

Summary
우리는 조직화 학적인 기술을 결합 하는 이미지 분석 시스템을 사용 하 여 겨울 휴면 중 달콤한 체리 (벚나무 avium L.)에서 난소 primordia에서 녹말 함유량을 계량 하는 방법론을 제시.
Abstract
작은 구조에서 전 분에 변경 수정 하 열매를 맺은의 발병 수 분에서 생식 단계를 포함 하 여 여러 식물 발달 과정, 동안 주요 이벤트와 연결 되어 있습니다. 그러나, 꽃 감 별 법 동안에 전 분 유사 알려져 있지 않습니다 완전히, 꽃 primordia의 특히 작은 구조에서 녹말 함유량 측정의 어려움 때문에 주로. 여기, 우리는 다른 단계와 녹말 함유량에 있는 변화를 관련 된 수 있는 현미경에 부착 된 이미지 분석 시스템을 사용 하 여 달콤한 체리 (벚나무 avium L.)의 난소 primordia에서 전 분의 정량화 하는 방법 설명 봄가을 휴면. 이 위해 꽃 봉 오리의 휴면 상태 제어 조건에 겨울 시간에 다른 순간에 전송 하는 촬영의 새싹 성장 평가 하 여 결정 됩니다. 난소 primordia에서 전 분의 정량화에 대 한 꽃 봉 오리는 순차적으로 수집, 고정, 파라핀 왁 스에 포함 된, sectioned, 그리고 난 물2Kl (요오드 화 칼륨 요오드). 준비는 현미경 관찰 하 고 명확 하 게 배경에서 전 분을 구분 하는 이미지 분석기에 의해 분석. 전 분 콘텐츠 값 공부 프레임의 녹말 함유량의 추정으로 각 픽셀의 광학 밀도의 합계를 고려 하 고 묻은 전 분에 해당 하는 이미지의 광학 밀도 측정 하 여 얻을 수 있습니다.
Introduction
온화한 우 디 사계절 그들의 성장과 발전을 변조 하 여 계절에 적응. 그들은 봄, 여름 동안 개발 하는 동안 그들은 휴면 겨울1에 서가 동안 성장 하 고 중지 합니다. 휴면 낮은 겨울 온도에서 살아남을 수 있습니다, 재미는 적절 한 budburst 봄2에 대 한 전제 조건. 온대 과일 생산 및 임업에 휴면의 중요 한 의미를 확인 하 고 예측 하는 휴면 기간3다양 한 노력을 끌고있다. 과일 나무 종에 경험적 실험 조건 및 꽃의 데이터를 기반으로 하는 통계 예측 강제 촬영 전송 가지 현재 추정 하는 연구를 수 있는 휴면의 날짜를 결정 하는 각 품종에 대 한 요구 사항을 재미 그러나, 생물 학적 과정에 따라 휴면 상태를 확인 하는 방법 불분명3남아 있습니다.
달콤한 체리 (벚나무 avium L.), 등 온대 과일 나무에 꽃이 피는 일년에 한 번 발생 하 고 보름 정도 지속. 그러나, 꽃을 차별화 하 고 이전 여름4기간 동안 약 10 개월 이전에 개발 시작. 꽃 primordia 겨울 동안 휴면 새싹 안에 남아가 동안 성장 하 고 중지 합니다. 이 기간에 각 품종 축적 적절 한 꽃4에 대 한 특정 재미 요구 해야 합니다. 겨울 동안 싹에서 phenological 변화의 부족에도 불구 하 고 꽃 primordia 순수 활성화 휴면, 동안 되며 온도 재미의 축적 최근 전 분 축적의 동적으로 연결 하거나 감소 난소 primordium의 세포 내에서 휴면 결정5에 대 한 새로운 접근을 제공합니다. 그러나, 작은 크기와 난소 primordium의 위치는 특별 한 방법론을 필요합니다.
전 분 우 디 식물 종6주요 저장 탄수화물 이다. 따라서, 변경 전 분에서 꽃 조직, 그들의 개발7,8을 지원 하기 위해 탄수화물을 필요의 생리 적 활동에 관련 되어있다. 생식 과정에서 다른 키 이벤트 또한 녹말 함유량 다른 꽃 구조, 꽃 밥 감수 분열 등9, 스타일이 나 난 수정10통해 꽃가루 관의 성장에에서 있는 변이 관련 됩니다. 조직화 학적인 기법 휴면 중 꽃 primordia의 각 특정 조직에 녹말의 검출을 허용합니다. 그러나, 어려움은 조직, 재배, 또는 년 중 콘텐츠 시간 또는 비교 전 분 축적/감소의 패턴에 따라 허용 하는 전 분을 측정에 남아 있습니다. 이것은 작은 양의 분석 기법11에 사용할 수 있는 조직입니다. 대신, 이미지 분석 현미경12 에 연결 된 식물 조직13의 아주 작은 샘플에서 전 분의 정량화를 수 있습니다.
