

Summary
여기 기능성 분자 단지가 추적 및 계량, 감지 있도록 박테리아 세포를 생활에 단일 분자 형광 현미경을 수행하는 프로토콜을 보여줍니다.
Abstract
살아있는 세포의 메커니즘에 전체 통찰력은 세포 수준에서 사건을 이끌어하고 직접 핵심 프로세스를 조사하여 얻을 수 있습니다. 생물 학적 시스템의 전단 복잡 날짜하려면 대신 상대적으로 원유 대량 앙상블 평균 측정을 사용하여 단일 시스템 연구에 초점을 맞추고, 너무 요구하는 정확한 단일 분자 실험가 발생했습니다. 그러나 많은 중요한 과정은 단지 하나의 수준에서 살아있는 세포 또는 몇 분자에서 발생, 앙상블 측정은 일반적으로이 사건의 확률과 이기종 자연을 마스크. 여기, 고급 광학 현미경 및 분석 영상 분석 도구를 사용하여 우리는 하나의 살아있는 세균 단일 분자의 정밀도로 휴대 방법과 우리가 생물 학적 기계 작동에 분자 단지 내의 역학 관계를 볼 수 있습니다 내에서 단백질을 모니터링하는 방법을 보여줍니다. 기술은 생리학 직접 관련이있다. 그들은 최소한 - 섭동의 공부에서 생물 학적 샘플을 비침습이며, 완전히 살아있는 재료에 수사 숙련 있으며, 생물 물리학의 다른 단일 분자 접근법에 즉시 사용할 수 없습니다 있습니다. 또한, 생물 학적 표본은 모든으로 자연스럽게 발생하는 것보다 훨씬 더 많은 단백질을 생성하기위한보다 일반적인지만 그다지 좋지 접근 방식에 반대하는 수정되지 않은 세포 변종 ( "게놈 인코딩") 거의 동일 수준의 찬란 - 태그 단백질 생산을 공부 ( '플라스미드 표현'). 따라서, 조사하게 될 실제 생물 학적 샘플이 자연의 생물에 매우 가까이 있으며, 실제 생리 프로세스에 따라서 관측 관련성이.
Protocol
- 이 절차를 시작하려면, 대장균에게 박테리아 세포를 표현 형광 단백질의 냉동 주식의 50 μl가 처음 사인을 알아낼 수 없을 아침에 C. 37도에서 하룻밤 다섯 ML의 LB 성장 매체 잡고 aerobically 재배,이 포화 문화의 50 μl가 추출됩니다 , 4 일에서 6 시간 동안 30도 각도의 C에서 최소 잠복기 M63 포도당 문화 미디어로 하위 교양. 여기서 우리는 전자 - 수송을 표현 한 중 서로 다른 두 가지 세포 변종을 사용하여 보여주 GFP, GFP에 융합 박테리아 flagellar 모터에 관련된 단백질을 표현하는 다른 융합 시토크롬.
- 세포 고정화 샘플로를 보는 경우도 증가하고 하위 문화에서 직접 수확하거나 그들이 "곁에"조건 하에서 볼 경우 박테리아 flagella을 잘라야 빠졌어 수 있습니다.
- 전단은 멸균 배관으로 가입이 무균 주사기로 구성된 장치로 하위 문화의 일반적으로 1 - 5ml 배치 포함됩니다. 전단은 좁은 튜브를 통해 문화를 밀어 각 주사기 펌프에서 추진하고 교류에 의해 이루어집니다. 이것은 필요한 전단의 범위에 따라 50 ~ 100 배 한 것입니다. 문화 다음 flagella 조각을 제거하는 최소한의 미디어 resuspended입니다 펠릿 세포에 centrifuged 있습니다.
- 우리는 그때 그때 드 이온화 물과 에탄올에 철저 rinsing하고 최소한 1 H. 위해 공중에 건조하기 위해 떠나는 20 분 에탄올에 코의 포화 용액에 immersing로 BK7 유리 coverslips 청소 준비
- 우리는 현미경으로 세포를 집에 간단한 흐름 셀을 구축. 이것은 BK7 유리 현미경 슬라이드에 파라핀 그리스의 라인을 그리기 다음 터널 - 샌드위치를 만들어 상단에있는 청소 coverlips 중 하나를 배치하고, 포셉 한 쌍의로 부드럽게 아래로 눌러, 5-10 μl의 흐름 세포 볼륨을주는 관련 .
