

Summary
Apresentamos uma metodologia para quantificar o teor de amido no primordia de ovário em doce de cereja (Prunus avium L.) durante a letargia do inverno por meio de um sistema de análise de imagem combinado com técnicas histoquímicas.
Abstract
Alterações em amido em pequenas estruturas estão associadas com eventos chaves durante vários processos de desenvolvimento da planta, incluindo a fase reprodutiva de polinização, a fecundação e o início da frutificação. No entanto, variações em amido durante a diferenciação de flor não são completamente conhecidas, principalmente devido à dificuldade de quantificar o teor de amido nas estruturas particularmente pequenos do primordia a flor. Aqui, descrevemos um método para a quantificação de amido no primordia de ovário de doce de cereja (Prunus avium L.), usando um sistema de análise de imagem anexado para o microscópio, que permite relativas as alterações no teor de amido com as diferentes fases de dormência do Outono à Primavera. Para essa finalidade, o estado de dormência dos botões florais é determinado pela avaliação o crescimento do broto de brotos transferidos para condições controladas em diferentes momentos no tempo de inverno. Para a quantificação de amido no primordia ovário, botões de flores são sequencialmente coletados, fixo, incorporados em cera de parafina, seccionados e manchados com I2Kl (iodo-iodeto de potássio). As preparações são observadas ao microscópio e analisadas por um analisador de imagem que claramente distingue o amido do fundo. Valores de conteúdo de amido são obtidos através da medição da densidade óptica da imagem que corresponde com a do amido manchado, considerando a soma da densidade óptica de cada pixel como uma estimativa do teor de amido dos quadro estudado.
Introduction
Plantas perenes lenhosas temperadas adaptar-se às estações da ano, modulando seu crescimento e desenvolvimento. Enquanto eles desenvolvem durante a primavera e o verão, eles param de crescer durante o Outono para ir dormente no inverno1. Apesar de letargia que lhes permite sobreviver a temperaturas de inverno baixa, refrigeração é um pré-requisito para uma adequada looks na primavera2. As implicações importantes de dormência na produção de frutas temperadas e florestas têm levado a diversos esforços para determinar e prever o período de dormência3. Em espécies de árvores de fruto, experimentos empíricos transferir brotos para forçar as condições e previsões estatísticas com base em dados da floração são abordagens atuais para determinar a data da quebra de dormência, o que permite que os pesquisadores estimar o relaxando os requisitos para cada cultivar. No entanto, como determinar o estado de letargia, baseado em processos biológicos permanece incerto3.
Floração em árvores de fruto de clima temperado, como doce de cereja (Prunus avium L.), ocorre uma vez por ano e dura cerca de duas semanas. No entanto, as flores começam a diferenciar e desenvolver cerca de 10 meses antes, durante o verão anterior4. Primordia flor param de crescer durante o Outono para permanecer dormente dentro os botões durante o inverno. Neste período, cada cultivar precisa acumular um determinado requisito arrepiante para adequada floração4. Apesar da falta de mudanças fenológicas nos botões durante o inverno, primordia flor é fisiologicamente ativa durante a dormência, e a acumulação de temperaturas de refrigeração foi recentemente associada com a dinâmica da acumulação de amido ou diminuir dentro das células do primórdio ovário, oferecendo uma nova abordagem para determinação de letargia5. No entanto, o pequeno tamanho e a localização do primórdio ovário requerem uma metodologia especial.
O amido é o carboidrato de armazenamento principal em espécie de planta lenhosa6. Assim, alterações em amido têm sido relacionadas à atividade fisiológica dos tecidos da flor, que precisa de carboidratos para apoiar o seu desenvolvimento7,8. Diferentes eventos-chave durante o processo reprodutivo também estão relacionados às variações no teor de amido em diferentes estruturas florais, como antera meiose9, o crescimento dos tubos de pólen através do estilo ou óvulo fecundação10. Histochemical técnicas permitem a detecção de amido em cada tecido específico do primordia flor durante a dormência. No entanto, a dificuldade permanece em quantificar esse amido para permitir que conteúdo entre os tecidos, cultivares ou anos seguindo seu padrão de acumulação/diminuir ao longo do tempo ou comparar o amido. Isto é devido à pequena quantidade de tecido disponível para as técnicas analíticas de11. Como alternativa, análise de imagem ligada à microscopia12 permite a quantificação do amido em muito pequenas amostras de tecido de planta13.
