

Summary
Vi præsenterer en metode til at kvantificere stivelsesindhold i æggestokken primordia i søde kirsebær (Prunus avium L.) i løbet af vinteren vækstdvale ved hjælp af et billede analysesystem kombineret med histokemiske metoder.
Abstract
Ændringer i stivelse i små strukturer er associeret med vigtige begivenheder under flere plante udviklingsprocesser, herunder den reproduktive fase fra bestøvning til befrugtning og udbrud af frugtsætning. Dog er variationer i stivelse under flower differentiering ikke helt kendt, hovedsagelig på grund af vanskeligheden ved at kvantificere stivelsesindhold i de særligt små strukturer af blomst primordia. Her, beskriver vi en metode til kvantitativ bestemmelse af stivelse i æggestokken primordia af søde kirsebær (Prunus avium L.) ved hjælp af et billede analysesystem knyttet til mikroskop, der gør det muligt for vedrørende ændringer i stivelsesindhold med de forskellige faser af vækstdvale fra efteråret til foråret. Med henblik herpå bestemmes vækstdvale status af blomsterknopper ved at evaluere bud vækst af skuddene overført til kontrollerede forhold på forskellige tidspunkter i vinteren. Til kvantificering af stivelse i æggestokkene primordia, blomsterknopper sekventielt indsamles, fastsættes, indlejret i paraffinvoks, sectioned, og farves med jeg2Kl (kaliumiodid-jod). Præparater, der er observeret under mikroskop og analyseres af et billede analyzer, der klart adskiller stivelse fra baggrunden. Stivelse indhold værdier fås ved måling af Ekstinktionen af det billede, der svarer til de farvede stivelse, overvejer summen af Ekstinktionen af hver enkelt pixel som et skøn over rammen studerede stivelsesindhold.
Introduction
Tempereret woody stauder tilpasses sæsoner af modulere deres vækst og udvikling. Mens de udvikler i løbet af foråret og sommeren, stoppe de voksende i løbet af efteråret at gå hvilende i vinteren1. Selvom vækstdvale tillader dem at overleve ved lave vintertemperaturer, er nedkøling en forudsætning for en korrekt budburst i foråret2. De vigtige implikationer af vækstdvale i tempererede frugtproduktion og skovbrug har ført til forskellige bestræbelser på at bestemme og forudsige vækstdvale perioden3. I frugt træarter, empiriske forsøg overføre skud at tvinge betingelser og statistiske forudsigelser baseret på data blomstring er nuværende metoder til at bestemme datoen for bruddet på vækstdvale, som giver forskere til at anslå den nedkøling krav for hver sort. Sådan bestemme vækstdvale status baseret på biologiske processer er imidlertid uklart3.
Blomstring i tempererede frugttræer, såsom søde kirsebær (Prunus avium L.), forekommer en gang om året og varer omkring to uger. Men blomsterne begynde at differentiere og udvikle omkring 10 måneder tidligere, under den forrige sommer4. Blomst primordia stoppe voksende i løbet af efteråret at forblive sovende inde knopper i løbet af vinteren. I denne periode skal hver kultivar ophobes en isnende krav for korrekt blomstring4. Trods manglen fænologiske ændringer i knopper i løbet af vinteren, blomst primordia er fysiologisk aktive under vækstdvale, og ophobningen af nedkølingen temperaturer har været for nylig forbundet med dynamikken i stivelse ophobning eller formindske i cellerne i æggestokken primordium, tilbyder en ny tilgang til vækstdvale bestemmelse5. Men den lille størrelse og placering af æggestokken primordium kræver en særlig metode.
Stivelse er den store opbevaring kulhydrat i woody plant arter6. Således, ændringer i stivelse er relateret til den fysiologiske aktivitet af blomst væv, som har brug for kulhydrater til at støtte deres udvikling7,8. Forskellige vigtige begivenheder i løbet af reproduktive processen er også relateret til variationer i stivelsesindhold i forskellige blomster strukturer, som anther meiose9, væksten af pollen rør gennem stil eller ovule befrugtning10. Histokemiske metoder tillade påvisning af stivelse i hver bestemt væv af blomst primordia under vækstdvale. Vanskeligheden er imidlertid fortsat i kvantificere stivelsen tillade efter dens mønster af ophobning/fald over tid eller sammenligne stivelsen indhold blandt væv, kultivarer eller år. Dette skyldes den lille mængde væv tilgængelig for analytiske teknikker11. Som et alternativ, billedanalyse knyttet til mikroskopi12 giver mulighed for kvantificering af stivelsen i meget små prøver af plante væv13.