현미경 및 이미지 분석을 결합 하는 접근 계량 식물 조직, callose14,15, microtubes 등 다른 부품의 콘텐츠 또는 특정 하 여 염색 하는 영역의 크기를 측정 하 여16, 전 분을 사용합니다 얼룩. 전 분, 대 한 그것은 검출 될 수 있다 쉽게 요오드 화 칼륨 요오드를 사용 하 여 (나2기) 반응17. 이 메서드는 매우 구체적인; 내가2기 내에서 전 분 곡물의 구조는 intercalates 및 전 분18amylose 내용에 따라 어두운 파란색 또는 붉은 갈색 색상, 형태. 섹션 I로 얼룩진2기 얼룩이 보여 전 분 및 이미지 분석 시스템19애매 녹말 검출 및 후속 정량화를 허용 하는 배경 조직 사이의 적절 한 대조. 이 염료는 화학 량 론, 요오드의 축적은 매우17을 변화할 수 있는 전 분 분자의 길이에 비례 합니다. 따라서, 픽셀 수로 표현 된 스테인드 영역의 크기 수 있습니다 반영 하지 정확 하 게 전 분의 내용 때문에 비슷한 크기의 스테인드 영역 필드 사이 녹말 함유량에서 높은 차이 찾을 수 없습니다. 대신, 녹말 함유량 살구8,13 에 다른 조직에 보고 되는 현미경에서 얻은 흑백 이미지에 묻은 알갱이의 광학 밀도 측정 하 여 평가 될 수 있습니다. , 19,20, 아보카도10,그리고 올리브21.
여기, 우리가 설명 달콤한 체리에 봄가 난소 primordium 조직에 녹말 함유량의 정량화와 휴면 상태의 실험 결정을 결합 하는 방법 이해 및 예측을 위한 새로운 도구를 제공 하 고 생물 학적 메커니즘의 연구에 따라 휴면 휴면 연결.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 휴면 결정과 식물 자료를 수집
- 예제 필드에 꽃 봉 오리. 휴면 연구 장기 실험 하 고 필요 성인 싹 수집 하 큰 만큼 나무와 나무의 개발 다음 봄 동안 손상 없이 모든 겨울을 쏘고. 특별 한 과수원 관리 훈련 시스템;에 따라 필요한 수 있습니다. 따라서, 가지 치기 과일 생산 목적 보다 덜 심각한 수 있습니다.
- 새싹 틈의 발병까지가 시작에서 매주 수집 하 고 10 꽃 봉 오리를 무게.
- 모자와 함께 10 mL 유리 튜브에 그들을 배치 하 여 꽃 봉 오리를 수정 하 고 샘플 4 ° c.에 24 h 이상 에탄올/아세트산 산 (3:1)의 통 솔루션에 흠뻑 젖어 그리고는 정착 액을 버리고 75% 에탄올, 넉넉한 샘플 포함 보장을 추가 합니다. 샘플 사용까지 4 ° C에서이 솔루션에 보존 될 수 있습니다.
참고: 꽃 봉 오리 안에 기포를 제거 하 고 부동에서 그들을 방지 하는 진공 침투를 사용할 수 있습니다. 이 조직에는 정착 액의 침투를 용이 하 게 하지만 조직의 구조를 손상 될 수 있습니다. 필요 하지 않은 경우 그것을 방지 하려고 합니다.
- 필드에서 촬영 샘플.
- 각 주, 버드 휴식의 발병까지가 시작에서 15-30 cm 길이 직경, 10 꽃 싹을 포함 된 5 m m의 3 개의 촬영을 가져가 라. 22 ± 1 ° C 12 h 빛 photoperiod에 성장 챔버에 물에 젖은 꽃 거품에 넣어.
- 10 일 후에 성장 챔버, 선택 하 고 무게는 촬영에서 10 꽃 봉 오리.
- 새싹 성장 평가 하 고 휴면 상태를 결정 합니다. 샘플링 기간 위치 조건에 조정 될 수 있다. 과수원 조건에서 (사라고사, 스페인, 41 ° 44'30 "N, 0 ° 47'00" W, 그리고 해발 220 m), 쏘고의 샘플링에서 11 월 30 일 2 월 말 또는 3 월 초까지 실시 되었다.
- 주간 평가 꽃 봉 오리의 응답 챔버의 적당 한 성장 조건까지 budburst 봄,가 시작 부분에서 필드에서 고른 10 싹의 무게를 비교 하 여.
- 차이가 없거나 그 차이 30% 미만, 새싹 재미 그들의 요구를 실현 하지가지고 고 endodormant22는 고려 하십시오. 차이가 있다면 30% 이상, 새싹 재미 그들의 요구를 충족 하 고 ecodormant22는 고려 하십시오.
2. 식물 재료 준비 전 분 정량화에 대 한
- 각 샘플링에서 선택에 대해 6 개의 고정된 싹 날짜 (단계 1.1 참조). 재배, 따라 각 달콤한 체리 꽃 봉 오리에는 최대 5 개의 꽃 primordia를 포함 되어 있습니다.
- 외부 버드 비늘을 제거 하 고 밖으로 건조에서 그것을 방지 하기 위해 75% 에탄올과는 시계 유리에 꽃 봉 오리를 놓습니다.
