

Summary
組織化学的手法と組み合わせる画像解析システムを使用して、冬の休眠中に甘果オウトウ (セイヨウミザクラL.) 卵巣原基のデンプン含量を定量化する手法を提案します。
Abstract
微小構造におけるでんぷんの変化は、受粉から受精・結実の発症に生殖期を含むいくつかの植物発達過程における主要なイベントに関連付けられます。しかし、花芽分化の進行中にでんぷんの変化知られていない完全に、花原基の特に小さい構造のデンプン含量を定量化の難しさのために主に。ここでは、に関するさまざまなフェーズとデンプン含量の変化を可能にする顕微鏡に接続されている画像解析システムを使用して甘いチェリー (Prunus avium l.) の卵巣原基における澱粉の定量化のため手法について述べる秋から春に休眠。この目的のため、花芽の休眠状態は冬時間の別の瞬間に、制御された条件に譲渡の芽の芽の成長を評価することによって決定されます。卵巣原基における澱粉の定量化, 花芽が順番に収集、固定、パラフィンに埋め込まれた、断面、私で染色2Kl (ヨウ素-ヨウ化カリウム)。準備は顕微鏡の下で観察を行い、背景から澱粉を明確に区別する画像解析によって分析します。デンプン コンテンツ値は検討フレームの澱粉含量の推定として各画素の光学濃度の合計を考慮したステンド グラスの澱粉に対応するイメージの光学密度を測定することによって取得されます。
Introduction
温帯木本は、その成長と発展を調節することによって、季節に適応します。彼らは春と夏の間作成し、彼らは冬1で休止状態に行く秋の間に成長を停止します。休眠により低い冬の温度で生き残るために、低温春2の適切な芽のための前提条件です。温帯果実生産・林業における休眠の重要な意味は、多様な取り組みを決定し、休眠期間3を予測するにつながっています。実証実験条件と開花のデータに基づいて統計的に予測を強制的に撮影を転送するは、フルーツの木の種を推定する研究を可能にする休眠打破の日付を決定する現在のアプローチ、各品種のための要件をゾッと。ただし、生物学的プロセスに基づく休眠状態を確認する方法不明3のままです。
甘果オウトウ (セイヨウミザクラl.) などの温帯果樹の開花は年に一度が発生し、約 2 週間続きます。ただし、花を区別し、前の夏4の間に約 10 ヶ月前を開発開始します。花原基は、冬の間に芽内休眠する秋の間に成長を停止します。この期間の各品種は、適切な開花4特定温要求量を蓄積する必要があります。冬の間に芽のフェノロジーの変化の欠乏にもかかわらず花原基が、休眠中に生理活性や低温処理温度の蓄積、最近デンプン蓄積の動的な関連付けられているまたは短縮卵巣原基の細胞内で休眠決定5の新しいアプローチを提供しています。ただし、サイズが小さいと卵巣原基の場所特別な方法論が必要です。
澱粉の木本種6の主要なストレージの炭水化物であります。したがって、でんぷんの変化は彼らの開発の7、8をサポートする炭水化物を必要とする花の組織の生理活性に関連しています。生殖のプロセス中に別のキー イベントも葯減数分裂9スタイルや胚珠の受精10まで花粉管の成長など、別の花の構造の澱粉含量の変化に関連しています。組織化学的技術は、休眠中、花原基のそれぞれの特定の組織にデンプンの検出を許可します。ただし、難易度は、そのデンプン含量の組織、品種、または年間次の蓄積/減少のパターンは時間の経過や澱粉を比較するようにの定量化に残ります。これは、組織分析手法11利用の少し量のためです。代わりに、顕微鏡12にリンクされている画像解析は、植物組織の13の非常に小さいサンプルの澱粉の定量化をことができます。
カロース14、チューブ15などの植物組織内の異なったコンポーネントの含量を定量化する特定の染色領域のサイズを測定することにより澱粉の16顕微鏡と画像解析を組み合わせたアプローチは使用されました。汚れ。でんぷん用簡単に検出できるヨウ化カリウム ヨウ素を使用して (私2KI) 反応17。この方法は非常に具体的です。私2KI デンプン粒のラミナー構造内主体しの暗い青や赤茶色の色を18の澱粉のアミロース含量によってを形作る。私と染色2KI 汚れ示す澱粉と背景組織画像解析システム19による明確なデンプン検出と後続の定量化が可能との間のコントラストが適切な。この染料は化学量論が、ヨウ素の蓄積は17を変わることができる高い澱粉分子の長さに比例。したがって、ピクセルの数として表された染色領域のサイズが内容が反映されない正確に澱粉、澱粉含量の高い違いは同じようなサイズのステンド グラス エリアとフィールドの間見つけることができるので。代わりに、デンプン含量はアプリコット8,13のさまざまな組織で報告されているように、顕微鏡から得られた黒と白の画像にステンド グラスの顆粒の光学密度を測定することによって評価される場合があります。,19、アボカド10、20、およびオリーブ21。