Tilgange kombinerer mikroskopi og billede analyse har været brugt til at kvantificere indholdet af forskellige komponenter i plantevæv, såsom callose14, microtubes15, eller stivelse16, af måling af størrelsen af området farvet af specifikke pletter. For stivelse, det kan let påvises ved hjælp af kaliumiodid-jod (jeg2KI) reaktion17. Denne metode er meget specifikke. Jeg2KI intercalates inden for stivelse korn laminar struktur og danner en mørk blå eller rødlig-brun farve, afhængigt af amylose indholdet af stivelse18. Sektioner farves med jeg2KI pletten Vis passende kontrast mellem stivelse og baggrunden væv, giver en utvetydig stivelse påvisning og den efterfølgende kvantificering af billede analyse system19. Selv om dette farvestof ikke er støkiometriske, er ophobning af jod proportional med længden af stivelse molekyle, der stærkt kan variere17. Således, størrelsen på de farvede område, udtrykt som antallet af pixels kan ikke nøjagtigt afspejle indhold af stivelse, da høje forskelle i stivelsesindholdet af blev fundet mellem felter med farvede områder af samme størrelse. Som et alternativ, kan stivelsesindhold vurderes ved at måle Ekstinktionen af granulerne farves på sort-hvide billeder fremstillet af mikroskop, som det er blevet rapporteret i forskellige væv i abrikos8,13 , 19, avocado10,20, og oliven21.
Her, beskriver vi en metode, der kombinerer den eksperimentelle bestemmelse af vækstdvale status med kvantificeringen af stivelsesindhold i æggestokken primordium væv fra efteråret til foråret i søde kirsebær, tilbyder et nyt værktøj til forståelse og forudsigelse af vækstdvale baseret på studier af de biologiske mekanismer forbundet med vækstdvale.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. vækstdvale beslutsomhed og plante materiale indsamling
- Prøve blomsterknopper i feltet. Vækstdvale undersøgelser er langsigtet eksperimenter og kræver voksen træer stor nok til at indsamle knopper og skyder hele vinteren uden at kompromittere træer udvikling i løbet af de næste forår. Særlige orchard forvaltning kunne være påkrævet afhængigt af uddannelsessystem; beskæring kan således mindre alvorlig end for frugt produktionsformål.
- Hver uge, fra starten af efteråret indtil starten af opløbet pause, indsamle og vejer 10 blomsterknopper.
- Fix blomsterknopper ved at placere dem i en 10 mL glasrør med en hætte og blød prøverne i en Fikseringsvæske løsning af ethanol/eddikesyre (3:1) i mindst 24 timer ved 4 ° C. Derefter, kassere fiksativ og tilsættes 75% ethanol, generøst at sikre, at det dækker prøverne. Prøverne kan bevares i denne løsning ved 4 ° C indtil brug.
Bemærk: Vakuum infiltration kan bruges til at fjerne luftbobler inde i opløbet og forhindre dem i at flydende. Dette letter fiksativ indtrængen i væv men kunne skade struktur af væv. Prøv at undgå det hvis ikke nødvendige.
- Prøve skud i feltet.
- Hver uge, fra starten af efteråret indtil starten af opløbet pause, tag tre skud på 15-30 cm i længden og 5 mm i diameter, som indeholder mindst 10 blomsterknopper hver. Placer dem på vand-gennemblødt Blomsterhandler skum i en vækst kammer på 22 ± 1 ° C med en 12-timers lys lysperiode.
- Efter 10 dage i salen, vækst, vælge og vejer 10 blomsterknopper fra skuddene.
- Evaluere bud vækst og bestemme vækstdvale status. Stikprøveperioden har tilpasses til betingelserne i placeringen. I plantagen betingelser (Zaragoza, Spanien, 41 ° 44'30 "N, 0 ° 47'00" W, og 220 m over havets overflade), prøveudtagning af skuddene blev gennemført fra 30 November indtil slutningen af februar eller starten af marts.
- Ugentlig evaluere svar af blomsterknopper til de passende vækstbetingelser for afdeling, fra begyndelsen af efteråret indtil budburst i foråret, ved at sammenligne vægten af 10 knopper plukket fra feltet.
- Hvis der er ingen forskelle eller disse forskelle er mindre end 30%, overveje at knopperne ikke har opfyldt deres isnende krav og er endodormant22. Hvis forskellene er mere end 30%, overveje at knopper har opfyldt kravene til nedkøling og ecodormant22.
2. plante materiale forberedelse til stivelse kvantificering
- Vælg omkring seks faste knopper fra hver prøveudtagning dato (Se trin 1.1). Afhængigt af kultivarer indeholder hver søde kirsebær blomsterknop op til fem blomst primordia.
- Fjern eksterne bud skalaer og placere de bud på en urmager glas med 75% ethanol til at forhindre udtørring.
