

Summary
我们提出了一种方法, 以量化淀粉含量的卵巢原基在冬季休眠期间使用图像分析系统结合组织化学技术。
Abstract
在几个植物发育过程中, 淀粉的变化与关键事件有关, 包括从授粉到受精的生殖阶段和果实的开始。然而, 淀粉在花分化过程中的变化尚不完全清楚, 主要是由于难以量化淀粉含量的特别小的结构的花原体。在这里, 我们描述了一种方法, 以定量淀粉在卵巢原基甜樱桃 (金银花) 使用图像分析系统附加到显微镜, 允许将淀粉含量的变化与不同阶段的关系从秋天到春天的休眠状态。为此, 通过评价在冬季不同时刻转移到可控条件下的芽芽生长, 确定花蕾的休眠状态。为了定量卵巢原基中的淀粉, 花蕾按顺序收集、固定、嵌入石蜡、切片, 并用 i2kl (碘化钾) 染色。在显微镜下观察制剂, 并通过图像分析仪进行分析, 该分析仪可将淀粉与背景明确区分开来。淀粉含量值是通过测量与染色淀粉相对应的图像的光学密度得到的, 同时考虑到每个像素的光学密度之和作为对所研究框架淀粉含量的估计。
Introduction
温带多年生植物通过调节生长发育来适应季节。虽然它们在春季和夏季发育, 但在秋天停止生长, 在冬季处于休眠状态。虽然休眠使他们能够在较低的冬季温度下生存, 寒意是一个先决条件, 在春天适当的芽2。休眠对温带水果生产和林业的重要影响, 导致了确定和预测休眠期的各种努力。在果树物种中, 将芽转移到强迫条件下的经验实验和基于开花数据的统计预测是目前确定休眠打破日期的方法, 这使得研究人员能够估计每个品种的冷却要求。然而, 如何根据生物过程确定休眠状态仍不清楚3。
在温带果树上开花, 如甜樱桃, 每年开花一次 , 持续约两周。然而, 花开始分化和发展大约10个月前, 在前一个夏天4。花原基在秋天停止生长, 在冬季在芽内保持休眠状态。在此期间, 每个品种需要积累适当开花的特定冷却要求 4.尽管在冬季芽缺乏物候变化, 但在休眠期间, 花原体在生理上是活跃的, 最近, 寒冷温度的积累与淀粉积累或减少的动态有关在卵巢原基细胞内, 为休眠测定提供了一种新的方法 5.然而, 卵巢原基的小尺寸和位置需要一种特殊的方法。
淀粉是木本植物中主要的碳水化合物.因此, 淀粉的变化与花组织的生理活性有关, 花组织需要碳水化合物来支持它们的发育7,8。不同的关键事件在生殖过程中也与不同花系中淀粉含量的变化有关, 如花药减数分裂9、花粉管的生长通过花粉室受精 10.组织化学技术可以在休眠期检测花原基每个特定组织中的淀粉。然而, 在量化淀粉, 使其随着时间的推移减少的模式或比较组织, 品种或年份之间的淀粉含量仍然是困难的。这是因为可用于分析技术的组织数量很少11。作为一种替代方法, 与显微镜12 相关联的图像分析允许对植物组织13的非常小的样本中的淀粉进行定量。
显微镜和图像分析相结合的方法已被用来量化植物组织中不同成分的含量, 如钙质14、微管15或淀粉16, 方法是测量按特定染色面积的大小。污渍。对于淀粉, 它可以很容易地检测到它使用碘钾 (i2ki) 反应17。此方法是非常具体的;i2 ki 在淀粉颗粒的层流结构内进行夹层, 并根据淀粉18的直链淀粉含量形成深蓝色或红棕色。染色 i2ki 染色的切片显示淀粉和背景组织之间的充分对比度, 允许明确的淀粉检测和随后的量化图像分析系统19。虽然这种染料不是化学计量的, 但碘的积累与淀粉分子的长度成正比, 淀粉分子的长度变化很大,变化很大。因此, 以像像像像素数表示的染色面积的大小可能无法准确地反映淀粉的含量, 因为在染色面积相似的字段之间可以发现淀粉含量的高差异。作为一种替代方法, 可以通过测量从显微镜获得的黑白图像上的染色颗粒的光学密度来评估淀粉含量, 因为在杏子8,13的不同组织中都有报道。,19, 鳄梨10,20和橄榄21.