Abordagens combinando análise de microscopia e imagem têm sido utilizadas para quantificar o conteúdo dos diferentes componentes nos tecidos da planta, tais como callose14, microtubos15, ou16, medindo-se o tamanho da área tingida por específicos de amido manchas. Para o amido, pode ser facilmente detectada usando o iodo-iodeto de potássio (eu2KI) reação17. Este método é altamente específico; Eu2KI intercala dentro da estrutura laminar de grãos de amido e dá forma a uma cor azul escuro ou marrom-avermelhada, dependendo do conteúdo de amilose do amido18. Seções manchadas com I2mancha KI mostrar o contraste adequado entre o amido e o tecido de fundo, permitindo uma deteção de amido inequívoca e posterior quantificação pelo de sistema de análise de imagem19. Embora este corante não é estequiométrica, o acúmulo de iodo é proporcional ao comprimento da molécula de amido, que altamente pode variar de17. Assim, o tamanho da área manchada, expressado como o número de pixels pode não refletir com precisão o teor de amido, desde altas diferenças no teor de amido podem ser encontradas entre os campos com áreas manchadas de tamanho similar. Como alternativa, o teor de amido pode ser avaliado através da medição da densidade óptica dos grânulos corados em preto e brancos imagens obtidas com o microscópio, como tem sido relatado em diferentes tecidos em Damasco8,13 , 19, abacate10,20e21de azeitona.
Aqui, descrevemos uma metodologia que combina a determinação experimental do estatuto de dormência com a quantificação do teor de amido no tecido do ovário Primórdio de Outono à Primavera em doce de cereja, oferecendo uma nova ferramenta para a compreensão e predição de dormência, baseada no estudo dos mecanismos biológicos ligados com letargia.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. dormência determinação e planta coleta de Material
- Os botões da flor no campo da amostra. Estudos de dormência são experiências a longo prazo e requerem adulto árvores grande o suficiente para coletar os botões e dispara todo o inverno sem comprometer o desenvolvimento das árvores durante a próxima primavera. Gestão de pomar especial pode ser necessária, dependendo do sistema de formação; assim, a poda pode ser menos grave do que para fins de produção de frutas.
- Cada semana, desde o início do outono até o início do abrolhamento, recolher e pesar 10 botões de flores.
- Corrigir os botões da flor, colocando-os num tubo de vidro de 10 mL com tampa e mergulhe as amostras em uma fixador solução de ácido acético/etanol (1:3) pelo menos 24 h a 4 ° C. Em seguida, descartar o fixador e adicionar etanol 75%, generosamente, garantindo que cobre as amostras. As amostras podem ser conservadas nesta solução a 4 ° C até o uso.
Nota: A infiltração de vácuo pode ser usada para remover as bolhas de ar dentro do broto e impedi-los de flutuante. Isso facilita a penetração do fixador para os tecidos, mas pode danificar a estrutura dos tecidos. Tente evitá-lo, se não for necessário.
- Os brotos no campo da amostra.
- Cada semana, desde o início do outono até o início do abrolhamento, leva três tiros de 15-30 cm de comprimento e 5 mm de diâmetro, contendo pelo menos 10 botões florais. Coloque-os na espuma de florista encharcada de água em uma câmara de crescimento a 22 ± 1 ° C, com um fotoperíodo de 12 h luz.
- Após 10 dias em câmara de crescimento, escolha e pesar 10 botões florais de brotos.
- Avaliar o crescimento do broto e determinar o estado de letargia. O período de amostragem deve ser ajustado para as condições do local. Nas condições de pomar (Zaragoza, Espanha, 41 ° 44'30 "N, 0 ° 47'00" W e 220 m acima do nível do mar), a amostragem da brotos foi realizada de 30 de novembro até o final de fevereiro ou início de março.
- Semanalmente avalie a resposta dos botões da flor para as condições de crescimento adequado da câmara, desde o início do outono até looks na primavera, comparando o peso dos 10 brotos colhidos a partir do campo.
- Se não existem diferenças ou essas diferenças são menos de 30%, consideram que os gomos não tem preenchido os requisitos de refrigeração e são endodormant22. Se as diferenças são mais de 30%, consideram que os gomos tem preenchido os requisitos de refrigeração e são ecodormant22.
2. planta de preparação Material para quantificação de amido
- Selecionados cerca de seis botões fixos de cada amostragem datam (consulte a etapa 1.1). Cultivares, dependendo de cada broto de flor de cereja doce contém até cinco primordia de flor.
- Remover as escamas externas broto e coloque o broto no vidro de um relojoeiro com 75% de etanol para evitar sequem.
- Dissecar o broto e extrair pelo menos um Primórdio de flor por broto com a ajuda de pinças de precisão e um bisturi oftalmológico sob um microscópio estereoscópico. O Primórdio de flor pode ser conservado em um tubo de vidro de 10 mL com 75% de etanol a 4 ° C, ou proceder de imediato à etapa seguinte.