- 꽃 봉 오리를 해 부하 고 정밀 집게의 도움과 입체 현미경 안과 메스 새싹 당 적어도 하나의 꽃 primordium 추출 합니다. 꽃 primordium 4 ° C에 75% 에탄올 10 mL 유리 튜브에 보존 될 수 있습니다 또는 다음 단계로 바로 진행 합니다.
- 제 3 부 틸 알코올 시리즈에서 샘플 탈수.
- 75 %3 차 부 틸 알콜 아낌없이 샘플을 커버 하 고 1.5 h에 대 한 그들을 품 어 (미정)의 10 mL에 75% 에탄올 솔루션을 바꿉니다. 파스퇴르 피 펫 솔루션 삭제 하도록 유용할 수 있습니다.
- 이후 순수 미정 실내 온도 (< 20 ° C)에서 crystalizes와 매우 휘발성 이며 독성 공기 추출, 30 ° C에서 건조 난로에 단계 2.2.1, 상승 농도 (85%, 95%, 및 100 %v / v, 및 순수한 미정 3 x)와 함께 TBA를 추가 반복 합니다. 미정 샘플 crystalizes, 그것은 조직 손상 것입니다.
- 파라핀18에 샘플을 포함 합니다.
- 공기 추출 전날 60 ° C에서 건조 하는 난로 있는 파라핀 진주를 도입 하 여는 파라핀을 녹여. 파라핀 진주 또한, 열판에 녹아 수 있습니다 하지만 액체 파라핀 60 ° C에 그리고 조직 손상을 방지 하기 위해 높은 온도에 아닙니다 확인 하십시오.
- 미정 미정과 파라핀 오일 (1:1)의 혼합으로 대체 하 고 60 ° c.에 건조 난로 내부 24 h에 대 한 품 어 그런 다음, 미정의 믹스를 대체 하 고 순수한 파라핀 오일 파라핀을 녹여 및 60 ° c.에 건조 난로 안에 적어도 6 h에 대 한 품 어 반복 2 x 적어도 4-6 d에 대 한 마지막 변경 품 어 고.
- 열 표면에 작은 기본 금형에 각 샘플 배치 포함 카세트 속도 파라핀 왁 스에 포함 된. 차가운 표면 위에 그것을 배치 하 고 왁 스가 고형화 되 면 블록을 제거 합니다.
- 섹션 하 고 준비를 rehydrate.
- 준비 하는 Haupt의 접착제: 30 ° C에 증류수 100 mL에 일반 녹 스 젤라틴의 1 g을 분해 다음, 2 세대의 페 놀 느슨한 (C6H5오)와 글리세롤의 15 mL를 추가 합니다. 브러시와 유리 슬라이드에 걸쳐 Haupt의 접착제 하 고 1% 포름알데히드 용액 한 방울을 추가 합니다.
- 섹션 각 파라핀와 톰에 10 µ m에서 차단 하 고 유리 슬라이드에 섹션을 배치 Haupt의 접착제로 덮여.
- 건조;까지 단면도 유리 슬라이드를 35-40 ° C에서 열 표면에 놓으십시오 그런 다음 유리 슬라이드 슬라이드 랙 이동 합니다.
- Dewax 및 리하이드레이션 솔루션을 준비 합니다. 3 유리 얼룩 요리, Histoclear II:ethanol (1:1, v/v)로 또 하나, 에탄올 시리즈 Histoclear II의 200 mL를 배치 (100%, 70%, 40%, v/v), 그리고 최종 증 류 물 세척.
- 3 세척, 각 5 분, Histoclear II 및 Histoclear II:ethanol (1:1, v/v)와 섹션 dewax 슬라이드 랙 보장 솔루션 완전히 슬라이드, 커버 유리 얼룩 요리를 내 놓고, 다음, 다음을 하나의 솔루션에서 슬라이드 랙 이동 합니다.
- 2 민 세척의 다음 시리즈에 의해 섹션을 rehydrate: 에탄올 시리즈 (100%, 70%, 40%, v/v) 및 최종 증 류 물 세척. 마지막으로, 실내 온도에 유리 슬라이드 건조.
- 섹션18얼룩.
- 나에 게2기 얼룩을 준비: 요오드 화 칼륨 (KI), 요오드의 0.2 g 2 세대 분해 (나2) 100 mL의 증류수에. 신선한 방울 적용 나2기 5 분에 대 한 섹션을 통해, 다음, 얼룩의 과잉 압 지와 그것을 흡수 하 여 삭제 하 고. 신속 하 게 다음 단계를 진행 합니다.
- 합성 설치 미디어의 작은 방울을 적용, 작은 커버 유리 상단에 놓고 하드를 누릅니다. 일단 장착 미디어 건조, 난소 전 분의 예비 평가 대 한 밝은 분야 현미경으로 관찰 합니다.