本文介绍了一种将休眠状态的实验测定与秋季至春季甜樱桃卵巢原基组织淀粉含量的定量相结合的方法, 为理解和预测提供了一种新的工具。休眠的基础上研究的生物机制与休眠。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 休眠测定和植物材料收集
- 在田野里的花蕾取样。休眠研究是长期的实验, 需要成年树足够大, 可以在整个冬天收集芽和芽, 而不会影响树木在明年春天的发育。根据培训系统, 可能需要特别的果园管理;因此, 修剪可能比水果生产的严重程度要小。
- 每周, 从初秋到芽断裂开始, 收集和称重 1 0个花蕾。
- 将花蕾放入带瓶盖的10毫升玻璃管中, 并在4°c 下将样品浸泡在乙醇/乙酸 (3:1) 的固定溶液中至少24小时。然后, 丢弃固性剂, 加入75% 的乙醇, 慷慨地确保它覆盖样品。样品可在4°C 下保存在该溶液中, 直至使用。
注: 真空渗透可用于去除芽内的气泡, 防止其漂浮。这有利于固定物进入组织, 但可能会损害组织的结构。如果没有必要, 尽量避免这样做。
- 在田野里的芽取样。
- 每周, 从初秋到芽断裂开始, 取三枝, 长15-30 厘米, 直径5毫米, 每个花蕾至少有10个花蕾。将它们放在水浸的花店泡沫上, 放在22±1°c 的生长室中, 并有12小时的光周期。
- 在生长室呆了10天后, 从芽中摘下并称重10个花蕾。
- 评估芽的生长, 并确定休眠状态。取样周期必须根据地点的条件进行调整。在果园条件下 (西班牙萨拉戈萨, 北纬 41°44 ' 30 ", 西经零期 47 ' 00", 海拔220米), 从11月30日至2月底或3月初对芽进行了取样。
- 每周通过比较从田间采摘的10个花蕾的重量, 评估花蕾对室内适宜生长条件的反应, 从初秋到春季的芽爆裂。
- 如果没有差异或这些差异小于 30%, 请考虑芽没有满足其冷却要求, 是内生 22.如果差异超过 30%, 请考虑芽已满足其冷却要求, 并且是生态激素22。
2. 淀粉定量的植物材料制备
- 从每个采样日期选择大约六个固定芽 (请参阅步骤 1.1)。根据不同的品种, 每个甜樱桃花芽包含多达五个花原基。
- 取出外部芽鳞片, 用75% 乙醇将芽放在制表师的玻璃上, 以防止其干燥。
- 在立体显微镜下, 用精确的钳子和眼药部手术刀对花蕾进行解剖, 每芽提取至少一个花原基。花原基可以保存在一个10-mL 玻璃管与75% 乙醇在4°c 下, 或立即进行下一步。
- 在三丁醇系列中脱水样品。
- 将75% 的乙醇溶液替换为10% 毫升的75% 的三丁醇 (TBA), 以慷慨地覆盖样品, 并将其孵育1.5 小时。巴斯德移液器可用于丢弃溶液。
- 重复步骤 2.2.1, 在30°c 的干燥炉中加入浓度上升的 TBA (85%、95% 和 100%, v2; 和3x 纯 TBA), 并在空气萃取的情况下结晶, 因为纯 TBA 在室温 (< 20°c) 下结晶, 并且非常易挥发和有毒。如果 TBA 与样品结晶, 就会损害组织。
- 将样品嵌入石蜡18中.
