

Summary
Se presenta una metodología para cuantificar el contenido de almidón en los primordios del ovario en cerezo dulce (Prunus avium L.) durante la dormancia invernal mediante el uso de un sistema de análisis de imagen combinado con técnicas histoquímicas.
Abstract
Cambios en el almidón en pequeñas estructuras se asocian a eventos clave durante varios procesos de desarrollo de planta, incluyendo la fase reproductiva de polinización, fecundación y el inicio de la fructificación. Sin embargo, variaciones en almidón durante la diferenciación de flores están completamente desconocidas, principalmente debido a la dificultad de cuantificar el contenido de almidón en las estructuras particularmente pequeños de los primordios de flores. Aquí, describimos un método para la cuantificación de almidón en los primordios del ovario de la cereza dulce (Prunus avium L.) mediante el uso de un sistema de análisis de imagen atado microscopio, que permite la relación con los cambios en el contenido de almidón con las diferentes fases de latencia de otoño a primavera. Para ello, el estado de dormancia de las yemas de flor se determina evaluando el crecimiento de yemas de brotes transferidos a condiciones controladas en diferentes momentos en época de invierno. Para la cuantificación de almidón en los primordios del ovario, capullos son secuencialmente recogidos, fijo, embebidos en parafina, seccionadas y teñidas con2Kl (yodo yoduro de potasio). Las preparaciones se observa bajo el microscopio y analizadas por un analizador de imagen que distingue claramente el almidón desde el fondo. Valores de contenido de almidón se obtienen midiendo la densidad óptica de la imagen que corresponde a almidón teñido, considerando la suma de la densidad óptica de cada píxel como una estimación del contenido de almidón del marco estudiado.
Introduction
Perennes leñosas templados se adaptan a las estaciones modulando su crecimiento y desarrollo. Mientras que se desarrollan durante la primavera y el verano, dejan de crecer durante el otoño para ir latentes en invierno1. Aunque la inactividad les permite sobrevivir a bajas temperaturas del invierno, escalofriante es un requisito previo para una adecuada brotación en primavera2. Las implicaciones importantes de inactividad en la producción de frutas de clima templado y bosques han provocado diversos esfuerzos para determinar y predecir el período de dormancia3. En especies frutales, experimentos empíricos transferencia brotes a forzar las condiciones y pronósticos estadísticos basados en datos de la floración son enfoques actuales para determinar la fecha de la ruptura del reposo invernal, que permite a los investigadores estimar la escalofriante los requisitos para cada cultivar. Sin embargo, cómo determinar el estado de latencia basado en procesos biológicos sigue siendo confuso3.
Floración en frutales templados, como la cereza dulce (Prunus avium L.), se produce una vez al año y dura aproximadamente dos semanas. Sin embargo, flores empiezan a distinguir y desarrollar unos 10 meses antes, durante el verano anterior4. Primordios de flores dejen de crecer durante el otoño para permanecer latente dentro de los brotes durante el invierno. En este período, cada cultivar tiene que acumular un requisito escalofriante especial para floración adecuada4. A pesar de la falta de cambios fenológicos en los brotes durante el invierno, primordios de flores son fisiológicamente activos durante la latencia, y la acumulación de temperaturas de refrigeración se ha asociado recientemente a la dinámica de acumulación de almidón o disminuir dentro de las células del primordio de ovario, que ofrece un nuevo enfoque para la determinación de latencia5. Sin embargo, el pequeño tamaño y la ubicación del primordio de ovario requieren una metodología especial.
El almidón es el carbohidrato principal de almacenamiento en plantas leñosas especies6. Por lo tanto, cambios en almidón han sido relacionados con la actividad fisiológica de los tejidos de la flor, que necesitan carbohidratos para apoyar su desarrollo7,8. Diferentes eventos claves durante el proceso reproductivo también están relacionadas con variaciones en el contenido de almidón en diferentes estructuras florales, como antera meiosis9, el crecimiento de los tubos de polen a través de la fertilización de estilo u óvulo10. Técnicas histoquímicas permiten la detección de almidón en cada tejido en particular de los primordios de flores durante la latencia. Sin embargo, la dificultad sigue siendo en la cuantificación de ese almidón para permitir después su patrón de acumulación o disminuir con el tiempo o comparar el almidón contenido entre tejidos, cultivares o años. Esto es debido a la poca cantidad de tejido disponible para el11de técnicas analíticas. Como alternativa, análisis de imagen vinculados a microscopia12 permite la cuantificación del almidón en muy pequeñas muestras de tejido de planta13.