ここで、理解と予測のための新しいツールを提供している秋から春甘果オウトウ、卵巣原基組織の澱粉含量の定量を休眠状態の測定を組み合わせた手法を説明します。生物学的メカニズムの研究に基づく休眠、休眠とリンク。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 休眠決定と植物資料集
- フィールドの花のつぼみをサンプルします。休眠研究長期実験大人芽を収集するために十分な大きさの木および次のばねの間に木の開発を損なうことがなくすべての冬の撮影を必要とされます。特別な果樹園経営がトレーニング システムによって要求されます。したがって、剪定は果実生産用より重症度の低い可能性があります。
- 秋芽を破る、発病までの開始から、毎週収集し、花蕾が 10 の重量を量る。
- キャップ 10 mL ガラス管にそれらを配置することによって花の蕾を修正し、エタノール ・酢酸 (3:1) 4 ° C で、少なくとも 24 時間の固定液に試料を浸漬定着剤を破棄し、75% エタノール, 寛大サンプルをカバーしていることを確認を追加します。サンプルを使用するまで 4 ° C でこのソリューションで節約します。
注: 真空浸透は、芽の中の空気の泡を削除し、フローティングからそれらを防ぐために使用できます。これは組織への定着剤の浸透が容易になりますが、組織の構造に損傷を与える可能性があります。必要ではない場合、それを避けるようにします。
- フィールドでの撮影をサンプルします。
- 毎週、秋芽を破る、発病までの開始から、15-30 cm 長さ、5 mm 径、花蕾を少なくとも 10 を含む 3 つの芽を取る。22 ± 1 ° C 12 h 光日長成長チャンバ内の水に浸した花屋泡上に置きます。
- 成長室で 10 日後で拾うし、新芽から花芽の 10 の重量を量る。
- 芽の成長を評価し、休眠状態を確認します。サンプリング周期は場所の条件に調整すること。果樹園の条件で (スペイン、サラゴサ、41 ° 44'30"N, 0 ° 47'00"W と海抜 220 m)、シュートのサンプリングは、2 月の終わりか 3 月の初めまで 11 月 30 日から実施されました。
- 春、芽まで秋の初めから室の適当な生育条件をフィールドから選んだ 10 芽の重量を比較することによって花の蕾の応答を評価する毎週。
- 違いがないか、それらの違いは 30% 未満は、芽が、背筋が寒くなる要件に満たない、endodormant22であることを検討してください。違いは 30% 以上である場合は、芽が、背筋が寒くなる要件を満たしている、ecodormant22であることを検討してください。
2. 植物デンプン定量仕込み
- 各サンプリングから選択約 6 固定芽の日付 (手順 1.1 参照)。品種によっては、各甘い桜の花芽には最大 5 つの花原基が含まれています。
- 外部芽スケールを削除するし、乾燥からそれを防ぐために 75% エタノールを時計のガラスに芽を配置します。
- 芽を解剖し、精密鉗子のヘルプと実体顕微鏡下眼科メス芽あたりの少なくとも 1 つの花原基を抽出します。花原基は 4 ° c、75% エタノール 10 mL ガラス管に保存することができます。 または次のステップに進んでください。
- 第三ブチル アルコール シリーズのサンプルを脱水します。
- 75% エタノール溶液を 75% 三級ブチル アルコール寛大サンプルをカバーし、それらをインキュベート 1.5 h (未定) 10 mL に置き換えます。パスツール ピペットは、ソリューションを破棄することができます。
- 純粋なこの常温 (< 20 ° C) とは非常に揮発性で有毒なので空気抽出、30 ° C で乾燥ストーブで手順 2.2.1 上昇濃度 (85%、95%、100%、v/v; と純粋な未定 3 x) TBA を追加するを繰り返します。未定はこのサンプルと、組織を破損するしまうと。
- パラフィン18サンプルを埋め込みます。
- 空気抽出前の日に 60 ° C で乾燥のストーブでパラフィン真珠を導入することにより、パラフィンを溶かします。パラフィン真珠、ホット プレートにも溶かすことができるが、流動パラフィンが 60 ° c および組織の損傷を避けるためより高い温度ではないことを確認します。