- Dissekere opløbet og udvinde mindst én blomst primordium pr. bud ved hjælp af præcision pincet og en oftalmologisk skalpel under en stereoskopisk mikroskop. Blomst primordium kan bevares i et 10 mL glasrør med 75% ethanol ved 4 ° C, eller Fortsæt til næste trin, straks.
- Dehydrere prøver i en tertiær butyl alkohol serie.
- Erstatte 75% ethanol opløsning med 10 mL af 75% tertiære butyl alkohol (TBA) til generøst dækker prøverne og inkuberes dem for 1,5 h. Pasteur pipette kan være nyttigt at skille sig af med løsningen.
- Gentag trin 2.2.1, tilføje TBA med stigende koncentrationer (85%, 95% og 100% v/v; og 3 x med ren TBA) i en tørring ovn ved 30 ° C med luft udsugning, da ren TBA krystalliserer ved stuetemperatur (< 20 ° C) og er meget flygtige og giftige. Hvis TBA krystalliserer med prøven, vil det skade væv.
- Integrere prøverne i paraffin18.
- Smelte paraffin ved at indføre paraffin perler i tørring ovnen ved 60 ° C med luft udsugning den foregående dag. Paraffin perler også kan smeltes på en kogeplade, men sikre, at de flydende paraffin er på 60 ° C og ikke ved en højere temperatur, at undgå vævsskader.
- Erstatte TBA med en blanding af TBA og paraffin olie (1:1), og der inkuberes i 24 timer inde i en tørring ovn ved 60 ° C. Derefter erstatte blanding af TBA og paraffinolie med rene smelte paraffin og Inkuber i mindst 6 h inde en tørring ovn ved 60 ° C. Gentag det 2 x og Inkuber den sidste ændring for mindst 4-6 d.
- Placer hver prøve på et lille metal base mug over en varme overflade, indlejret i paraffinvoks og tempo indlejring kassetten. Placer det over en kold overflade og fjerne blokken, når voks er størknet.
- Sektion og rehydrere forberedelserne.
- Forberede Haupts lim: opløse 1 g af plain Knox gelatine i 100 mL destilleret vand ved 30 ° C; derefter tilsættes 2 g af phenol løs krystaller (C6H5OH) og 15 mL glycerol. Sprede den Haupt limen over et glas dias med en pensel og tilføje en dråbe af en 1% formaldehyd løsning.
- Sektion hver paraffin blokere på 10 µm i en rotatorisk mikrotomen og placere sektioner på glas dias dækket med Haupts klæbemiddel.
- Placer glas dias med sektioner over en varme overflade på 35-40 ° C indtil tør; derefter, flytte glas dias til et dias rack.
- Forberede dewax og rehydrering løsninger. Placere 200 mL af Histoclear II i tre glas-farvning retter, en anden en med Histoclear II:ethanol (1:1, v/v), en ethanol serie (100%, 70%, 40% v/v), og en afsluttende destilleret vand vask.
- Afvoksning sektioner med tre vasker, hvert 5 min, i Histoclear II og i Histoclear II:ethanol (1:1, v/v). Placer slide rack inde de glas-farvning retter, at sikre, at løsningen helt dækker dias, og derefter flytte diaset rack fra én løsning til næste.
- Rehydrere afsnittene af den følgende række af 2-min vasker: en ethanol serie (100%, 70%, 40% v/v) og en afsluttende destilleret vand vask. Endelig tør glas dias ved stuetemperatur.
- Pletten afsnit18.
- Forberede jeg2KI pletten: Opløs 2 g kaliumjodid (KI) og 0,2 g jod (jeg2) i 100 mL destilleret vand. Påfør en dråbe af friske jeg2KI over afsnit2 i 5 min og derefter slette overskydende af pletten ved at absorbere det med en trækpapir. Hurtigt videre til næste trin.
- Påfør en lille dråbe af et syntetisk montering medier, placere en lille cover glas på toppen og tryk hårdt. Når montering media tørrer, observere under en lysfelt mikroskop for en indledende vurdering af æggestokken stivelse.
Bemærk: Dette trin er ikke obligatoriske, selvom en bedre kontrast mellem stivelsen og baggrunden er opnået. Hvis afsnittet skal genbruges med andre pletter efter stivelse kvantificering, anbring dækslet glas over dråbe af pletten og kassér overskydende ved at absorbere det med en trækpapir (Se 2.5.1). Derefter, efter stivelse kvantificering, vaskes ud I2KI pletten med destilleret vand og sted præparater over varme overflade på 35-40 ° C indtil tør.
3. kvantificering af kartoflernes stivelsesindhold
- Kalibrere de optiske betingelser.