- 在60°c 的干燥炉中引入石蜡, 并在前一天提取空气, 将石蜡熔化。石蜡珍珠也可以在热板上融化, 但要确保液体石蜡在 6 0°c, 而不是在较高的温度下, 以避免组织损伤。
- 将 TBA 替换为 TBA 和石蜡油 (1: 1) 的混合物, 并在60°c 的烘干炉内孵育24小时。然后, 将 TBA 和石蜡油的混合物换成纯熔融石蜡, 在60°c 的干燥炉内孵育至少6小时。重复 2倍, 孵育最后一个变化至少 4-6 d。
- 将每个样品放置在热表面的小金属基模上, 嵌入石蜡中, 并调整嵌入盒式磁带的速度。将其放在冷的表面上, 并在蜡凝固后取出块。
- 对制剂进行分割和补水。
- 制备 Hauppt 的粘合剂: 在30°c 的蒸馏水中溶解1克普通诺克斯明胶;然后加入2克苯酚松散晶体 (c6h5oh) 和15毫升甘油。用刷子将 Hauppt 的粘合剂涂在玻璃滑梯上, 并加入1% 的甲醛溶液。
- 将每个石蜡块在旋转微料中的10微米处进行分割, 并将部分放置在用 Haupt 胶粘剂覆盖的玻璃滑梯上。
- 将玻璃滑块与切片放在35-40°c 的热表面上, 直至干燥;然后, 将玻璃幻灯片移动到幻灯片架。
- 准备脱蜡和补液溶液。将200毫升的组织化 II 放入三个玻璃染色盘中, 另一个与组织清晰 II: 乙醇 (1: 1, v2)、乙醇系列 (100%、70%、40%、v2) 和最终蒸馏水清洗。
- 在组织清洗 ii 和组织清楚 ii 中, 用三个洗涤, 每个 5分钟, 去倒部分: 乙醇 (1: 1, v/v)。将滑片架放在玻璃染色盘内, 确保溶液完全覆盖幻灯片, 然后将滑片架从一个解决方案移动到下一个解决方案。
- 通过以下2分钟洗涤系列对切片进行补水: 乙醇系列 (100%、70%、40%、v) 和最终蒸馏水清洗。最后, 在室温下擦干玻璃滑梯。
- 污渍部分 18。
- 准备 i2ki 染色: 在100毫升蒸馏水中溶解2克碘化钾 (ki) 和0.2 克碘 (i2)。在部分上涂一滴新鲜的 i2ki 5分钟, 然后用吸墨纸吸收, 去除多余的污渍。快速继续下一步。
- 涂上一小滴合成安装介质, 在上面放置一个小盖板玻璃, 用力按压。一旦安装介质干燥, 在明亮的场显微镜下观察卵巢淀粉的初步评估。
注意: 此步骤不是强制性的, 虽然获得了更好的对比淀粉和背景之间。如果淀粉定量后必须与其他污渍重复使用, 请将盖板玻璃放在污渍的滴上, 用吸墨纸吸收多余的污渍 (见 2.5.1)。然后, 在淀粉定量后, 用蒸馏水冲洗出 i2ki 染色, 并将制剂放在热表面的热表面上, 直到干燥。
3. 淀粉含量的量化
- 校准光学条件。
注: 图像分析仪用于检测染色淀粉的检测水平直接取决于光线条件和显微镜的放大倍率;因此, 为所有评估的制剂确定这些条件。根据可用的显微镜和光线条件调整此处建议的调整。- 在20X 放大倍率下调整光圈膜片, 并控制亮度或亮度。
- 确保滤芯上没有过滤器, 并在显微镜中选择明亮的字段条件。选择合适的放大倍率 (例如, 甜樱桃卵巢原基的 40x)。
- 控制图像采集条件。通过图像调整相机设置, 在没有组织的情况下进行染色准备收购业务预查看。
- 将亮度固定在 50%, 增益固定在 1.0 x, 直方图将伽玛值确定为 1.00, 对比度为 0-100, 与亮度分布直方图的限制排列在一起。
- 激活过曝光/曝光不足功能, 并在过度曝光的限度上调整曝光时间。
- 将白平衡函数应用于完整图像, 以显示图像中所有中性颜色的组件, 而无需任何色调和对完整图像的底纹校正, 以创建校正和均匀的图像。