참고:이 단계는 비록 전 분와 배경 사이의 더 나은 대조를 얻은 없는 필수. 섹션 전 분 정량화 후 다른 얼룩 다시 사용할 경우, 커버 유리 위에 얼룩의 드롭 놓고 압 지와 그것을 흡수 하 여 과잉을 삭제 (2.5.1 참조). 다음, 전 분 정량화, 후 씻어는 밖으로2기는 증류수와 장소는 열 이상의 준비 건조까지 35-40 ° C에 표면 얼룩.
3입니다. 녹말 함유량의 정량화
- 보정 광학 조건.
참고: 이미지 분석기 스테인드 녹말을 검출 하는 데 사용 검출 레벨은 직접 현미경;의 확대 및 조명 조건에 따라 따라서, 이러한 조건을 평가 하는 모든 준비에 대 한 수정. 조정 가능한 현미경 및 조명 조건에 여기 제안 된 적응.- 확대 및 밝기 제어 또는 빛의 강도 X 20에 조리개 다이어 프 램을 조정 합니다.
- 필터 홀더에 필터가 되어 있는지 확인 하 고 현미경에서 밝은 필드 조건을 선택 합니다. (예를 들어, 40 X 달콤한 체리 난소 primordium) 적합 한 확대를 선택 합니다.
- 이미지 수집 조건을 제어 합니다. 조직을 통해 이미지 없이 스테인드 준비 카메라 설정을 조정 | 수집 | 사전 보기.
- 50%, 1.0에서 이득에 밝기를 수정 x, 그리고 감마 1.00 및 0-명암 값 히스토그램 지표 100, 밝기 분포 히스토그램의 한계와.
- 과도/노출 부족 기능을 활성화 하 고 노출 과도의 한계에 노출 시간을 조정 합니다.
- 화이트 밸런스 기능 수정 및 균질 이미지를 만드는 완전 한 이미지를 어떤 색조와 음영 보정 없이 이미지의 모든 중립 착 색 구성 요소를 표시 하려면 전체 이미지에 적용 됩니다.
- 투과율 (T) 값으로 얻을 수 있습니다 다른 광학 밀도 (OD)의 회색 레벨 (0, 검정, 255, 흰색)의 컨트롤 값을 가져올 이미지 분석 시스템 보정.
- 제어, 간주 조직 없이 스테인드 준비의 이미지를 수집 하 고는 흑인과 백인 이미지를 통해 측정의 회색 수준을 측정 | 회색 측정 | 회색 보정 | 참조 값 = 0 | 측정 | 보정 | 확인. 이 100% 투과율;과 해당 따라서, 0 세에 따라 광학 밀도 = 2-티 로그
- 4 빛의 양을 줄여 4N 필터와 동일한 준비의 이미지를 취득, 및 측정은 흑인과 백인 이미지를 통해 측정의 회색 수준 | 회색 측정 | 회색 보정 | 참조 값 0.6 = | 측정 | 보정 | 확인. 이 25% 투과율;과 해당 따라서, 외경에 따라 0.6의 광학 밀도 = 2-티 로그
- 빛, 고려 검정 없이 동일한 준비의 이미지를 취득 하 고는 흑인과 백인 이미지를 통해 측정의 회색 레벨을 측정 | 회색 측정 | 회색 보정 | 참조 값 = 1 | 측정 | 보정 | 확인. 이 0% 투과율;과 해당 따라서, 외경에 따라 1의 광학 밀도 = 2-티 로그
- 전 분을 감지 합니다.
- 적어도 300 dpi (인치당 도트 수) 해상도와 TIFF 형식에서 측정 필드의 컬러 이미지를 취득 합니다.
- 스테인드 지역에 해당 하는 이진 이미지를 만듭니다. 세 가지 색 임계값 설정 (값 0-사이 각 255) 바이너리 이미지 정확 하 게 반영 하는 관찰을 통해 이미지 묻은 전 분 알갱이 | 검색 | 빨강, 파랑 및 녹색의 임계값을 선택 | 확인. 다양 한 준비와 조직 최종 감지 수준을 조정 하에서 반복적으로 시각적 비교를 확인 합니다. 저장 하 고 모든 준비가이 수준을 사용 합니다.
- 전 분을 계량. 이미지 분석 시스템으로 흑백 이미지에 원본 컬러 이미지를 변환 합니다. 이진 이미지는 흑백 이미지를 통해 이미지에 겹쳐 마스크로 사용 하 여 | 바이너리 편집. 모든 픽셀은 마스크 를 통해 측정에서의 광학 밀도의 합을 측정 | 회색 수준 | 확인 측정된 분야에 전 분 내용으로이 값을 고려 하십시오.
- 반복 단계 3.4.1-꽃 primordia의 난소에서 녹말 함유량의 대표 값을 가져오는 난소 primordia의 3.4.3 4 장소.