- Desidrate as amostras em uma série de álcool butílico terciário.
- Substitua a solução de etanol 75% com 10 mL de álcool butílico terciário de 75% (TBA) para cobrir generosamente as amostras e incube-os por 1,5 h. Uma pipeta de Pasteur pode ser útil para descartar a solução.
- Repita a etapa 2.2.1, adicionando TBA com concentrações crescentes (85%, 95% e 100%, v/v; e 3x com TBA pura) em um forno de secagem a 30 ° C, com extração de ar, desde TBA puro cristaliza-se em temperatura ambiente (< 20 ° C) e é muito volátil e tóxico. Se TBA cristaliza-se com a amostra, prejudicaria os tecidos.
- Incorpore as amostras em parafina18.
- Derreta a parafina, introduzindo as pérolas de parafina na secagem estufa a 60 ° C, com extração de ar do dia anterior. As pérolas de parafina podem também ser derretidas em uma chapa de fogão, mas certifique-se de que a parafina líquida é a 60 ° C e não a uma temperatura mais elevada, para evitar danos aos tecidos.
- Substituir o TBA com uma mistura de óleo de TBA e parafina (1:1) e incube por 24h dentro de um forno de secagem a 60 ° C. Em seguida, substituir a mistura de TBA e com puro óleo de parafina derreter parafina e incubar durante pelo menos 6 h dentro de um forno de secagem a 60 ° C. Repetir 2x e incubar a última alteração pelo menos 4-6 d.
- Coloque cada amostra em um pequeno molde base metal sobre uma superfície de calor, incorporado em cera de parafina e passo a fita de encastre. Coloque-o sobre uma superfície fria e remover o bloco, uma vez que a cera é solidificada.
- Seção e reidratar os preparativos.
- Prepare o adesivo do Haupt: dissolver 1 g de gelatina de Knox Lisa em 100 mL de água destilada a 30 ° C; em seguida, adicione 2 g de cristais de fenol solto (C6H5OH) e 15 mL de glicerol. Adesivo do Haupt espalhados por uma lâmina de vidro com uma escova e adicionar uma gota de solução de formol a 1%.
- Seção cada parafina bloco a 10 µm em um micrótomo rotatórios e colocar as seções sobre a lâmina de vidro coberta com adesivo do Haupt.
- Coloque as lâminas de vidro com as seções sobre uma superfície de calor a 35-40 ° C até secar; em seguida, mova as lâminas de vidro de um porta-lâminas.
- Prepare as soluções dewax e reidratação. Coloque 200 mL de Histoclear II em três pratos de vidro-manchando, mais uma com Histoclear II:ethanol (1:1, v/v), uma série de etanol (100%, 70%, 40%, v/v) e uma lavagem final de água destilada.
- Dewax as seções com três lavagens, cada 5 min, no II Histoclear e em Histoclear II:ethanol (1:1, v/v). Coloque o porta-lâminas dentro os pratos de vidro-mancha, garantindo que a solução cobre completamente os slides e, em seguida, mova o porta-lâminas de uma solução para a próxima.
- Hidratar as seções pela seguinte série de lavagens de 2 min: uma série de etanol (100%, 70%, 40%, v/v) e uma lavagem final de água destilada. Finalmente, seca as lâminas de vidro à temperatura ambiente.
- Manche as seções18.
- Eu preparar2mancha KI: dissolver 2 g de iodeto de potássio (KI) e 0,2 g de iodo (eu2) em 100 mL de água destilada. Aplique uma gota fresca eu2KI sobre as seções por 5 min e, em seguida, descartar o excesso de mancha de absorvê-lo com um toalhete de papel. Rapidamente, prossiga para a próxima etapa.
- Aplicar uma pequena gota de uma mídia de montagem sintética, coloque um pequeno vidro de tampa em cima e pressione com força. Uma vez que os meios de comunicação de montagem secam, observe sob um microscópio de campo claro para uma avaliação preliminar do amido do ovário.
Nota: Este passo não é obrigatório, embora é obtido um melhor contraste entre o amido e o plano de fundo. Se a seção tem que ser reutilizado com outras manchas após a quantificação de amido, coloque o tampa de vidro sobre a gota de mancha e elimine o excesso de absorvê-lo com um toalhete de papel (ver 2.5.1). Em seguida, após a quantificação de amido, lavar para fora o que2KI mancha com água destilada e coloque a preparação em fogo de superfície a 35-40 ° C até secar.