- 置き換える、未定未定、パラフィン オイル (1:1) のミックスと 60 ° C で乾燥ストーブ内 24 時間インキュベートその後、未定のミックスを交換し、純粋なパラフィン オイル パラフィンを溶かすし、60 ° C で乾燥ストーブ内少なくとも 6 時間インキュベートそれを繰り返して 2 倍と、少なくとも 4-6 d の最後の変更を孵化させなさい。
- 熱表面に小さな金型ベースの各サンプルを配置パラフィン ワックス、および埋め込みカセット ペース埋め込まれています。冷たい表面上に配置し、ワックスが凝固したらブロックを削除します。
- セクションは、水分補給の準備。
- ハウプトの接着剤の準備: プレーンのノックス ゼラチン 30 ° c; 蒸留水 100 mL に 1 g を溶解2 g フェノール緩んで結晶 (C6H5オハイオ州) とグリセリン 15 mL を加えます。ハウプトの接着剤をブラシでスライド ガラスに広がる、1% ホルムアルデヒド溶液のドロップを追加します。
- セクション各パラフィン回転ミクロトームで 10 μ m でブロックし、スライド ガラス上にセクションを配置は、ハウプトの接着剤で覆われています。
- まで乾燥; 35-40 ° C で熱表面上のセクションにスライド ガラスを配置します。その後、スライド ガラスをスライド ラックに移動します。
- Dewax と補水ソリューションを準備します。3 ガラス染色料理、もう 1 つの Histoclear の II:ethanol (1:1、v/v)、エタノール シリーズに Histoclear II の 200 mL (100%, 70%, 40 %v/v)、および最終的な蒸留水洗浄。
- 3 つの洗浄、各 5 分、Histoclear II および Histoclear II:ethanol (1:1、v/v) でセクションを dewax します。確保ソリューションが完全にスライドを覆うガラス染色料理内スライド ラックを置き、その後、1 つのソリューションのスライド ラックを次に移動します。
- 2 分洗浄の次シリーズでセクションを水分補給: エタノール シリーズ (100%, 70%, 40 %v/v) と最終的な蒸留水洗浄。最後に、常温ガラス スライドを乾燥させます。
- セクション18を染色します。
- 私2KI 染色の準備: 2 g、ヨウ化カリウム (KI) とヨウ素の 0.2 g を溶解 (私2) 100 mL の蒸留水で。新鮮な滴を適用私25 分のセクション上氣と、あぶらとり紙で吸収することによって、汚れの余分は切り捨てます。すぐに次のステップに進みます。
- 合成実装メディアの小滴を適用の上に、小さなカバー ガラスを配置し、ハードを押します。一度メディアをマウントは乾燥、卵巣澱粉の予備的評価の明視野顕微鏡下で観察します。
注: 澱粉と背景のコントラストが得られるが、この手順する必須ではありません。セクションに他の汚れとデンプン定量後再利用することがある場合汚れのドロップ上カバーのガラスを置き、あぶらとり紙で吸収することによって余分は切り捨てる (2.5.1 を参照)。デンプン定量後、洗浄に2KI は蒸留水と場所熱上準備は 35-40 ° C で乾燥するまで表面ステインです。
3. 澱粉含量の定量
- 光の条件を調整します。
注: デンプンを検出する画像解析装置を用いた検出レベルは直接光の条件と、顕微鏡の倍率に依存してしたがって、評価のすべての準備のためのこれらの条件を修正します。利用可能な顕微鏡と光の条件をここで提案された調整を適応します。- X 倍率と明るさコントロールまたは光強度 20 で絞りを調整します。
- フィルター ホルダーにフィルターされていないことを確認し、顕微鏡で明るいフィールド条件を選択します。(例えば甘果オウトウ卵巣原基の 40 X) 適切な倍率を選択します。
- 画像取得条件を制御します。組織を介して画像なしステンド グラスの準備とカメラの設定を調整する |買収 |前に表示。
- 50%、1.0 の利得で明るさを修正 x とガンマ値 1.00 0 - のコントラスト、ヒストグラムの指標 100、輝度分布ヒストグラムの限界と並べる。
- 露出オーバー/アンダー関数をアクティブにし、露出オーバーの制限で露光時間を調整します。
- ホワイト バランス機能を修正と同質なイメージを作成する完全な画像に任意の色のトーンやシェーディング補正なしのイメージのすべての中立着色されたコンポーネントを表示する完全な画像に適用されます。