- 3.4.3 3.4.1-단계를 반복 하 고 각 수집 날짜의 다른 꽃에 3.4.5 단계.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
휴면 연구 재미 요구 사항이 충족 되는 때 순간의 결정을 해야 합니다. 필드 조건 (그림 1A)에서 겨울 동안 phenological 변화의 부족에도 불구 하 고 벚꽃 복구할 수 없습니다 적당 한 조건에서 성장의 용량 낮은 온도에서 일정 기간을 통과할 때까지. 겨울 시간 동안 제어 조건 약 실 (그림 1B)에 촬영의 정기 전이 꽃 봉 오리의 휴면 상태의 평가 허용. 꽃 봉 오리 성장의 평가 버드 체중 증가 측정 하 여 이루어졌다. 반면, 휴면, 동안 변경 관찰 될 수 있었다 적당 한 조건 (그림 1C)의 10 일 후 일단 휴면 극복, 새싹 늘어났다 고 성장 챔버 (그림 1D)에 버스트. 이 분석의 결과 허용을 싹의 휴면 상태. 겨울 동안 발생 하는 다른 온도 때문 휴면 올해에 따라 다른 날짜에 극복 했다. 반면, 연구의 1 년 동안 휴면의 1 월에 발생, 2 년 제시 온화한 겨울; 따라서, 재미 행 약 3 주 후, 2 월에 발생 했습니다.
달콤한 체리 박차, 어디 꼭대기 새싹 식물 새싹 이며 옆 꽃 봉 오리는 꽃 봉 오리 (그림 1C 및 1d)에 꽃 봉 오리를 맺는 다. Undifferentiated 싹 여름의 끝에 꽃 이나 식물 새싹으로 분화 하기 시작 하 고 그들은 겨울에 휴면을 입력, 여러 꽃 primordia 꽃 봉 오리, 수많은 녹색 비늘에 의해 보호 되 고 갈색 외부 비늘 (적용 안에 남아 그림 1C 와 그림 2A). 꽃 봉 오리의 해 부 (그림 2A) 안에 작은 꽃 primordia 보였다. 각 꽃 봉 오리 포함 1-5 개별 꽃 primordia (그림 2B). 각 꽃 primordium의 작은 크기에도 불구 하 고 꽃의 모든 부분을 구별 하는 하 고 구분할 수 있습니다:는 암 꽃 술는 anthers, 꽃잎 및 sepals (그림 2C). 조직화 학적인 방법 사용 (알코올: 아세트산 [3:1] 기정, 파라핀 왁 스를 포함, 톰 단면, 그리고 녹말 요오드 기반 얼룩) 허용 꽃 내 전 분의 분포 (그림 2D관찰 primordium 조직 ).
난소 primordium에 전 각 섹션에서 정량 했다. 1337 µ m2, 40 x 확대, 4 개의 측정 달콤한 체리 난소 primordium (그림 3A)에 있는 전 분의 일반적인 레이아웃을 표현. 전 분 알갱이 했다 후 배경에서 명확 하 게 고유2기 얼룩 (그림 3B). 전 분은 빨간색, 녹색 및 파랑의 임계값 정의 된 색상 매개 변수 (그림 3C)에 따라 시스템에 의해 만들어진 바이너리 이미지에 의해 덮여 있었다 모든 전 분 알갱이 관찰 될 때까지 색상을 조정 하 여 이미지 분석 시스템에 의해 확인 되었다. 얻은 녹말 함유량의 값 (그림 3D) 흑백 이미지에 마스크 아래 모든 픽셀의 광학 밀도의 측정의 결과 이었다.
전 분의 정량화 겨울 (그림 4) 동안 동적 전 분의 일관 된 패턴을 공개 했다. 최대 두 년에 120, 000와 140000 사이 값을 도달 하는 동안 일관 되 게, 이른 겨울에 있는 전 분의 금액 미만 400003의 광학 밀도 값 제시. 최대 값 (그림 4A) 1 년 동안 1 월에서 도달 되었다, 그러나 그것은 2 년 (그림 4B) 2 월에 발생 했습니다. 휴면의 안 색이이 결과 대조, 전 분의 최대 금액 발생 수반 두 년에 놀 아 요 행.
이 방법은 프레임 휴면 관련 전 분 내용에 변경 하기 위해서는 난소 조직 (그림 5B)에 전 분 정량화와 수반 휴면 상태 (그림 5A)의 결정이 필요 합니다.