3. quantificação do teor de amido
- Calibre as condições ópticas.
Nota: Os níveis de deteção usados pelo analisador de imagem para detectar o amido manchado dependem directamente as condições de luz e ampliação do microscópio; assim, corrigi essas condições para todos os preparativos avaliadas. Adaptar-se a adaptação aqui proposta ao microscópio disponível e as condições de luz.- Ajuste o diafragma de abertura nas 20 X ampliação e o controle de brilho ou intensidade de luz.
- Certifique-se que existem sem filtros no porta-filtro e selecione uma condição brilhante-campo no microscópio. Selecione uma ampliação apropriada (por exemplo, 40 X para doce de cereja Primórdio de ovário).
- Controle as condições de aquisição de imagem. Ajustar as configurações da câmera com uma preparação manchada sem tecido através de imagem | Aquisição | Pré-visualização.
- Corrigir o brilho em 50%, o ganho em 1.0 x e os indicadores de histograma do gamma valor em 1,00 e o contraste no 0 - 100, alinhando-se com os limites do histograma de distribuição de brilho.
- Activar a função de subexposição/superexposição e ajustar o tempo de exposição no limite de superexposição.
- Aplica a função de balanço de branco a imagem completa para exibir todos os componentes de cor neutra da imagem sem qualquer tonalidade de cor e a correção de sombreamento para a imagem completa para criar uma imagem corrigida e homogênea.
- Calibre o sistema de análise de imagem para obter os valores de controle de nível de cinza (0, preto; 255, branco) de diferentes densidades ópticas (OD) que são obtidos pelos valores de transmitância (T).
- Adquirir uma imagem de uma preparação manchada sem tecido, considerado controle branco, e medir o nível de cinza da imagem em preto e branco através de medida | Cinza medida | Calibrar o cinza | Valor de referência = 0 | Medida | Calibrar | Okey. Isso corresponde com uma 100% de transmitância; assim, uma densidade óptica de 0, de acordo com OD = 2 - log T.
- Adquirir uma imagem da mesma preparação com um filtro 4N, que reduz a quantidade de luz 4 x e medir o nível de cinza da imagem em preto e branco através de medida | Cinza medida | Calibrar o cinza | Valor de referência = 0.6 | Medida | Calibrar | Okey. Isso corresponde com a transmitância de 25%; assim, uma densidade óptica de 0,6, de acordo com OD = 2 - log T.
- Adquirir uma imagem da mesma preparação sem luz, considerado negro, e medir o nível de cinza da imagem em preto e branco através de medida | Cinza medida | Calibrar o cinza | Valor de referência = 1 | Medida | Calibrar | Okey. Isso corresponde com uma 0% de transmitância; assim, uma densidade óptica de 1 segundo OD = 2 - log T.
- Detecta amido.
- Adquira uma imagem colorida do campo para medir em formato TIFF com resolução de pelo menos 300 pontos por polegada (dpi).
- Crie uma imagem binária correspondente à área manchada. Definir os limites de três cores (valores entre 0 - 255 para cada um) até que a imagem binária reflete exatamente os grânulos de amido manchada observados, através de imagem | Detectar | Selecione os limiares de vermelho, azul e verde | Okey. Fazer comparações visuais repetidamente em várias preparações e tecidos para afinar os níveis finais da deteção. Armazenar e usar estes níveis para todas as preparações.
- Quantificar o amido. Converta a imagem original de cor em uma imagem preto e branca com o sistema de análise de imagem. Use a imagem binária como uma máscara sobreposta na imagem em preto e branco através de imagem | Editar binário. Medir a soma da densidade óptica de cada pixel sob a máscara através de medida | Nível de cinza | Okey e considerar esse valor como o teor de amido no campo medido.
- Repita as etapas 3.4.1 - 3.4.3 em quatro lugares do primordia de ovário para obter um valor representativo do teor de amido em ovário de primordia de flor.
- Repita as etapas 3.4.1 - 3.4.3 e passo 3.4.5 em flores diferentes de cada data de coleta.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Estudos de letargia exigem a determinação do momento quando forem satisfeitos os requisitos de refrigeração. Apesar da falta de mudanças fenológicas durante o inverno sob condições de campo (figura 1A), árvores de cereja não recuperar a capacidade de crescimento em condições adequadas até eles passam um determinado período sob baixas temperaturas. A transferência regular dos rebentos de uma câmara de condições controladas (figura 1B) durante o tempo de inverno permitiu a avaliação do estado de letargia dos botões da flor. A avaliação do crescimento do broto de flor foi feita medindo-se o aumento de peso de broto. Enquanto, durante a dormência, nenhuma alteração foi observada após 10 dias de condições apropriadas (Figura 1), uma vez que era superar a letargia, os gomos incharam e explosão na câmara de crescimento (Figura 1). Os resultados desta análise permitiu o status de dormência dos botões a ser estabelecida. Devido às diferentes temperaturas que ocorrem durante o inverno, a dormência foi superada em datas diferentes, dependendo do ano. Enquanto que, durante o primeiro ano do estudo, a quebra de dormência ocorreu em Janeiro, o segundo ano apresentou um inverno mais ameno; assim, o cumprimento de refrigeração ocorreu cerca de três semanas depois, em fevereiro.