Enfoques combinando análisis de microscopía e imagen se han utilizado para cuantificar el contenido de diferentes componentes en los tejidos vegetales, como el Callosa14, microtubos15, o16, midiendo el tamaño de la zona teñida por específico del almidón manchas. Para el almidón, puede ser fácilmente detectada mediante el yodo yoduro de potasio (I2KI) reacción17. Este método es altamente específico; I2KI intercala dentro de la estructura laminar de los granos de almidón y se forma un color azul o marrón rojizo oscuro, dependiendo del contenido de amilosa del almidón18. Secciones teñidas con mi mancha KI2Mostrar contraste adecuado entre el almidón y el tejido de fondo, permitiendo una detección inequívoca del almidón y la posterior cuantificación por el de sistema de análisis de imagen19. Aunque este colorante no es estequiométrica, la acumulación de yodo es proporcional a la longitud de la molécula de almidón, que puede variar muy17. Así, el tamaño de la zona manchada que se expresa como el número de píxeles puede no reflejar con precisión el contenido de almidón, ya que altas diferencias en contenido de almidón podrían encontrarse entre campos con áreas manchadas de tamaño similar. Como alternativa, el contenido de almidón puede ser evaluado midiendo la densidad óptica de los gránulos teñidos en blanco y negro imágenes obtenidas en el microscopio, como ha sido reportado en diversos tejidos en albaricoque8,13 , 19, aguacate10,20y oliva21.
Aquí, describimos una metodología que combina la determinación experimental del estado de latencia con la cuantificación del contenido de almidón en el tejido del primordio de ovario desde el otoño a la primavera en cerezas, ofreciendo una nueva herramienta para la comprensión y predicción de latencia basado en el estudio de los mecanismos biológicos relacionados con dormancia.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. colección Material determinación latencia y planta
- Brotes de la flor en el campo de la muestra. Estudios de latencia son experimentos a largo plazo y requieren adultos árboles suficientemente grandes para recoger los cogollos y brotes durante todo el invierno sin comprometer el desarrollo de los árboles durante la primavera siguiente. Manejo de huerta especial puede ser requerido según el sistema de formación; así, la poda puede ser menos severa que para fines de producción de fruta.
- Cada semana, desde el inicio del otoño hasta el inicio de la brotación, recoger y pesar 10 brotes de flor.
- Fijar los botones florales, colocando en un tubo de vidrio de 10 mL con una tapa y tomar las muestras en una solución fijadora de etanol/ácido acético (3:1) durante al menos 24 h a 4 ° C. Luego, deseche el fijador y añadir etanol del 75%, generosamente asegurando que cubre las muestras. Las muestras pueden conservarse en esta solución a 4 ° C hasta su uso.
Nota: Infiltración de vacío se puede utilizar para eliminar las burbujas de aire dentro de la yema y evitar que flote. Esto facilita la penetración del fijador en los tejidos pero podría dañar la estructura de los tejidos. Tratar de evitar si no es necesario.
- Los brotes en el campo de la muestra.
- Cada semana, desde el inicio del otoño hasta el inicio de la brotación, tomar tres sesiones de 15-30 cm de longitud y 5 mm de diámetro, que contiene por lo menos 10 brotes de flor. Colóquelas en la espuma de Florista empapada en agua en una cámara de crecimiento a 22 ± 1 ° C con un fotoperiodo de 12 h luz.
- Después de 10 días en la cámara de crecimiento, seleccionar y pesar 10 capullos de los brotes.
- Evaluar el crecimiento del brote y determinar el estado de latencia. El período de muestreo ha de ajustarse a las condiciones de la situación. En las condiciones de la huerta (Zaragoza, España, 41 ° 44'30 "N, 0 ° 47'00" W y 220 m sobre el nivel del mar), el muestreo de los brotes se llevó a cabo del 30 de noviembre hasta finales de febrero o principios de marzo.
- Cada semana evaluar la respuesta de brotes de la flor a las condiciones de crecimiento adecuado de la cámara, desde el principio del otoño hasta el rompimiento de la yema en la primavera, comparando el peso de los 10 cogollos recogidos del campo.
- Si existen diferencias o esas diferencias son menos del 30%, considera que los brotes no han cumplido con sus requerimientos de frío y endodormant22. Si las diferencias son más del 30%, consideran que los brotes han cumplido con sus requerimientos de frío y ecoletargo22.
2. planta preparación Material para la cuantificación de almidón
- Seleccionados brotes fijados unos seis de cada muestreo fechan (vea el paso 1.1). Dependiendo de los cultivares, cada brote de flor de la cereza dulce contiene hasta cinco primordios de flores.
- Eliminar las escamas de la yema externo y colocar la yema en el vidrio de un relojero con etanol al 75% para evitar que se resequen.