- 校准图像分析系统, 以获得通过透射率 (T) 值获得的不同光学密度 (od) 的灰度 (0, 黑色; 255, 白色) 的控制值。
- 获取无组织染色制剂的图像, 考虑控制白色, 并通过测量测量黑白图像的灰度灰色度量校准灰色参考值 = 0测量值校准好吧好吧好吧好吧这对应于100% 的透光率;因此, 光学密度为 0, 根据 OD = 2-log T。
- 使用4N 滤镜获取相同制备的图像, 从而减少光的 4倍, 并通过测量测量黑白图像的灰度灰色度量校准灰色参考值 = 0。6测量值校准好吧好吧好吧好吧这对应于25% 的透光率;因此, 光学密度 0.6, 根据 OD = 2-log T。
- 获取不带光的相同制备的图像, 被视为黑色, 并通过测量测量黑白图像的灰度灰色度量校准灰色参考值 = 1测量值校准好吧好吧好吧好吧这对应于0% 的透光率;因此, 光学密度为 1, 根据 OD = 2-log T。
- 检测淀粉。
- 获取字段的彩色图像, 以 TIFF 格式测量, 分辨率至少为每英寸 300点 (dpi)。
- 创建与染色区域相对应的二进制图像。设置三个颜色阈值 (每个值在0-255 之间), 直到二进制图像准确地反映了通过图像观察到的染色淀粉颗粒检测选择红色、蓝色和绿色的阈值好吧好吧好吧好吧在各种制剂和组织中反复进行视觉比较, 以调整最终的检测水平。存储和使用这些级别的所有准备工作。
- 对淀粉进行定量。使用图像分析系统将原始彩色图像转换为黑白图像。使用二进制图像作为叠加掩码上的黑白图像通过图像二进制编辑。通过测量测量掩码下每个像素的光学密度之和 灰色级别可以,并考虑这个值作为淀粉含量在测量领域。
- 重复步骤 3.4.1-3.4.3 在四个地方的卵巢原基, 以获得一个代表性的价值的淀粉含量在卵巢的原始体。
- 重复步骤 3.4.1-3.4.3 和步骤3.4.5 在不同的花的每个收集日期。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
休眠研究需要确定满足冷却要求的时刻。尽管在野外条件下冬季缺乏物候变化 (图 1a), 但樱桃树在低温下度过一定时期之前, 在适当的条件下无法恢复生长能力。在冬季, 芽定期转移到受控条件室 (图 1b), 从而可以评估花蕾的休眠状态。通过测定花蕾重量的增加, 对花蕾生长进行了评价。虽然在休眠期间, 在10天的适当条件下无法观察到任何变化 (图 1c), 但一旦克服休眠, 生长室的芽就会膨胀和爆裂 (图 1C)。这一分析的结果使芽的休眠状态得以确定。由于冬季发生的温度不同, 根据年份的不同, 在不同的日期克服了休眠问题。而在研究的第一年, 休眠的打破发生在 1月, 第二年则是一个温和的冬天;因此, 令人寒心的满足发生在大约三个星期后, 在2月。
甜樱桃在刺中带有花蕾, 其中顶芽是植物芽, 侧芽是花蕾 (图 1C和1C)。在夏末开始分化为花蕾或植物芽, 当它们在冬季进入休眠状态时, 几个花原体仍留在芽内, 由无数的绿色鳞片保护, 并被棕色的外鳞片覆盖 (图 1c和 图 2a)。花蕾的解剖显示了花芽内的小花原基 (图 2a)。每个花蕾含有一到五个单独的花原体 (图 2B)。尽管每个花原基的小尺寸, 花的所有部分都是区分的, 可以区分: 雌蕊, 花药, 花瓣, 和隔片 (图 2c)。使用组织化学技术 (酒精: 醋酸 [3: 1] 固定、石蜡包埋、微分片和碘基淀粉染色) 可以观察到淀粉在花原基组织中的分布 (图 2d)).