Doce de cereja tem botões florais em esporas, onde o broto apical é um broto vegetativo e os botões laterais são botões de flores (Figura 1 e 1 D). Gustativas indiferenciadas começaram a se diferenciar em flor ou brotos vegetativos no final do verão, e quando eles entram a dormência no inverno, várias primordia flor permanece dentro do broto, protegidos por numerosas escamas verdes e coberto por escamas exteriores marrom ( Figura 1 e Figura 2A). A dissecação do bud flor mostrou o primordia de florzinha dentro (Figura 2A). Cada flor continha primordia de flores individuais de um a cinco anos (Figura 2B). Apesar do pequeno tamanho do primórdio cada flor, todas as partes de uma flor são diferenciadas e podem ser distinguidas: o pistilo, as anteras, as pétalas e sépalas (Figura 2). O uso de técnicas histoquímicas (álcool: acética [3:1] fixação, incorporação de cera de parafina, seccionamento de micrótomo e coloração à base de iodo amido) permitiu a distribuição do amido dentro a flor tecidos Primórdio a serem observados (Figura 2D ).
Amido no Primórdio ovário foi quantificado em cada seção. Quatro medidas de µm 13372, na ampliação de 40x, representaram o layout geral do amido no Primórdio de ovário doce de cereja (Figura 3A). Grânulos de amido foram claramente distintos do fundo após eu2KI coloração (Figura 3B). O amido foi identificado pelo sistema de análise de imagem, ajustando a cor limiares de vermelho, verde e azul até que todos os grânulos de amido observaram foram cobertos pela imagem binária criada pelo sistema com base nos parâmetros de cor definida (Figura 3). Os valores obtidos de teor de amido foram o resultado da medida da densidade óptica de cada pixel sob a máscara da imagem preto e branco (Figura 3D).
A quantificação de amido revelou um padrão consistente de amido dinâmico durante o inverno (Figura 4). Consistentemente, a quantidade de amido no inverno adiantado apresentou um valor de densidade óptica de menos de 40.0003, enquanto o montante máximo alcançado um valor entre 120.000 e 140.000 em dois anos. Enquanto o valor máximo foi alcançado em janeiro durante o primeiro ano (Figura 4A), ocorreu em fevereiro, no segundo ano (Figura 4B). Contrastando com estes resultados com a tez de dormência, a quantidade máxima de amido ocorreu concomitantemente com o cumprimento de refrigeração em dois anos.
Essa abordagem requer a determinação do estado de dormência (Figura 5A) concomitantemente com a quantificação de amido no tecido do ovário (Figura 5B) para enquadrar as alterações do teor de amido em relação à dormência.

Figura 1: montagem Experimental para a determinação do estatuto de botões de flores de cereja doce letargia. (A) ramos, durante o inverno, mostram os botões dormentes, fechado e coberto por escamas marrons escuras. (B), este painel mostra brotos transferidos para a câmara de crescimento. Em meados de Janeiro, alguns cultivars permanecidos dormentes com os botões ainda fecharam (seta branca), enquanto outros foram capazes de crescer, apresentando gomos expandidos (seta preta). (C) este painel mostra um detalhe de uma filmagem com botões de flores dormentes localizados lateralmente e um único broto vegetativo localizado na posição apical no calor. (D), este painel mostra um detalhe de um broto, uma vez que dormência foi superada após 10 dias em câmara de crescimento, apresentando inchaço de broto. Clique aqui para ver uma versão maior desta figura.