그림 1: 달콤한 벚꽃의 꽃 봉 오리의 휴면 상태의 결정에 대 한 실험 설정. (A) 분 지, 겨울 동안 휴면 싹 폐쇄 하 고 어두운 갈색 비늘으로 덮여 표시. (B)이이 패널 성장 챔버로 촬영을 보여줍니다. 1 월 중순에 일부 cultivars 싹 휴면 남아 여전히 폐쇄 (흰색 화살표), 다른 성장, 수 있었다 하는 동안 치명적된 싹 (검은색 화살표)를 보여주는. (C)이이 패널 옆에 있는 휴지 꽃 봉 오리와 단일 식물 새싹 박차 꼭대기 위치에 촬영의 세부 정보를 보여줍니다. (D)이이 패널 보여줍니다 촬영의 세부 사항을 일단 휴면 성장 챔버에 10 일 후에 극복 했다 새싹 붓기를 보여주는. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
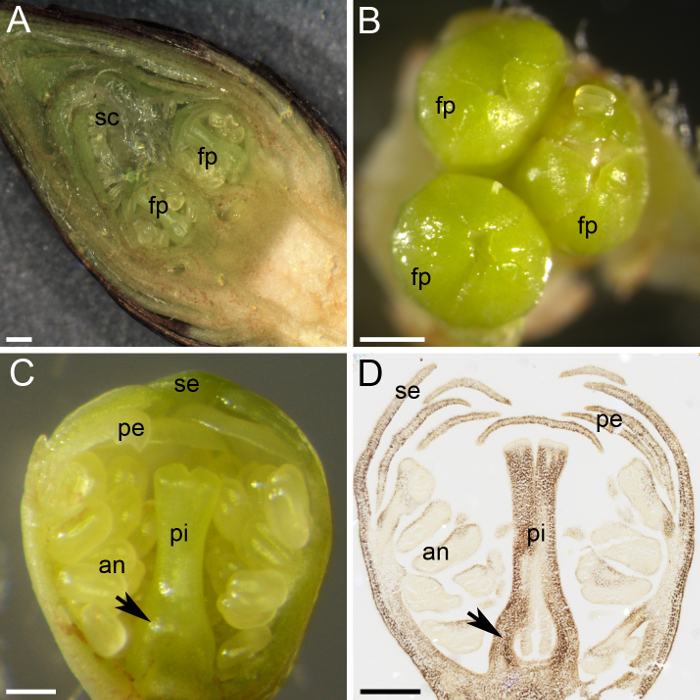
그림 2: 식물의 전 분 정량화에 대 한 재료 준비. (A)이이 패널 표시 횡단 부분의 꽃 봉 오리, 두 꽃 primordia (fp)를 보여주는 수많은 비늘 (sc)에 의해 보호 됩니다. (B) 3 꽃 primordia (fp)는 꽃 봉 오리에 모입니다. (C)이이 패널 표시 꽃 primordium 횡단 부분의 모든 whorls와 분화: sepals (se), 꽃잎 (pe), anthers (한), 그리고 암 꽃 술 (pi). 난소 primordium 암 꽃 술 (화살표)의 기지에서 수훈이 있다. (D) A 중간 부분의 꽃 primordium 수집 및 1 월에 고정, 파라핀 왁 스에 포함 된, 구분, 되었고 나 물 전 분을 위한2기 (짙은 갈색). 이 패널에서는 난소 primordium (화살표)를 보여 줍니다. 스케일 바 패널 A 와 B 에 500 µ m 및 패널 C 와 D의 200 µ m입니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 3: 달콤한 체리 난소 primordium 부 량 전 분. (A)이이 패널 난소 primordium의 중간 부분을 보여줍니다 난 물2기, 콘텐츠는 전 분에 4 개의 프레임을 보여주는 측정 했다. (B)이이 패널 난소 primordium의 세부 사항을 표시합니다. 전 분 알갱이 짙은 갈색에 물 들 다. (C)이이 패널 표시는 전 분에 모조 색상 이미지에 파랑의 다른 그늘에 해당 합니다. (D)이이 패널 흑백 원본 이미지에 (파란색) 나2기 묻은 전 분을 취재 하는 바이너리 이미지 마스크를 보여 줍니다. 광학 밀도 마스크에 의해 포함 하는 원본 이미지의 픽셀에만 측정 됩니다. 스케일 바 패널 A 에서 100 µ m와 20 µ m B - D패널에 있습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.

그림 4: 달콤한 체리 난소 primordia에서 전 분 정량화의 대표적인 결과 다른 겨울 온도 조건의 2 년 동안 봄가 매월 수집. (A)이이 패널 표시 결과 2010-2011 년에서 추운 겨울을 했다 합니다. 놀 아 요 행 (눈송이) 1 월, 전 분의 최대 크기와 수반에 발생 했습니다. (B)이이 패널 했다 온화한 겨울 2011-2012 년에서 결과 표시 합니다. 놀 아 요 행 (눈송이) 2 월, 전 분의 최대 크기와 수반에 발생 했습니다. 값은 평균 ± 표준 오차의 의미. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
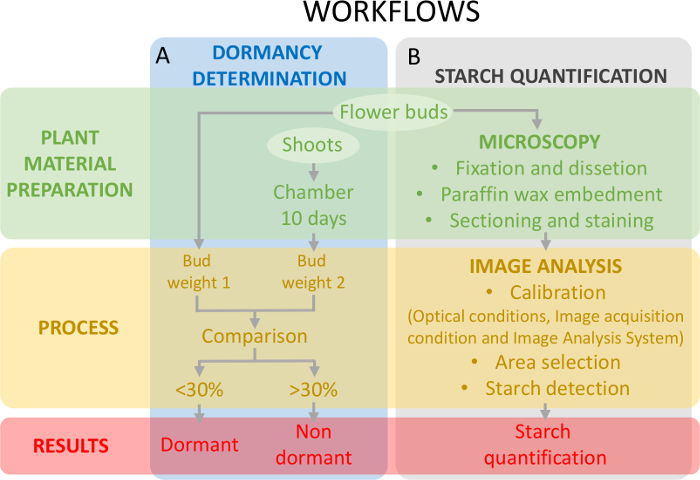