- 透過率 (T) の値によって取得される別の光学密度 (OD) の灰色のレベル (0、黒; 255、白) のコントロール値を取得する画像解析システムを調整します。
- 白、コントロールを考慮せず組織、染色準備のイメージを取得し、黒と白のイメージを介してメジャーの灰色レベルを測定 |灰色の測定 |グレーの調整 |基準値 = 0 |メジャー |校正 |OK。これは 100% の透過率; に対応します。このように、OD によると 0 の光学密度 = 2 - t. をログに記録
- 4 光の量を減らす 4 n フィルターと同じ準備のイメージを取得 x、および測定の黒と白のイメージを介してメジャー グレー レベル |灰色の測定 |グレーの調整 |基準値 0.6 を = |メジャー |校正 |OK。これは 25% の透過率; に対応します。このように、OD によると 0.6 の光学密度 = 2 - t. をログに記録
- 光、考え抜かれた黒なしと同じ準備のイメージを取得し、黒と白のイメージを介してメジャーの灰色レベルを測定 |灰色の測定 |グレーの調整 |基準値 = 1 |メジャー |校正 |OK。これは透過率が 0% で; に対応します。このように、OD によると、1 の光学密度 = 2 - t. をログに記録
- デンプンを検出します。
- 少なくとも 300 ドット/インチ (dpi) の解像度で TIFF 形式で測定するフィールドのカラー画像を取得します。
- 染色領域に対応するバイナリ イメージを作成します。3 色の閾値を設定 (0 - の間の値ごとの 255) バイナリ イメージをまさに反射を介して画像, デンプン顆粒まで |検出 |赤、青、緑のしきい値を選択 |OK。様々 な準備や組織最終的な検出レベルを調整する繰り返し視覚的比較を行います。格納し、すべての準備のためのこれらのレベルを使用します。
- 澱粉を定量化します。画像解析システムと黒と白の画像に元のカラー画像を変換します。黒と白のイメージにを介して画像重畳マスクとしてバイナリ イメージを使用 |バイナリ編集。マスクを介して測定の下のすべてのピクセルの光学濃度の合計を測定 |灰色のレベル |OK 、測定フィールド内デンプン含量としてこの値を考慮します。
- 3.4.1 - 花原基の卵巣のデンプン含量の代表的な値を取得する卵巣原基 3.4.3 で 4 場所の手順を繰り返します。
- 各収集日の別の花で 3.4.5 をステップ、手順 3.4.1 - 3.4.3 繰り返します。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
休眠研究は、低温の要件を満たしているとき瞬間の定量を必要とします。フィールド条件 (図 1 a) の下で冬の間に季節の変更の欠乏にもかかわらず彼らは低温下で一定期間を経過するまで、桜の木は適切な状態で成長の容量を回復しません。冬の間の管理された条件室 (図 1 b) にシュートの正規の転移は、花芽の休眠状態の評価を許可しました。花芽の成長の評価は、芽の重量の増加を測定することによって行われていた。一方、休眠中に変更できることを確認適切な条件 (図 1) の 10 日後休眠を克服したら、芽が膨らんだし、成長室 (図 1) でのバーストします。この分析の結果は、確立する芽の休眠状態を許可されます。異なる温度が冬の間に発生して、休眠は、年によっては異なる日付で克服されました。一方、研究の最初の年の間に、1 月に発生した休眠打破の 2 年目提示より穏やかな冬。したがって、低温の履行後、2 月に約 3 週間が発生しました。
甘果オウトウは、スパーズ、栄養芽は、頂芽、側芽が花蕾 (図 1と1 D) で花芽を負いません。未分化の芽は、夏の終わりに花や植物の芽に分化を開始し、芽、多数緑の鱗によって保護され、覆われて黒い外側スケール (内部にいくつかの花の原基が残って冬季に休眠を入力するとき図 1と図 2 a)。花蕾の解剖を示した (図 2 a) の中の小さな花の原基。各花のつぼみには、5 人に 1 人の個々 の花原基 (図 2 b) が含まれています。各花原基の小さなサイズにもかかわらず、花のすべての部分が区別され、識別することができます: 雌しべ、葯、花弁と萼片 (図 2)。化学技術の使用 (アルコール: 酢酸 [3:1] 固定、パラフィン ワックスを埋め込む、ミクロトーム切片、およびデンプンのヨウ素による汚損) (図 2 D 観察する原基組織花内デンプンの分布を許可).