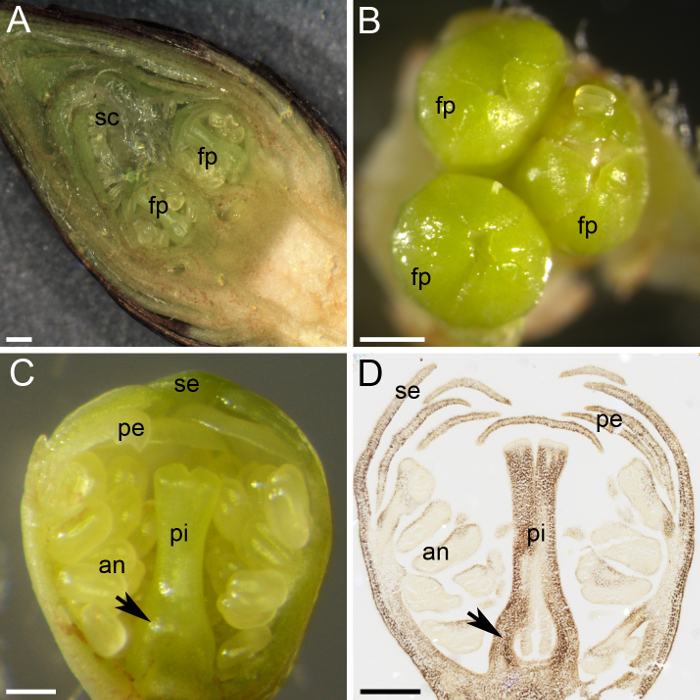
Figura 2: preparação de material para a quantificação do amido da planta. (A), este painel mostra uma seção transversal de uma flor em botão, mostrando dois primordia flor (fp) protegida por numerosas escamas (sc). (B) três primordia de flor (fp) reúnem-se em uma flor em botão. Este painel mostra uma seção transversal de um Primórdio de flor, (C) com todos os verticilos diferenciadas: sépalas (se), pétalas (pe), anteras (uma) e pistilo (pi). Distingue-se do primórdio de ovário na base do pistilo (seta). (D) A seção intermediária do primórdio flor foi coletada e fixo em Janeiro, incorporada em cera de parafina, seccionada e manchada com eu2KI (marrom escuro) para amido. Este painel mostra o Primórdio do ovário (seta). As barras de escala são 500 µm em painéis A e B e 200 µm em painéis C e D. Clique aqui para ver uma versão maior desta figura.

Figura 3: quantificação em doce de cereja Primórdio de ovário de amido. (A), este painel mostra uma seção média do primórdio ovário2KI, mostrando os quatro quadros na qual amido conteúdo manchado com foi medido. (B), este painel mostra um detalhe do primórdio do ovário. Os grânulos de amido são corados em marrom escuro. (C), este painel mostra uma imagem pseudocolorida no qual amido corresponde a diferentes tons de azul. (D), este painel mostra uma máscara de imagem binária eu cobrindo2KI-manchado amido (azul) na imagem original preto e branca. A densidade óptica é medida apenas em pixels da imagem original coberto pela máscara. As barras de escala são 100 µm no painel A e 20 µm em painéis B - D. Clique aqui para ver uma versão maior desta figura.

Figura 4: resultados representativos da quantificação amido no doce de cereja primordia de ovário coletados mensalmente do Outono à Primavera durante dois anos, das condições de temperatura diferentes inverno. (A), este painel mostra os resultados dos anos 2010-2011, que teve um inverno frio. O cumprimento de refrigeração (floco de neve) ocorreu em Janeiro, concomitantemente com a quantidade máxima de amido. (B), este painel mostra os resultados dos anos 2011-2012, que teve um inverno ameno. O cumprimento de refrigeração (floco de neve) ocorreu em fevereiro, concomitantemente com a quantidade máxima de amido. Os valores são a média ± erro padrão da média. Clique aqui para ver uma versão maior desta figura.
