R. M. The elasticity of single kettin molecules using a two-bead laser-tweezers assay. FEBS Lett. , 535-555 (2003).
- Lenn, T., Leake, M. C., Mullineaux, C. W. In vivo clustering and dynamics of cytochrome bd complexes in the Escherichia coli plasma membrane. Mol. Microbiol. 70, 1397-1407 (2008).
- Lenn, T., Leake, M. C., Mullineaux, C. W. Are Escherichia coli OXPHOS complexes concentrated in specialised zones within the plasma membrane. Biochem. Soc. Trans. 36, 1032-1036 (2008).
- Lo, C. J., Leake, M. C., Pilizota, T., Berry, R. M. Single-cell measurements of Membrane Potential, Sodium-Motive Force and Flagellar Motor Speed in Escherichia coli. Biophys. 93, 294-302 (2007).
- Lo, C. J., Leake, M. C., Berry, R. M. Fluorescence measurement of intracellular sodium concentration in single Escherichia coli cells. Biophys. J. 90, 357-3565 (2006).

{kind=link}