- El brote de disecar y extraer al menos un primordio de flores por yema con la ayuda de pinzas de precisión y un bisturí oftalmológico en un microscopio estereoscópico. El primordio de la flor puede conservarse en un tubo de vidrio de 10 mL con etanol al 75% a 4 ° C, o proceder de inmediato al siguiente paso.
- Deshidratan las muestras en una serie de alcohol butílico terciario.
- Reemplazar la solución de etanol 75% con 10 mL de alcohol butílico terciario del 75% (TBA) para cubrir generosamente las muestras e incúbelos durante 1,5 h. Una pipeta Pasteur puede ser útil para descartar la solución.
- Repita el paso 2.2.1, adición de TBA con concentraciones crecientes (85%, 95% y 100%, v/v; y 3 x con TBA pura) en una estufa de secado a 30 ° C con extracción de aire, ya que TBA puro se cristaliza a temperatura ambiente (< 20 ° C) y es muy volátil y tóxico. Si TBA se cristaliza con la muestra, dañaría los tejidos.
- Incrustar las muestras en parafina18.
- Derretir la parafina mediante la introducción de las perlas de la parafina en la estufa de secado a 60 ° C con extracción de aire el día anterior. Las perlas de parafina pueden también derretir sobre una placa, pero aseguran de que la parafina líquida es a 60 º C y no en una temperatura más alta, para evitar daño a los tejidos.
- Reemplazar la TBA con una mezcla de aceite de TBA y parafina (1:1) e incubar por 24 h en una estufa de secado a 60 ° C. Luego, vuelva a colocar la mezcla de TBA y aceite de parafina con parafina pura fusión e incubar durante al menos 6 h en una estufa de secado a 60 ° C. Repetir x 2 e incubar el último cambio para por lo menos 4-6 d.
- Coloque cada muestra en un pequeño molde base metal sobre una superficie de calor, embebidos en parafina y ritmo el cassette de inclusión. Coloque sobre una superficie fría y retire el bloque una vez que la cera se solidifica.
- Sección y rehidratar los preparativos.
- Preparar el adhesivo de Haupt: disolver 1 g de gelatina de Knox llano en 100 mL de agua destilada a 30 ° C; Luego, añadir 2 g de fenol cristales sueltos (C6H5OH) y 15 mL de glicerol. Extienda el adhesivo de Haupt sobre un portaobjetos de vidrio con un pincel y añadir una gota de una solución de formaldehído 1%.
- Sección cada parafina bloque a 10 μm en un micrótomo rotatorio y colocar los elementos en la diapositiva de cristal cubierto con el pegamento de Haupt.
- Coloque el portaobjetos con las secciones sobre una superficie de calor a 35-40 ° C hasta que se seque; después, mueve el portaobjetos a una parrilla.
- Preparar las soluciones dewax y rehidratación. Colocar 200 mL de Histoclear II en tres platos de tinción de cristal, otro con Histoclear II:ethanol (1:1, v/v) y una serie de etanol (100%, 70%, 40%, v/v) y un lavado final con agua destilada.
- Dewax las secciones con tres lavados, cada 5 min, Histoclear II y el Histoclear II:ethanol (1:1, v/v). Coloque la parrilla en los platos manchas de vidrio, asegurando que la solución cubre completamente las diapositivas y, luego, mueva la parrilla corrediza de una solución a la siguiente.
- Rehidratar las secciones de la siguiente serie de lavados de 2 minutos: una serie de etanol (100%, 70%, 40%, v/v) y un lavado final con agua destilada. Finalmente, secar los portaobjetos de vidrio a temperatura ambiente.
- Los artículos18de la mancha.
- I preparar la mancha KI2: disolver 2 g de yoduro de potasio (KI) y 0,2 g de yodo (I2) en 100 mL de agua destilada. Aplique una gota de dulce I2KI sobre las secciones de 5 minutos y, a continuación, deseche el exceso de tinción absorbiendo con un papel secante. Proceder rápidamente al siguiente paso.
- Aplique una pequeña gota de un medio de montaje sintético, coloque un vaso pequeño de la tapa en la parte superior y presione con fuerza. Una vez que se seca en los medios de montaje, observar bajo un microscopio de campo brillante para una evaluación preliminar del almidón de ovario.
Nota: Este paso no es obligatorio, aunque se obtiene un mejor contraste entre el almidón y el fondo. Si la sección tiene que ser reutilizado con otras manchas después de la cuantificación de almidón, coloque el cubierta de vidrio sobre la gota de colorante y deseche el exceso absorbiendo con un papel secante (véase 2.5.1). Luego, después de la cuantificación de almidón, lavar a I2KI la mancha con agua destilada y coloque la preparación sobre un fuego superficial a 35-40 ° C hasta que se seque.