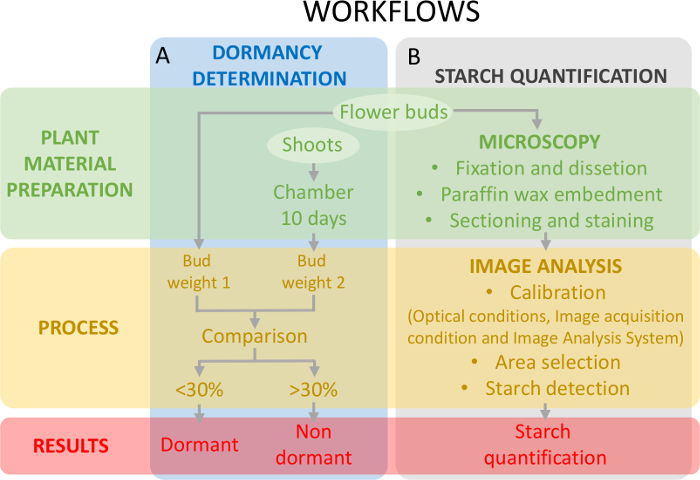
Figura 5: esquema do desenho experimental para avaliar o estado de dormência de gemas e a quantificação de amido no primordia de ovário em doce de cereja. (A), este painel mostra os fluxos de trabalho da determinação do estado de letargia: a preparação de material de planta, o processo e os resultados obtidos. (B), este painel mostra os fluxos de trabalho da quantificação amido: histochemical preparação dos botões para uma observação microscópica do amido ou da fécula, a detecção de análise de imagem de amido e a quantificação de amido. Clique aqui para ver uma versão maior desta figura.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Dormência em plantas perenes lenhosas apresenta claras implicações na produção de frutas e florestais em mudanças climáticas, embora o processo biológico por trás de dormência permanece obscuro. Estudos de dormência podem ser abordados de diferentes pontos de vista, mas a pesquisa à procura de um marcador biológico para a letargia do inverno tem se intensificado nos últimos anos. No entanto, a maioria das tentativas de encontrar um inequívoco indicador mostrando quando um botão está quebrado letargia tem sido vencida3. A metodologia descrita aqui, combinando técnicas histoquímicas18 com imagem de análise, tem sido muito útil para examinar a relação entre reservas de hidratos de carbono de um tecido especial e sua atividade fisiológica nos botões doce de cereja durante as diferentes fases de letargia5 e também pode ser aplicado a outras espécies e tecidos10,13,20.
Reservas de carboidrato na forma de amido desempenham um papel importante no desenvolvimento da flor e o processo reprodutivo7,10,13,20,21,23 e na sazonalidade dos temperados perenes lenhosas6. Diferentes estudos sobre letargia prestado atenção ao amido dentro os gomos24. No entanto, devido ao seu pequeno tamanho, vários botões todo são necessários para a utilização de métodos de análise quantitativas e a detecção de variações quantitativas em particulares tecidos ou células é limitada devido ao efeito de mascaramento das células circundantes. A combinação das técnicas histoquímicas com o sistema de análise de imagem oferece uma boa oportunidade para estudar as alterações do teor de amido das diferentes estruturas dentro do broto.
Este método tem a limitação de evitar que a quantificação do conteúdo exato de amido no tecido, mas permite que o conteúdo de amido relativo a quantificar a seguir amido quantitativo mudanças ao longo do tempo25 e comparar o amido conteúdo de diferentes tecidos13,20, cultivares ou anos5. A fim de permitir a comparação correta entre os valores de densidade óptica entre campos, tecidos e botões, a calibração do sistema (condições de luz, coloração intensidade e ampliação) e a fixação de limites devem ser estabelecidos com precisão de cor , armazenados e utilizados para todos os preparativos.
Coloração de amido e quantificação com base em iodo de potássio permite o uso subsequente de outras manchas após a lavagem da seção. Assim, diferentes análises podem ser executadas sem preparações adicionais e montagem sintética mídias não são utilizadas5,20. Da mesma forma, de medidas morfométricas26 pode ser feito após a preparação do mesma, permitindo que o padrão de acúmulo de amido, para ser enquadrada em relação ao crescimento das diferentes estruturas5,13. O método pode ser adaptado a outras espécies com o ajuste dos níveis de cor que usam o analyzer para detectar o amido, que pode permitir que o estudo de outros processos de desenvolvimento que envolvem mudanças no teor de amido em pequenos grupos de células ou estruturas.
A relação entre a liberação de letargia e amido acumulação no primordia de ovário revelada pelo uso deste método fornece uma base sólida para compreender a base biológica da letargia e arrepiante requisitos5. No entanto, quantificação de amido por análise de imagem em secções de parafina pode se tornar muito complicado e demorado para estimar os requisitos de refrigeração de um grande número de cultivares. Os futuros esforços tem que ser focado em estudar indicadores biológicos confiáveis que facilmente podem indicar o estado de letargia da árvore. Entretanto, o padrão de variações de amido durante a dormência pode ser usado para enquadrar estudos fisiológicos e genéticos, e a combinação das técnicas histoquímicas com análise de imagem descritos neste documento pode ser usada em outras culturas perenes lenhosas para quantificar o teor de amido de diferentes tecidos em relação à dormência.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Os autores não têm nada para divulgar.
Acknowledgments
Os autores agradecem agradecer Maria Herrero e Eliseo Rivas sua discussão útil e conselhos. Este trabalho foi apoiado pelo Ministerio de Economía y competitividade — Fundo Europeu de Desenvolvimento Regional, a União Europeia [número de concessão BES-2010-037992 para E. F.]; o Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria [concessão números RFP2015-00015-00, RTA2014-00085-00, RTA2017-00003-00]; e o Gobierno de Aragón — Fundo Social Europeu, a União Europeia [Grupo Consolidado A12-17R].