Bemærk: Afsløring niveauer anvendes af billede analyzer til at opdage den farvede stivelse er direkte afhængig af lysforhold og forstørrelse i mikroskopet; således fastsætte disse betingelser for alle præparater vurderes. Tilpasse den justering, der er foreslået her til rådighed mikroskop og lysforhold.- Justere blænde mellemgulvet på 20 X Forstørrelse og lysstyrkekontrol eller lysintensitet.
- Sørg for, at der er ingen filtre på filterholderen, og vælg en lyse-feltet betingelse i mikroskopet. Vælg en egnet forstørrelse (f.eks.40 X for søde kirsebær æggestokken primordium).
- Styre image erhvervelse betingelser. Justere kameraets indstillinger med en farves forberedelse uden væv via billede | Køb | Før visning.
- Lave lysstyrke på 50%, gain på 1,0 x og histogram indikatorer gamma værdi på 1,00 og kontrast på 0 - 100, foring med grænserne for lysstyrke distribution histogram.
- Aktivere funktionen overeksponering/undereksponering og justere eksponeringstiden på grænsen af overeksponering.
- Anvend funktionen hvidbalance til den komplette billede at vise alle neutral-farvede dele af billedet uden hvilken som helst farvetone og skygge korrektion til komplet billedet for at oprette en korrigeret og ensartet billede.
- Kalibrere Image Analysis System for at opnå kontrolværdierne af det grå niveau (0, sort, 255, hvid) af forskellige optiske tætheder (OD), der er fremstillet af transmittans (T) værdier.
- Tilegne sig et billede af en farves forberedelse uden væv, betragtes kontrol hvide, og måle det grå niveau af den sorte-hvidt billede via foranstaltning | Grå foranstaltning | Kalibrere grå | Referenceværdien = 0 | Foranstaltning | Kalibrere | OK. Dette svarer til en 100% lystransmission; således, en optisk tæthed på 0, ifølge OD = 2 - log T.
- Tilegne sig et billede af det samme præparat med 4N filter, der reducerer mængden af lys 4 x og måle det grå niveau af den sorte-hvidt billede via foranstaltning | Grå foranstaltning | Kalibrere grå | Referenceværdien = 0,6 | Foranstaltning | Kalibrere | OK. Dette svarer til en 25% lystransmission; således, en optisk tæthed på 0,6, ifølge OD = 2 - log T.
- Tilegne sig et billede af den samme forberedelse uden lys, betragtes som sort, og måle det grå niveau af den sorte-hvidt billede via foranstaltning | Grå foranstaltning | Kalibrere grå | Referenceværdien = 1 | Foranstaltning | Kalibrere | OK. Dette svarer med en 0% lystransmission; således, en optisk tæthed af 1, ifølge OD = 2 - log T.
- Opdage stivelse.
- Erhverve et farvebillede af feltet til at måle i TIFF-format med en opløsning på mindst 300 punkter pr. tomme (dpi).
- Oprette et binært billede svarende til området farves. Sæt de tre farve tærskler (værdier mellem 0 - 255 for hver) indtil den binært billede nøjagtigt afspejler granulerne farves stivelse observeret, via billede | Afsløre | Vælg tærskelværdierne i rød, blå og grøn | OK. Foretage gentagne gange visuelle sammenligninger i forskellige præparater og væv til at tune op de sidste opdagelse niveauer. Opbevar og Anvend disse niveauer for alle præparater.
- Kvantificere stivelsen. Konvertere det oprindelige farve billedet til et sort-hvidt billede med billedet analysesystem. Bruge den binært billede som en overlejrede maske på sort og hvidt billede via billede | Binær Edit. Måle summen af Ekstinktionen af hver pixel under den maske via foranstaltning | Grå niveau | OK og betragte denne værdi som stivelsesindhold i det målte område.
- Gentag trin 3.4.1 - 3.4.3 i fire steder i æggestokken primordia at opnå en repræsentativ værdi af stivelsesindhold i æggestokken af blomst primordia.
- Gentag trin 3.4.1 - 3.4.3 og trin 3.4.5 i forskellige blomster af hver indsamling dato.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Vækstdvale undersøgelser kræver bestemmelse af det øjeblik, når de isnende krav er opfyldt. Trods manglen fænologiske ændringer i løbet af vinteren under markforhold (figur 1A) genvinder kirsebærtræer ikke kapacitet af vækst i passende betingelser indtil de passerer en vis periode under lave temperaturer. Regelmæssig overføringen af skud til en kontrollerede forhold kammer (figur 1B) i løbet af vinteren tilladt evaluering af vækstdvale status af blomsterknopper. Evaluering af blomsterknop vækst blev gjort ved at måle bud vægtforøgelse. Mens under vækstdvale, ingen ændringer, der kunne konstateres efter 10 dage af egnede betingelser (figur 1 c), når vækstdvale blev overvundet, knopper svulmede og brast i vækst kammer (fig. 1 d). Resultaterne af denne analyse tilladt vækstdvale status af knopper til at blive etableret. På grund af de forskellige temperaturer forekommer om vinteren, blev vækstdvale overvundet på forskellige tidspunkter, afhængigt af år. Mens det første år af studiet, bruddet på frøhvile opstod i januar, det andet år præsenteret en mildere vinter; således, den isnende opfyldelse opstod omkring tre uger efter, i februar.