그림 5: 새싹과 달콤한 체리에서 난소 primordia에서 전 분 정량화의 휴면 상태를 평가 하기 위해 실험 디자인의 계획. (A)이이 패널의 휴면 상태 결정의 워크플로 보여 줍니다: 식물 소재 준비, 과정, 그리고 얻은 결과. (B)이이 패널 전 분 정량화의 워크플로 보여 줍니다: 전 분, 전 분의 이미지 분석 탐지 및 전 분 정량화의 현미경 관찰을 위한 새싹의 조직화 학적인 준비. 이 그림의 더 큰 버전을 보려면 여기를 클릭 하십시오.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
우 디 사계절에 휴면 휴면 뒤에 생물 학적 과정 불분명 남아 있지만 과일 생산에서 명확한 의미와 변화 하는 기후에 있는 임업을 선물 한다. 휴면 연구는 다른 관점에서 접근 될 수 있다 하지만 겨울 휴면에 대 한 생물 학적 마커를 찾는 연구는 지난 년 동안을 강화 하고있다. 그러나 때 새싹 휴면 고장이 보여주는 명확한 지표를 찾을 대부분의 시도 실패3되었습니다. 분석 이미지, 조직화 학적인 기법18 결합 여기에서 설명한 방법론 특정 조직의 탄수화물 보유와 달콤한 체리 새싹에서 그것의 생리 적인 활동의 관계를 검토 하 매우 유용 했습니다. 휴면5 의 여러 단계와 다른 종 및 조직10,,1320에 적용할 수 있습니다.
전 분의 형태로 탄수화물 보유 재생 모두 생식 과정7,10,13,20,,2123 꽃 개발에 중요 한 역할을 하 고 온대 우 디 사계절6의 계절. 휴면에 대 한 다른 연구는 싹24내에서 전 분에 관심을 지불 했습니다. 그러나, 그들의 작은 크기로 인해 여러 전체 싹 양적 분석 방법의 사용에 필요한 이며 특정 조직이 나 세포의 양적 변화 감지 주변 세포의 마스킹 효과 의해 제한 됩니다. 이미지 분석 시스템으로 조직화 학적인 기술 조합 꽃 봉 오리 안에 다른 구조의 녹말 함유량에 변화를 공부 하는 좋은 기회를 제공 합니다.
이 방법은 조직에 있는 전 분의 정확한 콘텐츠의 정량화를 방지의 제한이 하지만을 사용 하면은 상대적인 전 분 시간25 이상 전 분 양적 변화에 따라 전 분을 비교 하 고 측정할 수의 콘텐츠 다른 조직13,20, 재배, 또는 년5. 분야, 조직, 및 싹 사이 광학 밀도의 값, 시스템 (조명 조건, 얼룩, 강도와 확대)의 교정 및 색상 임계값을 정확 하 게 설정 해야 합니다 설정의 정확한 비교를 허용 하기 위하여 저장, 및 모든 준비에 사용.
전 분 얼룩이 지 고 칼륨 요오드에 따라 정량화 섹션의 세척 후 다른 얼룩의 후속 사용을 수 있습니다. 따라서, 추가 준비 없이 다른 분석을 수행할 수 있습니다 및5,20을 사용 하는 합성 장착 미디어 하지 않습니다. 마찬가지로, 형태학 측정26 수 있도록 서로 다른 구조5,13의 성장에 관하여 액자를 전 분 축적의 패턴 같은 준비 후 할 수 있습니다. 메서드를 사용 하면 다른 구조 또는 전 분, 녹말 함유량 셀의 작은 그룹에 변경 내용을 포함 하는 다른 개발 프로세스의 연구를 허용할 수 있다을 감지 하는 분석기를 사용 하는 색상 레벨의 조정 종 적응 될 수 있다.
관계 휴면 릴리스 사이의 전 분 축적이이 방법의 사용에 의해 난소 primordia에서 휴면 및 재미 요구 사항5의 생물 학적 기초를 이해 하 사운드 기초를 제공 한다. 그러나, 파라핀 포함 섹션에 이미지 분석에 의해 전 분 정량화 매우 복잡 하 고 재배의 많은 수의 재미 요구 사항을 예상 시간이 되도록 밖으로 돌 수 있었다. 향후 쉽게 나무의 휴면 상태를 나타낼 수 있는 신뢰할 수 있는 생물학적 지표를 공부에 집중 해야 합니다. 한편, 휴면 중 전 분 변이의 패턴 추가 생리와 유전 연구, 프레임을 사용할 수 있습니다 그리고 여기에 설명 된 이미지 분석 조직화 학적인 기술 조합 기타 우 디 다년생 작물에 사용 될 수 있습니다. 휴면에 관하여 다른 조직의 녹말 함유량 계량.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
저자는 공개 없다.