各セクションは、卵巣原基における澱粉を定量しました。1337 μ m2、40 倍の倍率での 4 つの措置は、甘果オウトウ卵巣原基 (図 3 a) の澱粉の一般的なレイアウトを表されます。澱粉はあと背景と明確に区別された2KI (図 3 b) を染色します。澱粉はされた赤、緑、および青のしきい値のすべてのデンプン粒を観察するまでは、定義された色パラメーター (図 3) に基づくシステムで作成されたバイナリ イメージで覆われていた色を調整することで画像解析システムによって識別されます。得られるデンプン含量の値 (図 3 D) の黒と白のイメージのマスクの下のすべてのピクセルの光学濃度の測定の結果であった。
デンプンの定量では、澱粉 (図 4) 冬の間に動的の一貫したパターンを明らかにしました。一貫して、初冬のデンプンの量は、最大量まで 120,000 と 140,000 の間値両方の年で光学濃度値は 40,000 未満3を発表しました。一方、最大値は、(図 4 b) の 2 年目 2 月に発生した (図 4 a) の最初の年の 1 月に達されました。両年とも背筋が寒くなる履行を休眠の顔色とこれらの結果を対照的には、デンプンの最大量が付随して発生しました。
このアプローチ、休眠に関連してデンプン含量の変化をフレームに、付随して卵巣組織 (図 5 b) のデンプン定量を休眠状態 (図 5 a) の決定が必要です。

図 1: 甘い桜の花芽の休眠状態の決定のための実験の設定です。(A) 枝、冬の間に閉じられ、暗い茶色の鱗で覆われて休眠芽が表示。(B) このパネルは、芽の成長室に転送を示しています。1 月中旬には、いくつかの品種の芽の休眠のまま閉じたまま (白い矢印)、他が成長することができる間膨らんだ芽 (黒矢印) を表示します。(C) このパネルは横方向に位置する休眠花芽と拍車の根尖部の位置にある単一植物の芽の撮影の詳細を示しています。(D) このパネルを示しています撮影の詳細成長室で 10 日後休眠が克服された一度芽の腫脹を示します。この図の拡大版を表示するのにはここをクリックしてください。
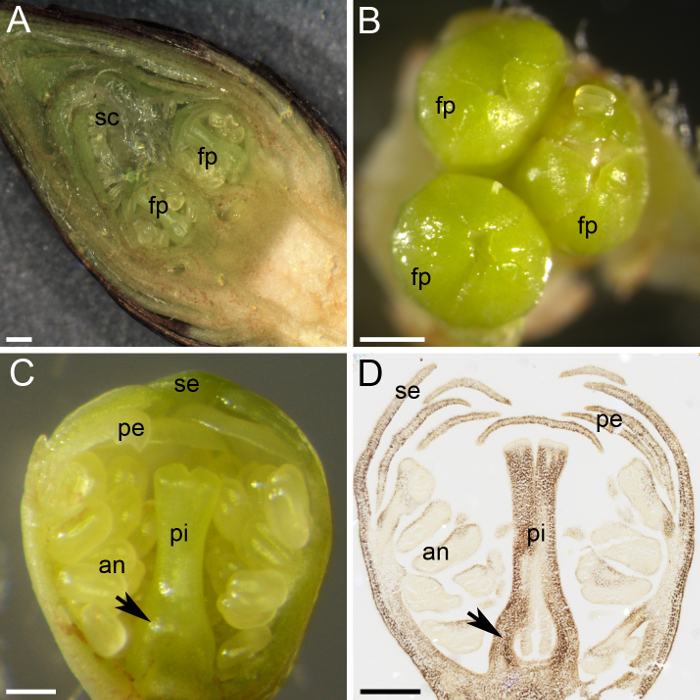
図 2: 植物のデンプン定量仕込みします。(A) このパネル多数スケール (sc) によって保護されて 2 つの花原基 (fp) を示す花芽の横断セクションを示しています。(B) 3 つ花原基 (fp) は、花のつぼみに集まります。(C) このパネルが花原基の横断セクションを示していますすべての渦巻きと分化: 萼片 (se)、花びら (pe)、葯 (、)、および雌ずい (pi)。卵巣原基は、雌しべ (矢印) の基地で区別されます。花原基の (D)、中段だった収集 1 月に固定、パラフィンに埋め込まれた、断面と私に染まった2デンプンの氣 (ダークブラウン)。このパネルには、卵巣原基 (矢印) が表示されます。スケール バーは、500 μ m パネルAとBとCとDのパネルで 200 μ m です。この図の拡大版を表示するのにはここをクリックしてください。

図 3: 澱粉の甘果オウトウ卵巣原基における定量化します。このパネル (A) 卵巣原基の中央セクションを示しています私に染まった2KI、どの澱粉中の 4 つのフレームを示す内容を測定しました。(B) このパネルは、卵巣原基の詳細を示しています。デンプン粒は、ダークブラウンに染色します。このパネル (C) 青の異なる色合いに対応する澱粉の擬似カラー画像が表示されます。(D) このパネルは、黒と白の元のイメージ (青)2KI ステンド澱粉をカバー私バイナリ イメージ マスクを示しています。光学濃度は、マスクで覆われて元の画像のピクセルでのみ測定されます。スケール バーは、パネルを100 μ m とB - Dのパネルで 20 μ m です。この図の拡大版を表示するのにはここをクリックしてください。