Søde kirsebær bærer blomsterknopper i spurs, hvor den apikale bud er en vegetativ bud og lateral knopper er blomsterknopper (figur 1 c og 1 D). Udifferentierede knopper begyndte at differentiere i blomst eller vegetativt knopper i slutningen af sommeren, og når de angiver vækstdvale i vinteren, flere blomst primordia forblive inde i den knop, beskyttet af talrige grønne skæl og er omfattet af () brun ydre skæl Figur 1 c og figur 2A). Dissektion af blomsterknop viste de lille blomst primordia inde (figur 2A). Hver blomsterknop indeholdt et til fem individuelle blomst primordia (figur 2B). Trods den lille størrelse af hver blomst primordium, alle dele af en blomst er differentierede og kan skelnes: Støvvejen, støvknapperne, kronblade og bægerblade (figur 2 c). Brugen af histokemiske metoder (alkohol: eddikesyre [3:1] fiksering, paraffinvoks indlejring, mikrotomen skæring og jod-baserede stivelse farvning) tilladt fordelingen af stivelse inden for blomsten primordium væv skal overholdes (figur 2D ).
Stivelse i æggestokken primordium var kvantificeret i hver sektion. Fire foranstaltninger af 1337 µm2, på 40 x forstørrelse, repræsenteret det generelle layout af stivelse i søde kirsebær æggestokken primordium (figur 3A). Stivelse granulat var klart adskilles fra baggrunden efter jeg2KI farvning (figur 3B). Stivelsen blev identificeret af billede analysesystem, ved at justere farven tærskler i rød, grøn og blå indtil alle stivelse granulerne observeret var omfattet af den binært billede lavet af systemet er baseret på de definerede farve parametre (figur 3 c). Værdierne af stivelsesindhold modtaget var resultatet af måling af Ekstinktionen af hver pixel under masken på den sort-hvidt billede (figur 3D).
Kvantitativ bestemmelse af stivelse afslørede et konsekvent mønster af stivelse dynamisk i løbet af vinteren (figur 4). Konsekvent, præsenteret mængden af stivelse i tidlig vinter en optisk tæthed værdi mindre end 40.0003, mens den maksimale mængde nåede en værdi mellem 120.000 og 140.000 i begge år. Mens den maksimale værdi blev nået i januar under det første år (figur 4A), det fandt sted i februar i det andet år (figur 4B). Kontrast disse resultater med teint af vækstdvale, opstod den maksimale mængde stivelse samtidig med isnende opfyldelsen i begge år.
Denne tilgang kræver bestemmelse af vækstdvale status (figur 5A) samtidig med stivelse kvantificering på æggestokkene væv (figur 5B) for at ramme ændringerne i stivelsesindhold i vækstdvale.

Figur 1: eksperimenterende set-up til bestemmelse af vækstdvale status af blomsterknopper af søde kirsebær. (A) grene, i løbet af vinteren, Vis de sovende knopper lukket og dækket af mørk brun skalaer. (B) dette panel viser skud overført til vækst kammer. I midten af januar, nogle kultivarer forblev hvilende med knopper stadig lukket (hvid pil), mens andre var i stand til at vokse, viser svulmede knopper (sort pil). (C) dette panel viser en detalje af en skyde med sovende blomsterknopper ligger lateralt og en enkelt vegetativ bud ligger i den apikale position i sporen. (D) dette panel viser en detalje af en shoot når vækstdvale blev overvundet efter 10 dage i den voksende kammer, viser bud hævelse. Venligst klik her for at se en større version af dette tal.