Acknowledgments
저자는 기꺼이 그들의 도움이 토론과 조언을 위한 마리아 Herrero와 Eliseo 리 바 감사합니다. 이 작품은 정부의 드 Economía y Competitividad에 의해 지원 되었다-유럽 지역 개발 기금, 유럽 연합 [부여 수 있다-2010-037992 E. f]; Instituto Nacional de Investigación y 기술과 Agraria y 섬유 [보조금 번호 RFP2015-00015-00 RTA2014-00085-00, RTA2017-00003-00]; 그리고 Gobierno de Aragón-유럽 사회 기금, 유럽 연합 [Grupo Consolidado A12 17R].
Materials
| Name | Company | Catalog Number | Comments |
| Precision scale | Sartorius | CP225D | |
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Drying-stove | Memmert | U15 | |
| Paraffin Embedding station | Leica Microsystems | EG1140H | |
| Rotatory microtome | Reichert-Jung | 1130/Biocut | |
| Microtome blade | Feather | S35 | Stainless steel |
| Bright field microscope | Leica Microsystems | DM2500 | |
| Digital Camera | Leica Microsystems | DC-300 | |
| Image Analysis System | Leica Microsystems | Quantiment Q550 |
References
- Kurokura, T., Mimida, N., Battey, N. H., Hytönen, T. The regulation of seasonal flowering in the Rosaceae. Journal of Experimental Botany. 64 (14), 4131-4141 (2013).
- Rohde, A., Bhalerao, R. P.
- Fadón, E., Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environmental and Experimental Botany. 152, 28-36 (2018).
- Fadón, E., Rodrigo, J., Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees - Structure and Function. , In Press (2018).
- Fadón, E., Herrero, M., Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Frontiers in Plant Science. 9 (171), (2018).
- Loescher, W. H., Mccamant, T., Keller, J. D. Carbohydrate reserves, translocation and storage in woody plant roots. HortScience. 25 (3), 274-281 (1990).
- Hedhly, A., et al. Starch turnover and metabolism during flower and early embryo development. Plant Physiology. , (2016).
- Rodrigo, J., Hormaza, J. I., Herrero, M. Ovary starch reserves and flower development in apricot (Prunus armeniaca). Physiologia Plantarum. 108 (1), 35-41 (2000).
- Julian, C., Rodrigo, J., Herrero, M. Stamen development and winter dormancy in apricot (Prunus armeniaca). Annals of Botany. 108 (4), 617-625 (2011).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil starch reserves at anthesis correlate with final flower fate in avocado (Persea americana). PLoS One. 8 (10), e78467 (2013).
- Smith, A. M., Zeeman, S. C. Quantification of starch in plant tissues. Nature Protocols. 1 (3), 1342-1345 (2006).
- Eliceiri, K. W., et al.
- Rodrigo, J., Herrero, M. Influence of intraovular reserves on ovule fate in apricot (Prunus armeniaca L.). Sexual Plant Reproduction. 11, 86-93 (1998).
- Zhou, J., Spallek, T., Faulkner, C., Robatzek, S. CalloseMeasurer: A novel software solution to measure callose deposition and recognise spreading callose patterns. Plant Methods. 8 (1), (2012).
- Faulkner, C., et al. An automated quantitative image analysis tool for the identification of microtubule patterns in plants. Traffic. 18 (10), 683-693 (2017).
- Kuhn, B. F. Determination of starch in ovules of the sour cherry cv. "Stevnsbaer.". European Journal of Horticultural Science. 71 (3), 120-124 (2006).
- Johansen, D. A. Plant microtechnique. , McGraw-Hill. New York, NY. (1940).
- Ruzin, S. E. Plant microtechnique and microscopy. , Oxford University Press. New York, NY. (1999).
- Rodrigo, J., Rivas, E., Herrero, M. Starch determination in plant tissues using a computerized image analysis system. Physiologia Plantarum. 99 (1), 105-110 (1997).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Ovary starch reserves and pistil development in avocado (Persea americana). Physiologia Plantarum. 140 (4), 395-404 (2010).
- Suarez, C., Castro, A. J., Rapoport, H. F., Rodriguez-García, M. I. Morphological, histological and ultrastructural changes in the olive pistil during flowering. Sexual Plant Reproduction. 25, 133-146 (2012).
- Lang, G. A., Early, J. D., Martin, G. C., Darnell, R. L. Endodormancy, paradormancy, and ecodormancy - Physiological terminology and classification for dormancy research. HortScience. 22 (3), 371-377 (1987).
- Hedhly, A., Vogler, H., Eichenberger, C., Grossniklaus, U. Whole-mount clearing and staining of arabidopsis flower organs and siliques. Journal of Visualized Experiments. 2018 (134), 1-10 (2018).
- Kaufmann, H., Blanke, M. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. Journal of Plant Physiology. 218 (July), 1-5 (2017).
- Herrero, M., Dickinson, H. G. Pollen-pistil incompatibility in Petunia hybrida: changes in the pistil following compatible and incompatible intraspecific crosses. Journal of Cell Science. 36, 1-18 (1979).
- Carpenter, A. E., et al. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biology. 7 (10), R100 (2006).