3. cuantificación del contenido de almidón
- Calibrar las condiciones ópticas.
Nota: Los niveles de detección utilizados el analizador de imagen para detectar el almidón teñido son directamente dependientes de las condiciones de luz y magnificación del microscopio; por lo tanto, fijar estas condiciones para todas las preparaciones evaluadas. Adaptar el ajuste propuesto aquí para el microscopio disponible y las condiciones de luz.- Ajustar el diafragma de apertura en el 20 X aumentos y el control de brillo o intensidad de la luz.
- Asegúrese de que no hay ningún filtro en el soporte del filtro y seleccione una condición de campo brillante en el microscopio. Seleccione un aumento adecuado (p. ej., 40 X para cerezas primordio de ovario).
- Control de las condiciones de adquisición de imagen. Ajustar la configuración de la cámara con una preparación teñida sin tejido vía imagen | Adquisición | Vista previa.
- Fije el brillo al 50%, la ganancia en 1.0 x y los indicadores de histograma la gamma valor en 1.00 y el contraste en el 0 - 100, alineando con los límites del histograma de distribución de brillo.
- Activar la función de sobreexposición/subexposición y ajustar el tiempo de exposición en el límite de exposición.
- Aplicar la función de balance de blancos a la imagen completa para mostrar todos los componentes de la imagen sin ningún tono de color y la corrección del sombreado color neutro a la imagen completa para crear una imagen corregida y homogénea.
- Calibrar el sistema de análisis de imagen para obtener los valores de control del nivel de gris (0, negro; 255 blanco) de diferentes densidades ópticas (OD) que se obtienen los valores de transmitancia (T).
- Adquirir una imagen de una preparación teñida sin tejido, considerado control blanco y medir el nivel de gris de la imagen en blanco y negro a través de medida | Gris a medida | Calibra gris | Valor de referencia = 0 | Medida | Calibrar | OK. Esto se corresponde con una transmitancia de 100%; por lo tanto, una densidad óptica de 0, según OD = 2 - log T.
- Adquirir una imagen de la misma preparación con un filtro de 4N, que reduce la cantidad de luz de 4 x y medir el nivel de gris de la imagen en blanco y negro a través de medida | Gris a medida | Calibra gris | Valor de referencia = 0,6 | Medida | Calibrar | OK. Esto se corresponde con una 25% de transmitancia; por lo tanto, una densidad óptica de 0,6, según OD = 2 - log T.
- Adquirir una imagen de la misma preparación sin luz, considerado negro, y medir el nivel de gris de la imagen en blanco y negro a través de medida | Gris a medida | Calibra gris | Valor de referencia = 1 | Medida | Calibrar | OK. Esto se corresponde con una transmitancia de 0%; por lo tanto, una densidad óptica de 1, según OD = 2 - log T.
- Detectar el almidón.
- Adquirir una imagen a color del campo a medida en formato TIFF con una resolución de al menos 300 puntos por pulgada (PPP).
- Crear una imagen binaria correspondiente a la zona manchada. Establecer los límites de tres colores (valores entre 0 - 255 para cada uno) hasta que la imagen binaria refleja exactamente los gránulos de almidón teñidos observados, a través de imagen | Detectar | Seleccionar los umbrales de rojo, azul y verde | OK. Hacer comparaciones visuales repetidamente en varias preparaciones y tejidos para ajustar los niveles de detección final. Almacenar y usar estos niveles para todas las preparaciones.
- Cuantificar el almidón. Convertir la imagen original en color en una imagen en blanco y negro con el sistema de análisis de imagen. Utilizar la imagen binaria como una máscara superpuesta en la imagen en blanco y negro a través de imagen | Binary Edit. Medir la suma de la densidad óptica de cada píxel en la máscara a través de medida | Nivel de gris | OK y considerar este valor como el contenido de almidón en el campo medida.
- Repita los pasos 3.4.1 - 3.4.3 en cuatro lugares de los primordios de ovario para obtener un valor representativo de los contenidos de almidón en el ovario de primordios de flores.