Materials
| Name | Company | Catalog Number | Comments |
| Precision scale | Sartorius | CP225D | |
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Drying-stove | Memmert | U15 | |
| Paraffin Embedding station | Leica Microsystems | EG1140H | |
| Rotatory microtome | Reichert-Jung | 1130/Biocut | |
| Microtome blade | Feather | S35 | Stainless steel |
| Bright field microscope | Leica Microsystems | DM2500 | |
| Digital Camera | Leica Microsystems | DC-300 | |
| Image Analysis System | Leica Microsystems | Quantiment Q550 |
References
- Kurokura, T., Mimida, N., Battey, N. H., Hytönen, T. The regulation of seasonal flowering in the Rosaceae. Journal of Experimental Botany. 64 (14), 4131-4141 (2013).
- Rohde, A., Bhalerao, R. P.
- Fadón, E., Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environmental and Experimental Botany. 152, 28-36 (2018).
- Fadón, E., Rodrigo, J., Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees - Structure and Function. , In Press (2018).
- Fadón, E., Herrero, M., Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Frontiers in Plant Science. 9 (171), (2018).
- Loescher, W. H., Mccamant, T., Keller, J. D. Carbohydrate reserves, translocation and storage in woody plant roots. HortScience. 25 (3), 274-281 (1990).
- Hedhly, A., et al. Starch turnover and metabolism during flower and early embryo development. Plant Physiology. , (2016).
- Rodrigo, J., Hormaza, J. I., Herrero, M. Ovary starch reserves and flower development in apricot (Prunus armeniaca). Physiologia Plantarum. 108 (1), 35-41 (2000).
- Julian, C., Rodrigo, J., Herrero, M. Stamen development and winter dormancy in apricot (Prunus armeniaca). Annals of Botany. 108 (4), 617-625 (2011).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil starch reserves at anthesis correlate with final flower fate in avocado (Persea americana). PLoS One. 8 (10), e78467 (2013).
- Smith, A. M., Zeeman, S. C. Quantification of starch in plant tissues. Nature Protocols. 1 (3), 1342-1345 (2006).
- Eliceiri, K. W., et al.
- Rodrigo, J., Herrero, M. Influence of intraovular reserves on ovule fate in apricot (Prunus armeniaca L.). Sexual Plant Reproduction. 11, 86-93 (1998).
- Zhou, J., Spallek, T., Faulkner, C., Robatzek, S. CalloseMeasurer: A novel software solution to measure callose deposition and recognise spreading callose patterns. Plant Methods. 8 (1), (2012).
- Faulkner, C., et al. An automated quantitative image analysis tool for the identification of microtubule patterns in plants. Traffic. 18 (10), 683-693 (2017).
- Kuhn, B. F. Determination of starch in ovules of the sour cherry cv. "Stevnsbaer.". European Journal of Horticultural Science. 71 (3), 120-124 (2006).
- Johansen, D. A. Plant microtechnique. , McGraw-Hill. New York, NY. (1940).
- Ruzin, S. E. Plant microtechnique and microscopy. , Oxford University Press. New York, NY. (1999).
- Rodrigo, J., Rivas, E., Herrero, M. Starch determination in plant tissues using a computerized image analysis system. Physiologia Plantarum. 99 (1), 105-110 (1997).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Ovary starch reserves and pistil development in avocado (Persea americana). Physiologia Plantarum. 140 (4), 395-404 (2010).
- Suarez, C., Castro, A. J., Rapoport, H. F., Rodriguez-García, M. I. Morphological, histological and ultrastructural changes in the olive pistil during flowering. Sexual Plant Reproduction. 25, 133-146 (2012).
- Lang, G. A., Early, J. D., Martin, G. C., Darnell, R. L. Endodormancy, paradormancy, and ecodormancy - Physiological terminology and classification for dormancy research. HortScience. 22 (3), 371-377 (1987).
- Hedhly, A., Vogler, H., Eichenberger, C., Grossniklaus, U. Whole-mount clearing and staining of arabidopsis flower organs and siliques. Journal of Visualized Experiments. 2018 (134), 1-10 (2018).
- Kaufmann, H., Blanke, M. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. Journal of Plant Physiology. 218 (July), 1-5 (2017).
- Herrero, M., Dickinson, H. G. Pollen-pistil incompatibility in Petunia hybrida: changes in the pistil following compatible and incompatible intraspecific crosses. Journal of Cell Science. 36, 1-18 (1979).
- Carpenter, A. E., et al. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biology. 7 (10), R100 (2006).