卵巢原基中的淀粉在每个部分进行了量化。四种测量指标为 1337μm 2, 放大 40x, 代表了甜樱桃卵巢原基中淀粉的总体布局 (图 3 a).在 i2ki 染色 (图 3B) 后, 淀粉颗粒与背景明显区分开来。图像分析系统通过调整红色、绿色和蓝色的颜色阈值来识别淀粉, 直到所观察到的所有淀粉颗粒都被系统根据定义的颜色参数 (图 3C) 创建的二值图像覆盖。所获得的淀粉含量值是测量黑白图像上遮罩下每个像素的光学密度的结果 (图 3d)。
淀粉的定量显示了淀粉在冬季动态的一致模式 (图 4)。通常情况下, 初冬淀粉的数量的光学密度值不到 40, 0003, 而最大数量在这两年的价值达到 120, 000 至 140, 000 之间。虽然第一年1月份达到了最大值 (图 4 a), 但第二年2月达到了最大值 (图 4 b)。将这些结果与休眠肤色相比较, 淀粉的最大含量与这两年的冷却效果同时发生。
这种方法要求确定休眠状态 (图 5a) 与卵巢组织上的淀粉定量 (图 5A), 以确定淀粉含量的变化与休眠。

图 1: 测定甜樱桃花蕾休眠状态的实验装置.(a) 在冬季, 树枝显示休眠的芽关闭, 被深棕色鳞片覆盖。(b) 这个面板显示转移到生长室的芽。1月中旬, 一些品种仍然处于休眠状态, 芽仍然关闭 (白色箭头), 而另一些品种则能够生长, 显示出膨胀的芽 (黑色箭头)。(c) 这个面板显示了一个拍摄与休眠花蕾位于侧向和一个单一的植物芽位于顶端位置的刺。(d) 这个面板显示了一个拍摄的细节, 一旦在生长室的休眠克服了 10天, 显示芽肿胀。请点击这里查看此图的较大版本.
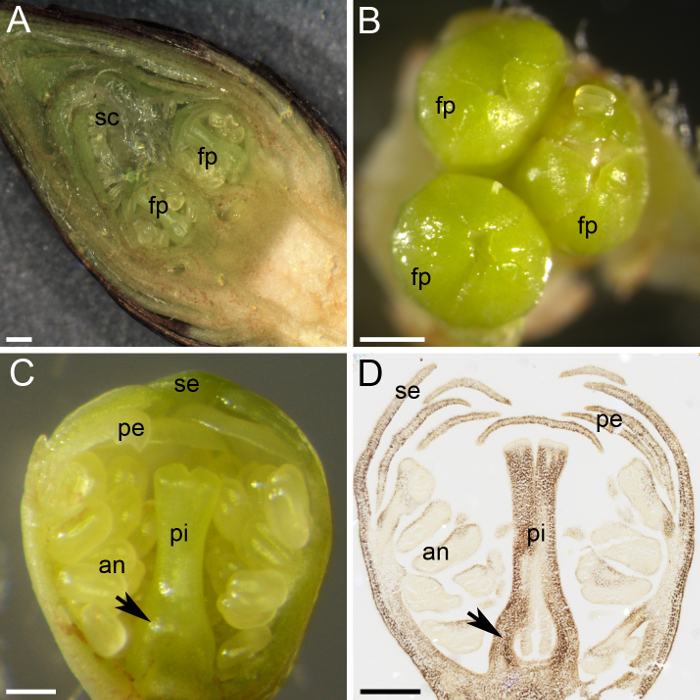
图 2: 淀粉定量的植物材料制备.(a) 这个面板显示花芽的横向部分, 显示两个花原生 (fp) 由许多鳞片 (sc) 保护。(b) 三朵花原体 (fp) 聚集在花蕾中。(c) 这个面板显示了花原基的横向部分, 所有的轮回都有区别: 隔片 (片)、花瓣 (pe)、花药 (a) 和雌蕊 (pi)。卵巢原基在雌蕊 (箭头) 的底部被区分。(d) 1月份收集并固定了一个花原基的中间部分, 嵌入石蜡中, 用 i 2 ki (深棕色) 染色.此面板显示卵巢原基 (箭头)。在a和b面板中, 刻度条为 500μm, 在 c 和d面板中, 刻度条为 200μm. 请点击这里查看此图的较大版本.

