

Summary
Här kan vi visa protokoll för att utföra en enda molekyl fluorescensmikroskopi på levande bakterieceller att funktionella molekylära komplex som ska upptäckas, spåras och kvantifieras.
Abstract
Full insyn i de mekanismer i levande celler kan endast uppnås genom att undersöka viktiga processer som framkallar och direkta händelser på cellnivå. Hittills skjuvning komplexiteten i biologiska system har orsakat exakt enda molekyl experiment för att vara alltför krävande, i stället för att fokusera på studier av enskilda system som använder relativt grova bulk ensemble-genomsnittet mätningar. Men många viktiga processer sker i den levande cellen i nivå med bara en eller ett fåtal molekyler, ensemble mätningar generellt maskera stokastiska och heterogena dessa händelser. Här med hjälp av avancerad optisk mikroskopi och analytiska bild analysverktyg vi visar hur man kan övervaka proteiner i en enda levande bakterie med en precision av enstaka molekyler och hur vi kan observera dynamiken inom molekylär komplex i fungerande biologiska maskiner. De tekniker som är direkt relevanta fysiologiskt. De är minimalt perturbative och icke-invasiv till biologiskt prov som undersöks och är fullt anpassad för utredningar i levande material, funktioner som inte är tillgängliga för andra en-molekyl strategierna i biofysik. Dessutom studerade biologiska prover alla producerar fluorescerande-märkta proteinet på nivåer som är nästan identiska med omodifierade celler stammar ("genomisk kodning"), i motsats till den vanligare men mindre bra metod för att generera betydligt mer protein än vad som förekommer naturligt (plasmid uttrycket "). Således, den verkliga biologiska prover som kommer att undersökas är betydligt närmare den naturliga organismer, och därför iakttagelser mer relevanta för verkliga fysiologiska processer.
Protocol
- Till att börja den här proceduren är 50 l fryst lager av fluorescerande proteinet uttrycka Escherichia coli bakterieceller first tinas och vuxit aerobt med skakning i 5 tillväxt ml LB media över natten vid 37 grader C. På morgonen är 50 l av denna mättade kultur extraherade och sub-odlade i minimal M63 media glukos kultur, inkubering i 30 grader C i 4 till 6 timmar. Här visar vi två olika cell-stammar, av vilka uttrycker en elektron-transport cytokrom smält till GFP, den andra som uttrycker ett protein som deltar i den bakteriella flagellar motorn smält till GFP.
- Celler kan antingen skördas direkt från växande sub-kultur om att se dem som immobiliseras prover, eller de kan vara klippt för att trunkera bakteriella flageller om visning under "bundna" villkor.
- Klippning innebär att placera vanligtvis 1-5ml av sub-kultur till en apparat som består av två sterila sprutor sällskap av sterila rör. Det klippning sker genom att växla trycka i varje sprutpump att driva kulturen genom den smala slangen. Detta görs 50-100 gånger, beroende på omfattningen av nödvändiga klippning. Kulturen är centrifugeras sedan till pellets cellerna, som är suspenderade i minimal media att ta bort flageller fragment.
- Vi sedan förbereda rengöras BK7 glas täckglas genom att sänka ner i en mättad lösning av KOH i etanol i 20 minuter, sedan skölj noga i avjoniserat vatten och etanol och lämnar torka i luften i minst 1 h.
- Vi konstruerar en enkel flow-cellen till hus cellerna i mikroskop. Detta innebär att rita linjer av paraffin fett på en BK7 glas objektsglas och sedan skapa en tunnel-sandwich genom att placera en av de rengöras coverlips ovanpå och trycka ner försiktigt med en pincett, vilket ger ett volymflöde cell på 5-10 l .
- För att observera immobiliserade celler vi fyller flödet-cellen genom injektion med 0,1% w / v lösning av poly-L-lysin, och låt den inkubera i rumstemperatur i minst 1 min. Vi spola därefter ut med 100 l minimal medier genom att injicera media från ena änden av flödes-cell och fuktspridande med silkespapper från den andra.
- Vi veken sedan genom 20 ìl av en 1:500 utspädning av 200 nm diameter latex mikrosfärer (Polysciences) i minimala media för att markera täckglaset ytan. Flödet-cellen är inverterad så att täckglaset är vänd nedåt, placeras på en plattform tydligt av ytan på ett enkelt fuktkammare, och inkuberas i rumstemperatur i 5 minuter. Obundet pärlor är sedan tvättas bort av fuktspridande genom 100 l minimal medier.