- 고정화 세포를 관찰하는 우리는 폴리 - L - 라이신의 0.1 % W / V 솔루션으로 주입하여 흐름 세포를 기입하고, 그것이 최소한 1 분 상온에서 품어 수 있습니다. 우리는 다음의 흐름 세포의 한쪽 끝을에서 미디어를 주입하고 다른 조직에서 종이로 위킹 100 μl 최소한의 미디어를 사용하여 밖으로 몰아내.
- 우리는 다음 coverslip 표면을 표시하는 최소한의 미디어에 200 nm의 직경 라텍스 microspheres (Polysciences)의 1:500 희석 20 μl를 통해 심지. 흐름 세포는 coverslip은 간단한 습도 챔버의 표면의 맑은 플랫폼에 배치 아래쪽에 직면해있다 이러한 거꾸로, 5 분 실온에서 incubated입니다. 언바운드 비즈 그런 다음 100 μl 최소한의 매체를 통해서도 위킹 씻겨 있습니다.
- 우리가 곁에 세포를 관찰하고자하는 경우에는 우리는 폴리 - L - 라이신 단계를 생략하고 대신 5 MG / ML 방지 flagellin 항체와 흐름 - 세포를 작성하십시오. 흐름 세포는 10 분 습도 챔버에서 개최하고 다음 위킹로를 통해 플러시됩니다.
- 세포 배양의 20 μl가 고정화 세포를 보는에 닿는 세포 또는 unsheared 샘플을 관찰하는 경우도 가지각색 샘플을 사용하여 흐름 세포를 통해 다음 사악한입니다.
- 흐름 셀은 뒤집혀 20 분 습도 챔버에 배치됩니다. 언바운드 세포 그런 다음 100 μl 최소 매체를 통해서도 위킹에 의해 씻겨 있습니다.
- 침지 기름 방울이 coverslip의 상단 표면의 중앙에 위치하며 흐름 세포 그 다음 높은 숫자 조리개 목적 렌즈와 광학 접촉을 만드는 사용자 정의 지어진 형광 현미경의 샘플 홀더에 가볍게 배치됩니다.
- 현미경 전자 - 곱한 카메라가 켜져 있고 카메라가 -70도 C로 냉각되도록 설정이며, 소프트웨어는 처음에 게인 컨트롤을 disenabling, 프레임 전송 모드에서 25 Hz의 전형적인 프레임 속도로 이미지를 얻기위한 설정 카메라.
- 브라이트 조명이 켜져 있으며, 이미지가 자신이 coverslip 표면 자신의 긴 축 병렬로 붙어되는 바탕으로 몇 군데로 세포의 적절한 셀 또는 그룹을 선택 초점으로 가지고있다. 초점은 잘게 coverslip 표면에 붙어있는 200 나노미터 라텍스 비즈 그냥 초점되는 것을 보장하기 위해 조정됩니다.
- 이미지 시퀀스는 셀 본문의 개요를 기록 브라이트 인수입니다. 브라이트 조명이 꺼져되고 카메라 게인을 최대로 사용할 수 있습니다.
- 총 내부 반사 형광 (또는 TIRF) 적절한 파장을 통해 레이저 광선을 (여기서, 녹색 형광 단백질 여기에 대해, 우리는 473 nm의 레이저를 사용)를 사용하여 표준 취득의 뒤쪽 초점 평면에 집중되는만큼 사전 설정 목적 렌즈지만 세포 샘플에 형광 여기에 대한 지극히 미미한 필드를 생성하는 광학 축에서 옆으로 있질.