图 3: 甜樱桃卵巢原基中的淀粉定量.(a) 这个面板显示了一个涂有 i2ki 的卵巢原基的中间部分, 显示了测量淀粉含量的四个框架。(b) 这个面板显示了卵巢原基的细节。淀粉颗粒被染成深棕色。(c) 此面板显示一个伪彩色图像, 其中淀粉对应于不同色调的蓝色。(d) 此面板显示一个二进制图像面膜覆盖 i2ki 染色淀粉 (蓝色) 的黑白原始图像。光学密度仅在掩码所覆盖的原始图像的像素中进行测量。在 a 面板中, 刻度条为 100μm, 在 b-d 面板中, 刻度条为 20μm. 请点击这里查看此图的较大版本.

图 4: 在两年不同的冬季温度条件下, 从秋季到春季每月采集的甜樱桃卵巢原基淀粉定量的代表性结果.(a) 该小组显示了2010-2011年冬季寒冷的结果。冷却的满足 (雪花) 发生在 1月, 伴随着淀粉的最大数量。(b) 该小组显示了2011-2012年的结果, 该年是一个温和的冬季。冷却的满足 (雪花) 发生在 2月, 伴随着淀粉的最大数量。这些值是平均值±的平均值的标准误差。请点击这里查看此图的较大版本.
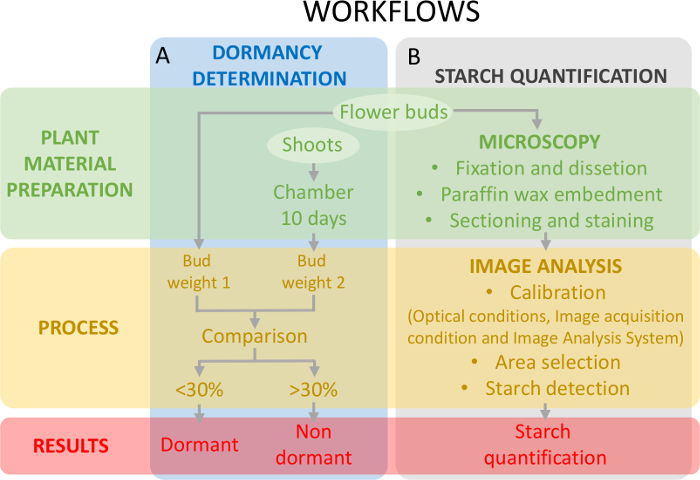
图 5: 评价甜樱桃卵巢原基芽休眠状态和淀粉定量的实验设计方案.(a) 此面板显示休眠状态确定的工作流程: 工厂材料的制备、工艺和获得的结果。(b) 该面板展示了淀粉定量的工作流程: 对淀粉进行显微观察的芽的组织化学制备、淀粉的图像分析检测和淀粉的定量。请点击这里查看此图的较大版本.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
在气候变化的情况下, 木本多年生植物的休眠对水果生产和林业产生了明显的影响, 尽管休眠背后的生物过程仍不清楚。休眠研究可以从不同的角度来探讨, 但在过去几年里, 寻找冬季休眠生物标记的研究有所加强。然而, 大多数试图找到一个明确的指标, 当一个芽已经打破休眠的失败 3.本文所描述的方法,将组织化学技术18与分析图像相结合, 对研究某一特定组织的碳水化合物储量与其在甜樱桃芽中的生理活性之间的关系非常有用在休眠5的不同阶段, 也可应用于其他物种和组织10,13,20.