- Repita los pasos 3.4.1 - 3.4.3 y paso 3.4.5 en flores diferentes de cada fecha de recolección.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Estudios de latencia requieren la determinación del momento cuando se cumplen los requisitos de refrigeración. A pesar de la falta de cambios fenológicos durante el invierno en condiciones de campo (figura 1A), cerezos no recuperan la capacidad de crecimiento en condiciones adecuadas hasta que pasen cierto periodo bajo temperaturas bajas. La transferencia regular de brotes a una cámara de condiciones controladas (figura 1B) durante el invierno permite la evaluación del estado de latencia de las yemas de flor. La evaluación del crecimiento del brote de flor se realizó midiendo el aumento en peso de yema. Mientras que, durante la latencia, no pudieran observarse cambios después de 10 días de las condiciones adecuadas (figura 1), fue superada la latencia, los cogollos se hincharon y estallan en la cámara de crecimiento (figura 1). Los resultados de este análisis permitieron el estado de dormancia de las yemas que se establecerá. Debido a las diferentes temperaturas durante el invierno, la latencia fue superada en distintas fechas, dependiendo del año. Mientras que, durante el primer año del estudio, la ruptura de la Dormición ocurrió en enero, el segundo año presenta un invierno más suave; así, el cumplimiento escalofriante ocurrió unas tres semanas después, en febrero.
Cereza dulce lleva capullos en espolones, donde la yema apical es un brote vegetativo y los brotes laterales son capullos (figura 1C y 1D). Brotes indiferenciados comenzaron a diferenciarse en flores o brotes vegetativos en el final del verano, y cuando entran en latencia en invierno, varios primordios de flores permanecen dentro de la yema, protegido por numerosas escamas verdes y cubiertas por escamas marrones externo ( Figura 1 y figura 2A). La disección de la yema floral mostró los primordios de flores pequeño interior (figura 2A). Cada yema floral había contenida primordios de flores individuales de uno a cinco (figura 2B). A pesar del pequeño tamaño de cada primordio de flor, todas las partes de una flor se distinguen y se pueden distinguidas: los sépalos (figura 2), los pétalos, las anteras y el pistilo. El uso de técnicas histoquímicas (alcohol: acética [3:1] fijación inclusión de parafina, microtomo de seccionamiento y tinción basada en yodo almidón) permite la distribución del almidón dentro de la flor de los tejidos del primordio ser observado (Figura 2D ).
Almidón en el primordio de ovario fue cuantificado en cada sección. Cuatro medidas de 1337 μm2, en el aumento de 40 x, representan la disposición general del almidón en el primordio de ovario cereza dulce (Figura 3A). Gránulos de almidón se distingue claramente del fondo después de I2KI tinción (figura 3B). El almidón fue identificado por el sistema de análisis de imagen, ajustar el color de los umbrales de rojo, verde y azul hasta que todos los gránulos de almidón fueron cubiertos por la imagen binaria creada por el sistema basado en los parámetros de color definido (figura 3). Los valores de contenido de almidón obtenidos fueron el resultado de la medida de la densidad óptica de cada píxel bajo la máscara de la imagen en blanco y negro (figura 3D).
La cuantificación de almidón reveló un patrón consistente de almidón dinámico durante el invierno (figura 4). Constantemente, la cantidad de almidón en invierno presenta un valor de densidad óptica de menos de 40.0003, mientras que la cantidad máxima alcanzó un valor de entre 120.000 y 140.000 en ambos años. Mientras que el valor máximo se alcanzó en enero durante el primer año (Figura 4A), ocurrió en febrero en el segundo año (Figura 4B). Contrastando estos resultados con la tez de la latencia, la cantidad máxima de almidón ocurrió concomitante con el escalofriante cumplimiento en ambos años.
Este enfoque requiere la determinación de la situación de inactividad (figura 5A) concomitante con la cuantificación de almidón en los tejidos del ovario (figura 5B) con el fin de enmarcar los cambios en el contenido de almidón en relación con la inactividad.

Figura 1: montaje Experimental para la determinación del estado de dormancia de las yemas de flor de cereza dulce. (A) ramas, durante el invierno, muestran las yemas en reposo cerrado y cubierto por escamas de color marrón oscuros. (B) este panel muestra a la cámara de crecimiento de brotes. A mediados de enero, algunos cultivares permanecidos inactivos con los brotes aún cerraron (flecha blanca), mientras que otros eran capaces de crecer, mostrando brotes hinchados (flecha negra). (C) este panel muestra un detalle de una sesión con capullos latentes situados lateralmente y una sola yema de madera situado en la posición apical en el estímulo. (D) este panel muestra un detalle de una sesión de inactividad fue superada después de 10 días en la cámara de crecimiento, que muestra hinchazón de yemas. Haga clic aquí para ver una versión más grande de esta figura.