図 4: 甘果オウトウ卵巣原基のデンプン定量の代表の結果は異なる冬温度条件の 2 年間春秋から毎月収集します。寒い冬を持っていた (A) このパネル ショー 2010-2011 年の結果身も凍る履行 (スノーフレーク) は、澱粉の最大量と付随して、1 月で発生しました。(B) このパネルは、穏やかな冬があった 2012 年結果を示しています。2 月、付随してデンプンの最大量に背筋が寒くなる履行 (スノーフレーク) が発生しました。値は、平均 ± 標準誤差平均です。この図の拡大版を表示するのにはここをクリックしてください。
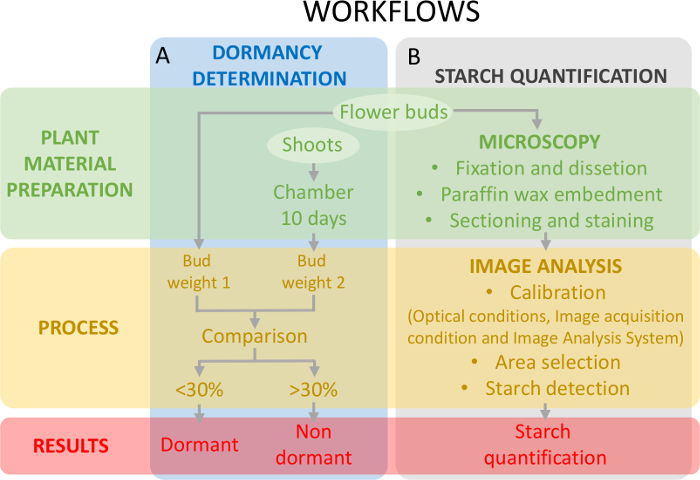
図 5: 芽と甘果オウトウ卵巣原基のデンプン定量の休眠状態を評価するために実験的なデザインのスキーム。(A) このパネルは、休眠状態の定量のワークフローを示しています: 植物材料の準備、プロセス、および得られた結果。(B) このパネル デンプン定量のワークフローを示しています: 澱粉、澱粉のイメージ分析検出およびデンプン定量の顕微鏡観察用の芽の組織化学的準備。この図の拡大版を表示するのにはここをクリックしてください。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
木質の多年生植物の休眠は休眠の背後にある生物学的プロセスは不明のままが果実生産の明確な含意と気候変動で森林を提示します。休眠研究は別の観点から近づくことができるが、最後の年にわたって冬の休眠の生物学的マーカーを探して研究を強めました。しかし、芽が休眠を壊れているときを示す明確な指標を見つけるほとんどの試みは失敗した3をされています。画像解析と組織化学的テクニック18を組み合わせること、記載方法は特定組織の貯蔵炭水化物と甘果オウトウの芽の生理活性の関係を明らかに非常に有用されています。休眠5のさまざまなフェーズで、他種と組織10,13,20に対しても適用できます。
デンプンの形で貯蔵炭水化物が花器の発育と生殖過程7,10,13,20,21,23の両方重要な役割を果たすと温帯木本6の季節。休眠に関する別の研究は、芽24内澱粉に注意を払っています。しかし、サイズが小さいためいくつかの全体の芽が定量分析法の使用のために必要な特定の組織や細胞の定量的変化の検出は、周囲のセルのマスキング効果によって制限されます。画像解析システムと組織化学的手法の組み合わせでは、芽の中の異なる構造の澱粉含量の変化を勉強する良い機会を提供しています。
このメソッドは組織内澱粉の正確な内容の定量化を防止する制限がありますのコンテンツを時間25以上定量的澱粉の変更を追跡して、澱粉を比較するために定量化する相対的なデンプンを可能異なったティッシュ13、20品種、または年5。フィールド、組織と芽の間の光学密度の値、(光の条件、染色の強さ、および倍率) システムのキャリブレーション、しきい値を正確に確立する必要があります色の設定の正しい比較を可能にするため、、格納され、すべての準備を使用します。
澱粉の染色とヨウ化カリウムに基づいた定量化は、セクションの洗浄後後の他の汚れの使用をことができます。このように、準備作業も新たに異なる分析を実行できる、メディアが合成の取り付け5,20。同様に、構造の異なる5,13の成長に関連してフレームにデンプンの蓄積パターンをできるように、同じ準備を経た形態測定26を行うことができます。メソッドは、他の構造体またはアナライザーを使用してセルの小さいグループのデンプン含量に変化を伴う他の発達過程の研究を許可することができます澱粉を検出する色レベルの調整と種に適応できます。