淀粉形式的碳水化合物储备在花卉发育和生殖过程中发挥着重要作用,7、10、13、20、21、23和在温带木本多年生植物的季节性中.对休眠的不同研究对24 芽内的淀粉进行了研究。然而, 由于其体积小, 需要几个完整的芽使用定量分析方法, 检测定量变化的特定组织或细胞受到周围细胞的掩蔽效应的限制。组织化学技术与图像分析系统的结合, 为研究芽内不同结构淀粉含量的变化提供了很好的机会。
该方法的局限性是防止对组织中淀粉的确切含量进行量化, 但允许对相对淀粉含量进行量化, 以跟踪淀粉随时间的定量变化25并比较淀粉含量。不同的组织13,20, 品种, 或5年。为了能够正确比较场、组织和芽之间的光学密度值, 必须准确地建立系统的校准 (光线条件、染色强度和放大倍率) 和颜色阈值的设置, 存储, 并用于所有准备工作。
淀粉染色和定量的基础上, 钾碘允许随后使用其他污渍后清洗部分。因此, 可以在不增加制剂和不使用合成安装介质 5,20的情况下进行不同的分析。同样, 形态测量26可以在相同的制备后完成, 允许淀粉积累的模式相对于不同结构的生长 5,13。该方法可以适应其他结构或物种与颜色水平的调整, 使用分析仪检测淀粉, 这可以允许研究涉及淀粉含量变化的其他发展过程, 在小组细胞。
该方法公布的卵巢原基休眠释放与淀粉积累之间的关系, 为了解休眠和冷却要求的生物学基础提供了良好的基础5。然而, 通过石蜡嵌入切片的图像分析对淀粉进行定量, 对于估计大量品种的冷却需求可能会非常繁琐和耗时。今后的努力必须集中在研究可靠的生物指标上, 这些指标可以很容易地表明树木的休眠状态。同时, 休眠过程中淀粉的变化规律可用于进一步的生理和遗传研究, 本文所述的组织化学技术与图像分析相结合, 可用于其他木本植物作物。量化不同组织的淀粉含量与休眠的关系。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者没有什么可透露的。
Acknowledgments
作者感激地感谢玛丽亚·赫雷罗和埃利塞奥·里瓦斯的有益讨论和建议。这项工作得到了欧洲联盟欧洲区域发展基金----欧洲区域发展基金----的支持 [赠款编号 bes-2010-037992 至 E. f.];国家调查研究所 [赠款编号 Rfp2015-00015-00, RTA全85-00, RTA2017-00003-00];和 Gobierno de Aragón----欧洲社会基金, 欧洲联盟 [Grupo 格兰达·阿林塔多 a12-17r]。
Materials
| Name | Company | Catalog Number | Comments |
| Precision scale | Sartorius | CP225D | |
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Drying-stove | Memmert | U15 | |
| Paraffin Embedding station | Leica Microsystems | EG1140H | |
| Rotatory microtome | Reichert-Jung | 1130/Biocut | |
| Microtome blade | Feather | S35 | Stainless steel |
| Bright field microscope | Leica Microsystems | DM2500 | |
| Digital Camera | Leica Microsystems | DC-300 | |
| Image Analysis System | Leica Microsystems | Quantiment Q550 |
References
- Kurokura, T., Mimida, N., Battey, N. H., Hytönen, T. The regulation of seasonal flowering in the Rosaceae. Journal of Experimental Botany. 64 (14), 4131-4141 (2013).