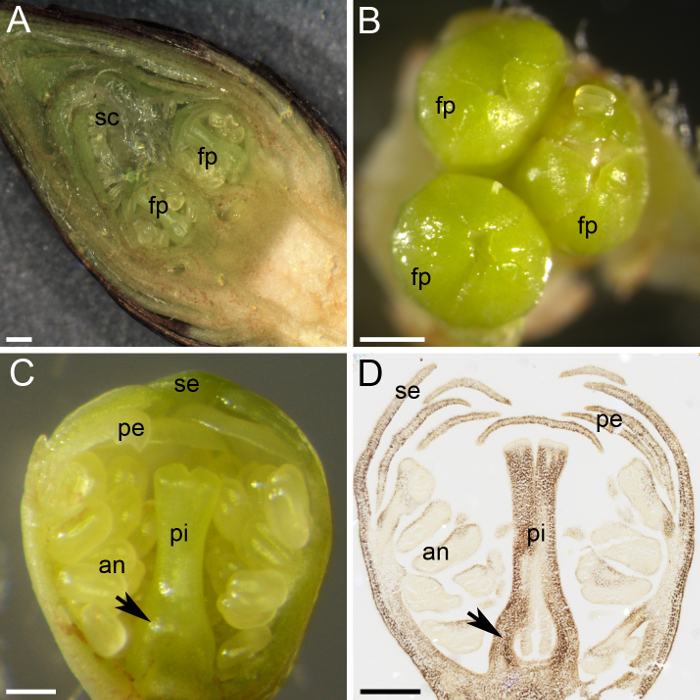
Figur 2: Plant materiale forberedelse til stivelse kvantificering. (A) dette panel viser en tværgående del af en blomsterknop, viser to flower primordia (fp) beskyttet af talrige skalaer (sc). (B) tre blomst primordia (fp) samles i en blomsterknop. (C) dette panel viser en tværgående del af en blomst primordium, med alle hvirvler opdelte: bægerblade (se), kronblade (pe), støvknapperne er (en) og Støvvejen (pi). Æggestokken primordium udmærker sig i bunden af pistil (pil). (D) A midterste del af en blomst primordium blev indsamlet og fast i januar, indlejret i paraffinvoks, snit og farves med jeg2KI (mørk brun) for stivelse. Dette panel viser æggestokken primordium (pil). Skala barer er 500 µm i paneler A og B og 200 µm i paneler C og D. Venligst klik her for at se en større version af dette tal.

Figur 3: stivelse kvantificering i søde kirsebær æggestokken primordium. (A) dette panel viser en midterste del af en æggestok primordium farves med jeg2KI, viser de fire rammer i hvilke stivelse indhold blev målt. (B) dette panel viser en detalje af æggestokken primordium. Granulerne stivelse farves i mørk brun. (C) dette panel viser en pseudocolor billede i hvilke stivelse svarer til forskellige nuancer af blå. (D) dette panel viser et binært billede maske dækker jeg2KI-farvede stivelse (blå) på de sorte og hvide originalbilledet. Ekstinktionen måles kun i pixel af det oprindelige billede dækket af masken. Skala barer er 100 µm i panelet A og 20 µm i paneler B - D. Venligst klik her for at se en større version af dette tal.

Figur 4: repræsentative resultater af stivelse kvantificering i søde kirsebær æggestokken primordia indsamlet hver måned fra efteråret til foråret i løbet af to år af forskellige vinter temperaturforhold. (A) dette panel viser resultaterne fra år 2010-2011, som havde en kold vinter. Den isnende opfyldelse (snefnug) opstod i januar, samtidig med den maksimale mængde af stivelse. (B) dette panel viser resultaterne fra år 2011-2012, som havde en mild vinter. Den isnende opfyldelse (snefnug) skete i februar, samtidig med den maksimale mængde af stivelse. Værdierne er den gennemsnit ± standardafvigelse for middelværdien. Venligst klik her for at se en større version af dette tal.