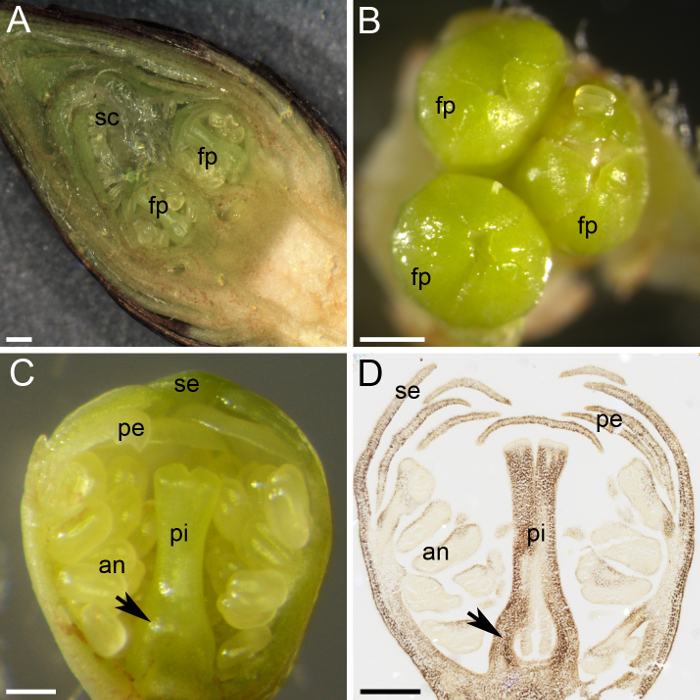
Figura 2: preparación de material para la cuantificación de almidón de la planta. (A) este panel muestra una sección transversal de un brote de flor, mostrando dos primordios de flores (PF) protegida por numerosas escamas (sc). (B) tres primordios de flores (PF) se reúnen en un brote de flor. (C) este panel muestra una sección transversal de un primordio de flor, con los verticilos diferenciados: sépalos (se), pétalos (pe), anteras (an) y pistilo (pi). El primordio de ovario se distingue en la base del pistilo (flecha). (D) una sección media de un primordio de flor fue recogido fijo en enero, incrustado en la cera de parafina, seccionada y manchado i2KI (marrón oscuro) para el almidón. Este panel muestra el primordio de ovario (flecha). Las barras de escala son 500 μm en paneles A y B y 200 μm en los paneles C y D. Haga clic aquí para ver una versión más grande de esta figura.

Figura 3: cuantificación en primordio de ovario cereza dulce de almidón. (A) este panel muestra una sección media de un primordio de ovario manchado i2KI, mostrando los cuatro marcos en que almidón contenido fue medido. (B) este panel muestra un detalle del primordio de ovario. Los gránulos de almidón se tiñen de color marrón oscuro. (C) este panel muestra una imagen pseudocolor que almidón corresponde a diferentes tonos de azul. (D) este panel muestra una máscara de imagen binaria que me almidón teñidos de KI2(azul) en la imagen original blanco y negro. La densidad óptica se mide sólo en los píxeles de la imagen original cubierto por la máscara. Las barras de escala son 100 μm en panel A y 20 en paneles B - D. Haga clic aquí para ver una versión más grande de esta figura.

Figura 4: resultados representativos de la cuantificación de almidón en primordios de ovario cerezas recogidas mensualmente desde el otoño a la primavera durante dos años de las condiciones de temperatura de invierno diferentes. (A) este panel muestra los resultados de los años 2010-2011, que tuvo un invierno frío. El cumplimiento escalofriante (copo de nieve) se produjo en enero, concomitante con la máxima cantidad de almidón. (B) este panel muestra los resultados de los años 2011-2012, que tuvo un invierno suave. El cumplimiento escalofriante (copo de nieve) se produjo en febrero, concomitante con la máxima cantidad de almidón. Los valores son la media ± el error estándar de la media. Haga clic aquí para ver una versión más grande de esta figura.