- 카메라 인수는 시작 레이저 셔터가 세균 내에 형광 단백질들을 흥분시키기 위해 열립니다. 인수의 레이저 강도와 속도에 대한 매개 변수는 다른 값으로 실험에 의해 조사를 받고 특정 생물 학적 시스템을 최적화할 필요가 있지만, 일반적인 범위 R세포막의 모바일 단백질 단지를 공부하는 elevant는 50-40 MS의 프레임 당 노출 시간, 직경 ~ 30 μm의의 원형 여기 영역 1-10 MW 레이저 파워입니다. 샘플은 photobleached, 일반적으로에 대한 ~ 10 S.까지 조명 아르
- 이 프로토콜은 두 세포의 시체가 coverslip과 flagellar 스텁 사이의 첨부 파일의 지점에 대한 회전되는 닿는 세포, 그리고 rigidly coverslip 표면에 고정되어 있습니다 고정화 세포를 사용할 수 있습니다.
- 단지 높은 사본 번호가 일부 세포 변종 이전 TIRF 영상에 셀 중 하나를 장대에 초기 회절 - 제한된 초점을 맞춘 레이저 표백제의 혜택을 누릴 수 있습니다. 이 표백제 (또는 FRAP) photobleaching 후 형광 복구에 사용하는 것과 동일합니다. 우리의 사용자 정의 현미경 사용 여기 레이저 광 중 일부는 FRAP 타입 표백에 사용되는 두 번째 독립적인 경로로 내려 먹이 수 있습니다. 일반적으로 1 - 10mW 레이저 전원 범위 10-300 MS의 전형적인 표백 시간과 함께 사용됩니다. 이후보다 쉽게 시각으로 그 지역에 확산 개별 단지를 허용 세포의 표백 영역에서보다 높은 영상 반면이 발생합니다.
- "slimfield"라는 다른 조명 모드를 고용하는 세포막에서보다 훨씬 빠른 확산 세포 세포질에서 단지를 시각화하십시오. 여기 초점 레이저가 옆으로 단지 하나의 세포를 포괄하기 위해 확장됩니다. 이것은 일반적으로 밀리초의 속도가 훨씬 빠르고 노출을 취할 수있게 아주 강렬한 필드를 생성합니다.
- 데이터 수집 후, 이미지가 사용자 정의 작성된 소프트웨어 (LabVIEW 8.5에서 코드)로 공급하고 있습니다. 이것은 자동으로 일반적으로 몇 나노미터의 정밀도로 세포에 형광 명소의 위치를 감지하고 그들의 크기와 밝기를 추출합니다. 추적 분자 복잡한 시간과 관련하여 photobleaching 추적은 다음 stoichiometry을 추정하는 데 사용되는 brightnessof은 단일 분자 복잡한을 만들어 많은 개인 형광 단백질 즉.
대표 결과 :
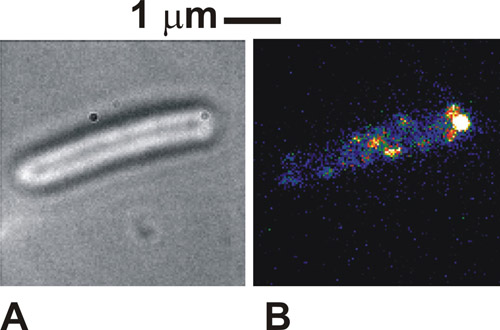
프로토콜이 완료되면 브라이트에서 볼 세포의 정확하게 이미지는 흰색 / 회색 세포 기관 (그림 1A)에 대한 어두운 세포 기관의 경계와 함께 매우 독특한 것입니다. 형광 사용하여 고정화 세포에서, 우리는 폭 일반적으로 250-300 nm의 (그림 1B)의 독특한 명소 강도를 볼 수 있습니다. 건강 닿는 곳에 전지 브라이트 이미지의 테더 부착 지점 주위에 회전 볼 수 있습니다. 형광 여기에서 우리 사건의 일부 분자 단지도 flagellar 모터와 태그 단백질의 지방화를 나타내는, 첨부 파일의 시점에서 볼 수 있습니다. 이 반점은 개별 분자 단지이며, 본 그들의 숫자는 조명 사용 모드와 방법에 단지 많은 실제로 한 번에 세포에 존재에 따라 달라집니다. 반점의 이동성 연구에서 특정 생물 학적 시스템에 따라 다릅니다. 관광 명소의 밀도가 처음 매우 높다면, 그 다음 초기 FRAP의 표백제가 이미징 명암을 향상시킬 수 수행, 여기를 사용한 표시된 cytochromes 경우가 있습니다.