関係休眠リリースとデンプンの間このメソッドの使用によって明らかにされた卵巣原基の蓄積は休眠と低温の要件5の生物学的基盤を理解するための健全な基盤を提供します。ただし、パラフィン埋め込まれたセクションの画像解析による澱粉の定量化は、非常に面倒で多数品種の低温の要件を推定する時間がかかりになるかもしれない。今後の取り組みは、木の休眠状態を示すことができます簡単に信頼性の高いの生物学的指標を勉強に集中するあります。一方、休眠中のデンプンのバリエーションのパターンはさらに生理的・遺伝的研究をフレームに使用できするその他の木質の多年生作物で記載画像解析と化学技術の組み合わせを使用できます。休眠に関連して異なったティッシュのデンプン含量を定量化します。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
著者が明らかに何もありません。
Acknowledgments
著者は、マリア ・ エレーロと Eliseo リバスを役に立つ議論のアドバイス感謝してありがとうございます。この作品は Ministerio デ Economía y Competitividad によって支えられた、欧州地域開発基金、欧州連合 [許可番号 BES 2010 037992 e. f.];セルバンテス ナシオナル ・ デ ・危惧 y テクノロジー Agraria y Alimentaria [許可番号 RFP2015-00015-00 RTA2014-00085-00、RTA2017-00003-00];大統領府・ デ ・ アラゴン -欧州社会基金、欧州連合 [Grupo Consolidado A12-17R]。
Materials
| Name | Company | Catalog Number | Comments |
| Precision scale | Sartorius | CP225D | |
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Drying-stove | Memmert | U15 | |
| Paraffin Embedding station | Leica Microsystems | EG1140H | |
| Rotatory microtome | Reichert-Jung | 1130/Biocut | |
| Microtome blade | Feather | S35 | Stainless steel |
| Bright field microscope | Leica Microsystems | DM2500 | |
| Digital Camera | Leica Microsystems | DC-300 | |
| Image Analysis System | Leica Microsystems | Quantiment Q550 |
References
- Kurokura, T., Mimida, N., Battey, N. H., Hytönen, T. The regulation of seasonal flowering in the Rosaceae. Journal of Experimental Botany. 64 (14), 4131-4141 (2013).