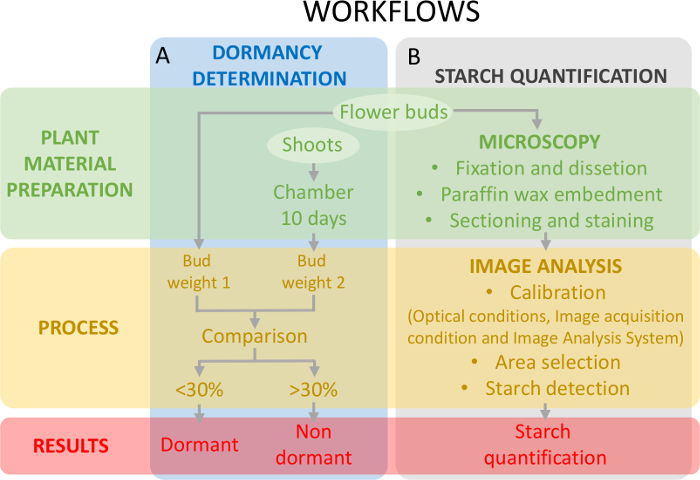
Figura 5: esquema del diseño experimental para evaluar el estado de latencia de yemas y la cuantificación de almidón en los primordios del ovario en cereza dulce. (A) este panel muestra los flujos de trabajo de la determinación de la condición de inactividad: la preparación del material de planta, el proceso y los resultados obtenidos. (B) este panel muestra los flujos de trabajo de la cuantificación de almidón: la preparación histoquímica de los cogollos de una observación microscópica de la cuantificación de almidón, el almidón y la detección de análisis de imagen de almidón. Haga clic aquí para ver una versión más grande de esta figura.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Dormición en plantas perennes leñosas presenta claras implicaciones en producción frutícola y forestal en un clima cambiante, aunque el proceso biológico detrás de latencia sigue siendo confuso. Estudios de dormancia pueden ser abordados desde diferentes puntos de vista, pero la investigación en busca de un marcador biológico para la dormancia invernal se ha intensificado en los últimos años. Sin embargo, la mayoría intenta encontrar un inequívoco indicador que muestra Cuándo un brote ha roto latencia ha sido fracasado3. La metodología descrita en este documento, la combinación de técnicas histoquímicas18 con análisis de imagen, ha sido muy útil para examinar la relación entre las reservas de hidratos de carbono de un tejido particular y su actividad fisiológica en yemas de cerezo dulce durante las diferentes fases de latencia5 y puede aplicarse también a otras especies y tejidos10,13,20.
Las reservas de carbohidratos en forma de almidón juegan un papel importante tanto en el desarrollo de la flor y el proceso reproductivo7,10,13,20,21,23 y en la estacionalidad de los perennes leñosos templado6. Diferentes estudios sobre latencia han prestado atención al almidón dentro de los cogollos24. Sin embargo, debido a su pequeño tamaño, varias yemas enteras son necesarios para el uso de métodos analíticos cuantitativos, y la detección de las variaciones cuantitativas en particular tejidos o células está limitada por el efecto de enmascaramiento de las células circundantes. La combinación de técnicas histoquímicas con el sistema de análisis de imagen ofrece una buena oportunidad para estudiar los cambios en el contenido de almidón de diferentes estructuras dentro de la yema.
Este método tiene la limitación de prevenir la cuantificación del contenido exacto de almidón en los tejidos pero permite que el contenido en almidón relativa a cuantificarse cambios almidón cuantitativo sobre tiempo25 y comparar el almidón contenido de diferentes tejidos13,20, cultivares o años5. Para permitir la correcta comparación de los valores de densidad óptica entre campos, los tejidos y brotes, la calibración del sistema (condiciones de luz, tinción intensidad y ampliación) y el ajuste de color se establecerá con precisión los umbrales , almacenados y utilizados para todas las preparaciones.
Manchas de almidón y cuantificación basado en yodo de potasio permite el uso posterior de otras manchas después del lavado de la sección. Así, diferentes análisis pueden realizarse sin preparación adicional y montaje sintético los medios no son usados de5,20. Asimismo, morfométricas medidas26 puede hacerse después de la preparación del mismo, permitiendo que el patrón de acumulación de almidón a enmarcarse en relación con el crecimiento de diferentes estructuras5,13. El método se puede adaptar a otras estructuras o especies con el ajuste de los niveles de color que utilice el analizador para detectar el almidón, que puede permitir el estudio de otros procesos de desarrollo que implican cambios en el contenido de almidón en pequeños grupos de células.
La relación entre la liberación de la latencia y el almidón de la acumulación en los primordios del ovario por el uso de este método proporciona una base sólida para entender la base biológica de la latencia y escalofriante requisitos5. Sin embargo, la cuantificación de almidón por análisis de imágenes de secciones parafina-encajadas podría llegar a ser muy engorrosa y requiere mucho tiempo estimar las necesidades de refrigeración de un gran número de cultivares. Futuros esfuerzos tienen que centrarse en el estudio de indicadores biológicos confiables que fácilmente pueden indicar el estado de dormancia del árbol. Mientras tanto, el patrón de variaciones de almidón durante la latencia se puede utilizar para otros estudios fisiológicos y genéticos del marco, y la combinación de técnicas histoquímicas con análisis de imagen descritos podría utilizarse en otros cultivos perennes leñosas a cuantificar el contenido de almidón de diferentes tejidos en relación con la inactividad.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Los autores no tienen nada que revelar.
Acknowledgments
Los autores agradecen con gratitud María Herrero y Eliseo Rivas por su útil discusión y consejos. Este trabajo fue financiado por el Ministerio de Economía y Competitividad — Fondo Europeo de Desarrollo Regional, la Unión Europea [número de concesión BES-2010-037992 para E. F.]; el Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria [números de concesión RFP2015-00015-00, RTA2014-00085-00, RTA2017-00003-00]; y el Gobierno de Aragón, Fondo Social Europeo, la Unión Europea [Grupo Consolidado A12-17R].