- Om vi vill följa uppbundna celler vi utelämnar poly-L-lysin steg och istället fylla den flow-cellen med 5 mg / ml anti-flagellin antikropp. Flödet-cell plats i fuktkammare i 10 min och sedan spolas igenom av fuktspridande.
- 20 ìl av cellodling är då ond genom flödes-cellen, antingen via klippt provet om att observera bundna celler eller unsheared provet i visning immobiliserade celler.
- Flödet-cellen är inverterad och placeras i fuktig kammare i 20 min. Obundet cellerna tvättas ut av fuktspridande genom 100 l minimal medier.
- En droppe immersionsolja är placerad i mitten av den översta ytan av täckglas och flödet-cellen placeras sedan försiktigt på till provet innehavaren av den inbyggda anpassade fluorescensmikroskop, vilket gör optisk kontakt med hög numerisk apertur objektiv.
- Mikroskopet elektron-multiplicera kameran är påslagen och kameran inställd på att kylas till -70 grader C, är programvaran inställd på att få bilder på en typisk bildfrekvens på 25 Hz i ram-överföringsläge, inledningsvis disenabling att få kontroll på kameran.
- Den brightfield belysningen slås på och bilden är i fokus, välja en lämplig cell eller grupp av celler som skall avbildas på grund av att de är fast med sina långa axel parallell med täckglaset ytan. Inriktningen är finjusteras för att säkerställa att 200 nm latex kulor fastnat på täckglas ytan är bara i fokus.
- En bildsekvens förvärvas i brightfield att spela in konturerna av cellkroppen. Den brightfield belysningen är avstängd och kameran vinsten är aktiverat för att maximalt.
- För en standard förvärv med total inre reflektion fluorescens (eller TIRF) en laserstråle över lämplig våglängd (här, för grönt fluorescerande protein excitation, vi använder en 473 nm laser) är förinställd för att vara fokuserad på baksidan fokalplan i objektivet, men förskjuts i sidled från optiska axeln för att generera en försvinnande fält för fluorescens excitation i cellen provet.
- Kameran Förvärvet är igång och lasern slutaren öppnas för att excitera det fluorescerande proteiner i bakterier. Parametrarna för laser intensitet och hastighet förvärv behöver optimeras för den specifika biologiska systemet är under utredning genom att experimentera med olika värden, men ett typiskt område rElevant att studera mobila proteinkomplex i cellmembranet är 1-10 mW laser makt över en cirkulär excitation område med en diameter på ca 30 mikrometer och en exponeringstid per ram för 5-40 ms. Prover upplyst fram photobleached, typiskt för ~ 10 s.
- Detta protokoll kan användas både för tjudrad celler där kroppen av cellen roterar kring en fästpunkt mellan täckglas och en flagellar stub, och för immobiliserade celler som strikt är fast på täckglaset ytan.
- Vissa celler stammar som har en hög Antal kopior av komplex nytta av en inledande diffraktion-begränsad fokuserad laser blekning på en pol av cellen före TIRF avbildning. Detta blekmedel motsvarar den som används i fluorescens återhämtning efter fotoblekning (eller FRAP). Med vår egen mikroskop några av excitation laserljus kan matas ut i en andra oberoende väg för FRAP-typ blekning. Typiskt 1-10mW laser effekt används, med en typisk blekmedel i intervallet 10-300 ms. Detta resulterar i mycket högre avbildning kontrast i blekt zon cellen låta enskilda komplex som därefter diffus in i detta område att vara lättare visualiseras.
- För att visualisera komplex i cellens cytoplasma som diffus mycket snabbare än de i cellmembranet en annan belysning läge som kallas "slimfield" är anställd. Här en fokuserad laser utökas i sidled för att bara omfatta en enda cell. Detta ger en mycket intensiv fält så mycket snabbare exponeringar typiskt en millisekund som skall vidtas.
- Efter datainsamling, är de bilder som matas in anpassade skriven mjukvara (kodade i LabVIEW 8,5). Detta upptäcker automatiskt positioner fluorescerande fläckar i cellerna med en precision av ett typiskt några nanometer och utdrag ur deras storlek och ljusstyrka. Den brightnessof den fotoblekning spår med avseende på tiden för en spårad Molecular Complex används sedan för att uppskatta stökiometri, dvs hur många enskilda fluorescerande proteiner bildar en enda molekylära komplex.