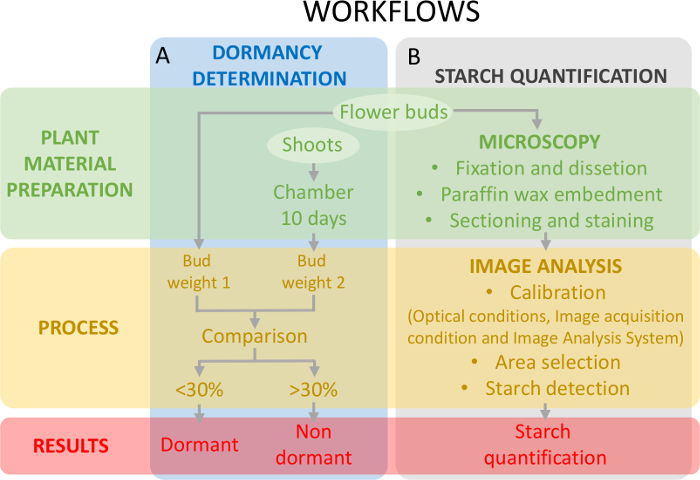
Figur 5: ordning af forsøgsdesign at vurdere vækstdvale status af knopper og stivelse kvantificering i æggestokken primordia i sød kirsebær. (A) dette panel viser arbejdsprocesser i vækstdvale status bestemmelse: plant materiale forberedelse, processen og de opnåede resultater. (B) dette panel viser arbejdsprocesser i en stivelse kvantificering: knopper histokemiske forberede en mikroskopisk observation af stivelsen, image analyse påvisning af stivelse og stivelse kvantificering. Venligst klik her for at se en større version af dette tal.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Vækstdvale i woody stauder præsenterer klart konsekvenser i frugtproduktion og skovbrug i et skiftende klima, selv om den biologiske proces bag vækstdvale uklart. Vækstdvale undersøgelser kan gribes an fra forskellige synspunkter, men forskningen på udkig efter en biologisk markør for vinteren vækstdvale har intensiveret i de seneste år. De fleste forsøg på at finde en utvetydig indikator viser Hvornår et bud har brudt vækstdvale har dog været mislykket3. Den metode beskrevet heri, kombinere histokemiske metoder18 med analyse billede, har været meget nyttigt at undersøge forholdet mellem kulhydrat reserver af en bestemt væv og dens fysiologiske aktivitet i søde kirsebær knopper under de forskellige faser i vækstdvale5 og kan også anvendes til andre arter og væv10,13,20.
Kulhydrat reserver i form af stivelse spiller en vigtig rolle både i blomst udvikling og den reproduktive proces7,10,13,20,21,23 og i sæsonbetinget i de tempererede woody stauder6. Forskellige undersøgelser på vækstdvale har betalt opmærksomhed til stivelse i knopper24. Men på grund af deres lille størrelse, flere hele knopper er nødvendige for anvendelsen af kvantitative analysemetoder, og påvisning af kvantitative variationer i bestemte væv eller celler er begrænset af maskering effekten af de omkringliggende celler. Kombinationen af histokemiske metoder med billede analysesystem tilbyder en god lejlighed til at studere ændringer i forskellige strukturer inde i opløbet stivelsesindhold.
Denne metode har begrænsning af forebyggelse kvantificering af det nøjagtige indhold af stivelse i vævet men tillader relative stivelsesindholdet til kvantificeres at følge kvantitative stivelse ændringer over tid25 og sammenligne stivelsen indhold forskellige væv13,20, kultivarer eller år5. For at tillade den korrekte sammenligning af værdier af optiske densitet mellem felter, væv og knopper, kalibrering af systemet (lysforhold, farvning intensitet og forstørrelse) og indstilling af farve tærskler skal fastsættes præcist , gemmes, og bruges til alle forberedelserne.
Stivelse farvning og kvantificering baseret på kalium jod giver mulighed for den efterfølgende anvendelse af andre pletter efter vask af afsnittet. Således forskellige analyser kan udføres uden yderligere forberedelser og syntetiske montering medier ikke er brugt5,20. Ligeledes kan morfometrisk målinger26 ske efter de samme forberedelser, så mønstret af stivelse ophobning at være indrammet i forhold til væksten i forskellige strukturer5,13. Metoden kan tilpasses andre strukturer eller arter med tilpasningen af den farveniveauer, at bruger analyseværktøjet til påvisning af stivelse, som kan give mulighed for undersøgelse af andre udviklingsmæssige processer, der involverer ændringer i stivelsesindhold i små grupper af celler.
Forholdet mellem vækstdvale frigivelse og stivelse ophobning i æggestokken primordia afsløret ved brug af denne metode giver et solidt grundlag for at forstå biologiske grundlag for vækstdvale og isnende krav5. Dog kan stivelse kvantificering af billedanalyse på paraffin-embedded sektioner vise sig for at være meget besværligt og tidskrævende at vurdere kravene til nedkøling af et stort antal sorter. Fremtidig indsats skal være fokuseret på at studere pålidelige biologiske indikatorer, der kan nemt angive vækstdvale status af træet. I mellemtiden, mønster af stivelse variationer i vækstdvale kan bruges til at indramme yderligere fysiologiske og genetiske undersøgelser, og kombinationen af histokemiske metoder med billedanalyse beskrevet heri kunne bruges i andre træagtige flerårige afgrøder til kvantificere forskellige væv i forbindelse med vækstdvale stivelsesindhold.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ikke noget at oplyse.
Acknowledgments
Forfatterne tak taknemmeligt Maria Herrero og Eliseo Rivas for deres nyttige diskussion og rådgivning. Dette arbejde blev støttet af Ministerio de Economía y Competitividad — Europæiske Regionaludviklingsfond, EU [tilskud antal BES-2010-037992 til E. F.]; Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria [grant numre RFP2015-00015-00, RTA2014-00085-00, RTA2017-00003-00]; og Gobierno de Aragón — Europæiske Socialfond, EU [Grupo Consolidado A12-17R].