- Rohde, A., Bhalerao, R. P.
- Fadón, E., Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environmental and Experimental Botany. 152, 28-36 (2018).
- Fadón, E., Rodrigo, J., Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees - Structure and Function. , In Press (2018).
- Fadón, E., Herrero, M., Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Frontiers in Plant Science. 9 (171), (2018).
- Loescher, W. H., Mccamant, T., Keller, J. D. Carbohydrate reserves, translocation and storage in woody plant roots. HortScience. 25 (3), 274-281 (1990).
- Hedhly, A., et al. Starch turnover and metabolism during flower and early embryo development. Plant Physiology. , (2016).
- Rodrigo, J., Hormaza, J. I., Herrero, M. Ovary starch reserves and flower development in apricot (Prunus armeniaca). Physiologia Plantarum. 108 (1), 35-41 (2000).
- Julian, C., Rodrigo, J., Herrero, M. Stamen development and winter dormancy in apricot (Prunus armeniaca). Annals of Botany. 108 (4), 617-625 (2011).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil starch reserves at anthesis correlate with final flower fate in avocado (Persea americana). PLoS One. 8 (10), e78467 (2013).
- Smith, A. M., Zeeman, S. C. Quantification of starch in plant tissues. Nature Protocols. 1 (3), 1342-1345 (2006).
- Eliceiri, K. W., et al.
- Rodrigo, J., Herrero, M. Influence of intraovular reserves on ovule fate in apricot (Prunus armeniaca L.). Sexual Plant Reproduction. 11, 86-93 (1998).
- Zhou, J., Spallek, T., Faulkner, C., Robatzek, S. CalloseMeasurer: A novel software solution to measure callose deposition and recognise spreading callose patterns. Plant Methods. 8 (1), (2012).
- Faulkner, C., et al. An automated quantitative image analysis tool for the identification of microtubule patterns in plants. Traffic. 18 (10), 683-693 (2017).
- Kuhn, B. F. Determination of starch in ovules of the sour cherry cv. "Stevnsbaer.". European Journal of Horticultural Science. 71 (3), 120-124 (2006).
- Johansen, D. A. Plant microtechnique. , McGraw-Hill. New York, NY. (1940).
- Ruzin, S. E. Plant microtechnique and microscopy. , Oxford University Press. New York, NY. (1999).
- Rodrigo, J., Rivas, E., Herrero, M. Starch determination in plant tissues using a computerized image analysis system. Physiologia Plantarum. 99 (1), 105-110 (1997).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Ovary starch reserves and pistil development in avocado (Persea americana). Physiologia Plantarum. 140 (4), 395-404 (2010).
- Suarez, C., Castro, A. J., Rapoport, H. F., Rodriguez-García, M. I. Morphological, histological and ultrastructural changes in the olive pistil during flowering. Sexual Plant Reproduction. 25, 133-146 (2012).
- Lang, G. A., Early, J. D., Martin, G. C., Darnell, R. L. Endodormancy, paradormancy, and ecodormancy - Physiological terminology and classification for dormancy research. HortScience. 22 (3), 371-377 (1987).
- Hedhly, A., Vogler, H., Eichenberger, C., Grossniklaus, U. Whole-mount clearing and staining of arabidopsis flower organs and siliques. Journal of Visualized Experiments. 2018 (134), 1-10 (2018).
- Kaufmann, H., Blanke, M. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. Journal of Plant Physiology. 218 (July), 1-5 (2017).
- Herrero, M., Dickinson, H. G. Pollen-pistil incompatibility in Petunia hybrida: changes in the pistil following compatible and incompatible intraspecific crosses. Journal of Cell Science. 36, 1-18 (1979).
- Carpenter, A. E., et al. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biology. 7 (10), R100 (2006).