Representativa resultat:
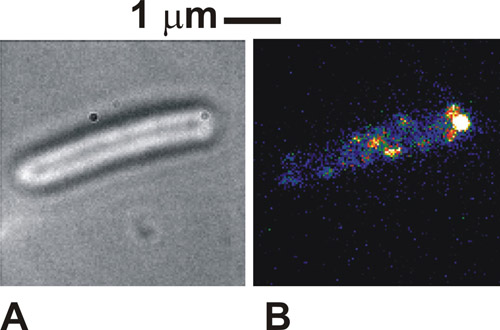
När protokollet är gjort på rätt sätt de bilder av cellerna visas i brightfield är mycket distinkta, med omkrets av cellens organ mörk mot en vit / grå cellkroppen (Figur 1a). I fluorescens immobiliserade med hjälp av celler, kan vi se tydliga fläckar intensitet, på typiskt 250-300 nm i bredd (Figur 1b). Friska, uppbundna celler kommer att synas att rotera runt den punkt tjuder bilaga i brightfield bilder. Enligt fluorescens excitation några molekylära komplex i vårt fall kan också ses vid fästpunkten, vilket indikerar en lokalisering av märkta proteinet med flagellar motorn. Dessa fläckar är enskilda molekylära komplex och antalet sett dem beror på den använda belysningen läget och hur många av komplex faktiskt finns i cellen vid en viss tidpunkt. Rörligheten hos fläckarna beror på den specifika biologiska systemet som studeras. Om tätheten av fläckar är initialt mycket högt, vilket är fallet med den märkta cytokrom används här, sedan utföra en första FRAP blekmedel kan förbättra avbildning kontrast.

Figur 1. (A) brightfield och (B) TIRF bild (falskt färg) till en orörlig Escherichia coli celler som uttrycker ett protein smält till grönt fluorescerande protein (GFP), som är känt för att vara inblandade i flagellar motorer av bakterier. Vänligen klicka här för att se en större version av figur 1.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Man måste vara försiktig att inte "över shear" cell för att titta på bundna bakterier, eftersom detta kan försämra funktionaliteten hos flagellar motorer. Det är viktigt att använda celler för mycket längre än en timme en gång på objektglas eftersom de kan bli syrefattigt. Betydande optimering kan krävas för att hitta den bästa mikroskopet avbildning villkor tillgodoses dina specifika biologiska systemet under utredning. Det kan vara klokt att försöka sig på avbildning med renat GFP ensam att fastställa den korrekta excitation intensitet laser som krävs för just ditt mikroskop system.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
Vi erkänner den typ donationer av bakteriestammar från grupper av Prof. Judith Armitage (University of Oxford, Storbritannien) och professor Conrad Mullineaux (Queen Mary University of London, Storbritannien). IMD finansieras gemensamt av Institutionen för biokemi (Oxford University) och OCISB, AR finansieras av en Engineering and Physical Sciences Research Council (EPSRC) DTC utbildningsbidrag, ND finansieras från bioteknik och Biological Sciences Research Council (BBSRC), MCL är finansieras av en Royal Society University Research Fellowship.
References
- Leake, M. C., Chandler, J. H., Wadhams, G. H., Bai, F., Berry, R. M., Armitage, J. P. Stoichiometry and turnover in single, functioning membrane protein complexes. Nature. 443, 355-358 (2006).
- Leake, M. C., Greene, N. P., Godun, R. M., Granjon, T., Buchanan, G., Chen, S., Berry, R. M., Palmer, T., Berks, B. C. Variable stoichiometry of the TatA component of the twin-arginine protein transport system observed by in vivo single-molecule imaging. Proc Natl Acad Sci U S A. 105, 15376-15381 (2008).
- Leake, M. C., Wilson, D., Gautel, M., Simmons, R. M., M, R. The elasticity of single titin molecules using a two-bead optical tweezers assay. Biophys. J. 87, 1112-1135 (2004).
- Leake, M. C., Wilson, D., Bullard, B., Simmons, R. M. The elasticity of single kettin molecules using a two-bead laser-tweezers assay. FEBS Lett. , 535-555 (2003).
- Lenn, T., Leake, M. C., Mullineaux, C. W. In vivo clustering and dynamics of cytochrome bd complexes in the Escherichia coli plasma membrane. Mol. Microbiol. 70, 1397-1407 (2008).
- Lenn, T., Leake, M. C., Mullineaux, C. W. Are Escherichia coli OXPHOS complexes concentrated in specialised zones within the plasma membrane. Biochem. Soc. Trans. 36, 1032-1036 (2008).
- Lo, C. J., Leake, M. C., Pilizota, T., Berry, R. M. Single-cell measurements of Membrane Potential, Sodium-Motive Force and Flagellar Motor Speed in Escherichia coli. Biophys. 93, 294-302 (2007).
- Lo, C. J., Leake, M. C., Berry, R. M. Fluorescence measurement of intracellular sodium concentration in single Escherichia coli cells. Biophys. J. 90, 357-3565 (2006).

{kind=link}