Materials
| Name | Company | Catalog Number | Comments |
| Precision scale | Sartorius | CP225D | |
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Drying-stove | Memmert | U15 | |
| Paraffin Embedding station | Leica Microsystems | EG1140H | |
| Rotatory microtome | Reichert-Jung | 1130/Biocut | |
| Microtome blade | Feather | S35 | Stainless steel |
| Bright field microscope | Leica Microsystems | DM2500 | |
| Digital Camera | Leica Microsystems | DC-300 | |
| Image Analysis System | Leica Microsystems | Quantiment Q550 |
References
- Kurokura, T., Mimida, N., Battey, N. H., Hytönen, T. The regulation of seasonal flowering in the Rosaceae. Journal of Experimental Botany. 64 (14), 4131-4141 (2013).
- Rohde, A., Bhalerao, R. P.
- Fadón, E., Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environmental and Experimental Botany. 152, 28-36 (2018).
- Fadón, E., Rodrigo, J., Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees - Structure and Function. , In Press (2018).
- Fadón, E., Herrero, M., Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Frontiers in Plant Science. 9 (171), (2018).
- Loescher, W. H., Mccamant, T., Keller, J. D. Carbohydrate reserves, translocation and storage in woody plant roots. HortScience. 25 (3), 274-281 (1990).
- Hedhly, A., et al. Starch turnover and metabolism during flower and early embryo development. Plant Physiology. , (2016).
- Rodrigo, J., Hormaza, J. I., Herrero, M. Ovary starch reserves and flower development in apricot (Prunus armeniaca). Physiologia Plantarum. 108 (1), 35-41 (2000).
- Julian, C., Rodrigo, J., Herrero, M. Stamen development and winter dormancy in apricot (Prunus armeniaca). Annals of Botany. 108 (4), 617-625 (2011).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil starch reserves at anthesis correlate with final flower fate in avocado (Persea americana). PLoS One. 8 (10), e78467 (2013).
- Smith, A. M., Zeeman, S. C. Quantification of starch in plant tissues. Nature Protocols. 1 (3), 1342-1345 (2006).
- Eliceiri, K. W., et al.
- Rodrigo, J., Herrero, M. Influence of intraovular reserves on ovule fate in apricot (Prunus armeniaca L.). Sexual Plant Reproduction. 11, 86-93 (1998).
- Zhou, J., Spallek, T., Faulkner, C., Robatzek, S. CalloseMeasurer: A novel software solution to measure callose deposition and recognise spreading callose patterns. Plant Methods. 8 (1), (2012).
- Faulkner, C., et al. An automated quantitative image analysis tool for the identification of microtubule patterns in plants. Traffic. 18 (10), 683-693 (2017).
- Kuhn, B. F. Determination of starch in ovules of the sour cherry cv. "Stevnsbaer.". European Journal of Horticultural Science. 71 (3), 120-124 (2006).
- Johansen, D. A. Plant microtechnique. , McGraw-Hill. New York, NY. (1940).
- Ruzin, S. E. Plant microtechnique and microscopy. , Oxford University Press. New York, NY. (1999).
- Rodrigo, J., Rivas, E., Herrero, M. Starch determination in plant tissues using a computerized image analysis system. Physiologia Plantarum. 99 (1), 105-110 (1997).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Ovary starch reserves and pistil development in avocado (Persea americana). Physiologia Plantarum. 140 (4), 395-404 (2010).
- Suarez, C., Castro, A. J., Rapoport, H. F., Rodriguez-García, M. I. Morphological, histological and ultrastructural changes in the olive pistil during flowering. Sexual Plant Reproduction. 25, 133-146 (2012).
- Lang, G. A., Early, J. D., Martin, G. C., Darnell, R. L. Endodormancy, paradormancy, and ecodormancy - Physiological terminology and classification for dormancy research. HortScience. 22 (3), 371-377 (1987).
- Hedhly, A., Vogler, H., Eichenberger, C., Grossniklaus, U. Whole-mount clearing and staining of arabidopsis flower organs and siliques. Journal of Visualized Experiments. 2018 (134), 1-10 (2018).
- Kaufmann, H., Blanke, M. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. Journal of Plant Physiology. 218 (July), 1-5 (2017).
- Herrero, M., Dickinson, H. G. Pollen-pistil incompatibility in Petunia hybrida: changes in the pistil following compatible and incompatible intraspecific crosses. Journal of Cell Science. 36, 1-18 (1979).
- Carpenter, A. E., et al. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biology. 7 (10), R100 (2006).