- Rohde, A., Bhalerao, R. P.
- Fadón, E., Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environmental and Experimental Botany. 152, 28-36 (2018).
- Fadón, E., Rodrigo, J., Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees - Structure and Function. , In Press (2018).
- Fadón, E., Herrero, M., Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Frontiers in Plant Science. 9 (171), (2018).
- Loescher, W. H., Mccamant, T., Keller, J. D. Carbohydrate reserves, translocation and storage in woody plant roots. HortScience. 25 (3), 274-281 (1990).
- Hedhly, A., et al. Starch turnover and metabolism during flower and early embryo development. Plant Physiology. , (2016).
- Rodrigo, J., Hormaza, J. I., Herrero, M. Ovary starch reserves and flower development in apricot (Prunus armeniaca). Physiologia Plantarum. 108 (1), 35-41 (2000).
- Julian, C., Rodrigo, J., Herrero, M. Stamen development and winter dormancy in apricot (Prunus armeniaca). Annals of Botany. 108 (4), 617-625 (2011).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil starch reserves at anthesis correlate with final flower fate in avocado (Persea americana). PLoS One. 8 (10), e78467 (2013).
- Smith, A. M., Zeeman, S. C. Quantification of starch in plant tissues. Nature Protocols. 1 (3), 1342-1345 (2006).
- Eliceiri, K. W., et al.
- Rodrigo, J., Herrero, M. Influence of intraovular reserves on ovule fate in apricot (Prunus armeniaca L.). Sexual Plant Reproduction. 11, 86-93 (1998).
- Zhou, J., Spallek, T., Faulkner, C., Robatzek, S. CalloseMeasurer: A novel software solution to measure callose deposition and recognise spreading callose patterns. Plant Methods. 8 (1), (2012).
- Faulkner, C., et al. An automated quantitative image analysis tool for the identification of microtubule patterns in plants. Traffic. 18 (10), 683-693 (2017).
- Kuhn, B. F. Determination of starch in ovules of the sour cherry cv. "Stevnsbaer.". European Journal of Horticultural Science. 71 (3), 120-124 (2006).
- Johansen, D. A. Plant microtechnique. , McGraw-Hill. New York, NY. (1940).
- Ruzin, S. E. Plant microtechnique and microscopy. , Oxford University Press. New York, NY. (1999).
- Rodrigo, J., Rivas, E., Herrero, M. Starch determination in plant tissues using a computerized image analysis system. Physiologia Plantarum. 99 (1), 105-110 (1997).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Ovary starch reserves and pistil development in avocado (Persea americana). Physiologia Plantarum. 140 (4), 395-404 (2010).
- Suarez, C., Castro, A. J., Rapoport, H. F., Rodriguez-García, M. I. Morphological, histological and ultrastructural changes in the olive pistil during flowering. Sexual Plant Reproduction. 25, 133-146 (2012).
- Lang, G. A., Early, J. D., Martin, G. C., Darnell, R. L. Endodormancy, paradormancy, and ecodormancy - Physiological terminology and classification for dormancy research. HortScience. 22 (3), 371-377 (1987).
- Hedhly, A., Vogler, H., Eichenberger, C., Grossniklaus, U. Whole-mount clearing and staining of arabidopsis flower organs and siliques. Journal of Visualized Experiments. 2018 (134), 1-10 (2018).
- Kaufmann, H., Blanke, M. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. Journal of Plant Physiology. 218 (July), 1-5 (2017).
- Herrero, M., Dickinson, H. G. Pollen-pistil incompatibility in Petunia hybrida: changes in the pistil following compatible and incompatible intraspecific crosses. Journal of Cell Science. 36, 1-18 (1979).
- Carpenter, A. E., et al. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biology. 7 (10), R100 (2006).