그림 1. (A) 브라이트와 고정화 대장균 세포에 대한 (B) TIRF 이미지 (거짓 색상)은 박테리아의 flagellar 모터에 관련된 것으로 알려진 녹색 형광 단백질 (GFP)을 융합 단백질을 표현. 하시기 바랍니다 여기를 클릭 그림 1의 더 큰 버전을 볼 수 있습니다.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
케어는이 이후 닿는 박테리아보고에 대해 "이상의 전단"세포가 flagellar 모터의 기능을 손상 수하지 않도록해야합니다. 그들은 산소가 고갈 될 수 있으므로 현미경 슬라이드에 한 시간보다 훨씬 오래 세포를 사용하는 것이 중요합니다. 상당한 최적화가 가장 현미경 이미징 조건 조사중인 특정 생물 학적 시스템에 음식을 제공 했었죠 찾을 수 필요할 수 있습니다. 그것은 특정 현미경 시스템에 필요한 정확한 강도 레이저 여기를 확인 혼자 정화 GFP를 사용하여 영상을 시도하는 것이 현명 수 있습니다.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
우리는 교수 유디트 아미티지 (옥스포드 대학, 영국)와 교수 콘라드 뮬리노 (런던 퀸 메리 대학, 영국)의 그룹에서 세균성 긴장의 종류의 기부금을 인정합니다. IMD는 공동 생화학 뎁트 오브 (옥스포드 대학)과 OCISB에 의해 후원되며 AR는 공학 및 물리 과학 연구위원회 (EPSRC) DTC의 학생이기 의해 후원되며 ND는 생물 공학 및 생물 과학 연구 협의회 (BBSRC)에서 재정 지원되며 MCL입니다 로열 사회 대학의 연구 활동에 의해 자금.
References
- Leake, M. C., Chandler, J. H., Wadhams, G. H., Bai, F., Berry, R. M., Armitage, J. P. Stoichiometry and turnover in single, functioning membrane protein complexes. Nature. 443, 355-358 (2006).
- Leake, M. C., Greene, N. P., Godun, R. M., Granjon, T., Buchanan, G., Chen, S., Berry, R. M., Palmer, T., Berks, B. C. Variable stoichiometry of the TatA component of the twin-arginine protein transport system observed by in vivo single-molecule imaging. Proc Natl Acad Sci U S A. 105, 15376-15381 (2008).
- Leake, M. C., Wilson, D., Gautel, M., Simmons, R. M., M, R. The elasticity of single titin molecules using a two-bead optical tweezers assay. Biophys. J. 87, 1112-1135 (2004).
- Leake, M. C., Wilson, D., Bullard, B., Simmons, R. M. The elasticity of single kettin molecules using a two-bead laser-tweezers assay. FEBS Lett. , 535-555 (2003).
- Lenn, T., Leake, M. C., Mullineaux, C. W. In vivo clustering and dynamics of cytochrome bd complexes in the Escherichia coli plasma membrane. Mol. Microbiol. 70, 1397-1407 (2008).
- Lenn, T., Leake, M. C., Mullineaux, C. W. Are Escherichia coli OXPHOS complexes concentrated in specialised zones within the plasma membrane. Biochem. Soc. Trans. 36, 1032-1036 (2008).
- Lo, C. J., Leake, M. C., Pilizota, T., Berry, R. M. Single-cell measurements of Membrane Potential, Sodium-Motive Force and Flagellar Motor Speed in Escherichia coli. Biophys. 93, 294-302 (2007).
- Lo, C. J., Leake, M. C., Berry, R. M. Fluorescence measurement of intracellular sodium concentration in single Escherichia coli cells. Biophys. J. 90, 357-3565 (2006).

{kind=link}