Materials
| Name | Company | Catalog Number | Comments |
| Precision scale | Sartorius | CP225D | |
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Drying-stove | Memmert | U15 | |
| Paraffin Embedding station | Leica Microsystems | EG1140H | |
| Rotatory microtome | Reichert-Jung | 1130/Biocut | |
| Microtome blade | Feather | S35 | Stainless steel |
| Bright field microscope | Leica Microsystems | DM2500 | |
| Digital Camera | Leica Microsystems | DC-300 | |
| Image Analysis System | Leica Microsystems | Quantiment Q550 |
References
- Kurokura, T., Mimida, N., Battey, N. H., Hytönen, T. The regulation of seasonal flowering in the Rosaceae. Journal of Experimental Botany. 64 (14), 4131-4141 (2013).
- Rohde, A., Bhalerao, R. P.
- Fadón, E., Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environmental and Experimental Botany. 152, 28-36 (2018).
- Fadón, E., Rodrigo, J., Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees - Structure and Function. , In Press (2018).
- Fadón, E., Herrero, M., Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Frontiers in Plant Science. 9 (171), (2018).
- Loescher, W. H., Mccamant, T., Keller, J. D. Carbohydrate reserves, translocation and storage in woody plant roots. HortScience. 25 (3), 274-281 (1990).
- Hedhly, A., et al. Starch turnover and metabolism during flower and early embryo development. Plant Physiology. , (2016).
- Rodrigo, J., Hormaza, J. I., Herrero, M. Ovary starch reserves and flower development in apricot (Prunus armeniaca). Physiologia Plantarum. 108 (1), 35-41 (2000).
- Julian, C., Rodrigo, J., Herrero, M. Stamen development and winter dormancy in apricot (Prunus armeniaca). Annals of Botany. 108 (4), 617-625 (2011).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil starch reserves at anthesis correlate with final flower fate in avocado (Persea americana). PLoS One. 8 (10), e78467 (2013).
- Smith, A. M., Zeeman, S. C. Quantification of starch in plant tissues. Nature Protocols. 1 (3), 1342-1345 (2006).
- Eliceiri, K. W., et al.
- Rodrigo, J., Herrero, M. Influence of intraovular reserves on ovule fate in apricot (Prunus armeniaca L.). Sexual Plant Reproduction. 11, 86-93 (1998).
- Zhou, J., Spallek, T., Faulkner, C., Robatzek, S. CalloseMeasurer: A novel software solution to measure callose deposition and recognise spreading callose patterns. Plant Methods. 8 (1), (2012).
- Faulkner, C., et al. An automated quantitative image analysis tool for the identification of microtubule patterns in plants. Traffic. 18 (10), 683-693 (2017).
- Kuhn, B. F. Determination of starch in ovules of the sour cherry cv. "Stevnsbaer.". European Journal of Horticultural Science. 71 (3), 120-124 (2006).
- Johansen, D. A. Plant microtechnique. , McGraw-Hill. New York, NY. (1940).
- Ruzin, S. E. Plant microtechnique and microscopy. , Oxford University Press. New York, NY. (1999).
- Rodrigo, J., Rivas, E., Herrero, M. Starch determination in plant tissues using a computerized image analysis system. Physiologia Plantarum. 99 (1), 105-110 (1997).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Ovary starch reserves and pistil development in avocado (Persea americana). Physiologia Plantarum. 140 (4), 395-404 (2010).
- Suarez, C., Castro, A. J., Rapoport, H. F., Rodriguez-García, M. I. Morphological, histological and ultrastructural changes in the olive pistil during flowering. Sexual Plant Reproduction. 25, 133-146 (2012).
- Lang, G. A., Early, J. D., Martin, G. C., Darnell, R. L. Endodormancy, paradormancy, and ecodormancy - Physiological terminology and classification for dormancy research. HortScience. 22 (3), 371-377 (1987).
- Hedhly, A., Vogler, H., Eichenberger, C., Grossniklaus, U. Whole-mount clearing and staining of arabidopsis flower organs and siliques. Journal of Visualized Experiments. 2018 (134), 1-10 (2018).
- Kaufmann, H., Blanke, M. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. Journal of Plant Physiology. 218 (July), 1-5 (2017).
- Herrero, M., Dickinson, H. G. Pollen-pistil incompatibility in Petunia hybrida: changes in the pistil following compatible and incompatible intraspecific crosses. Journal of Cell Science. 36, 1-18 (1979).
- Carpenter, A. E., et al. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biology. 7 (10), R100 (2006).
