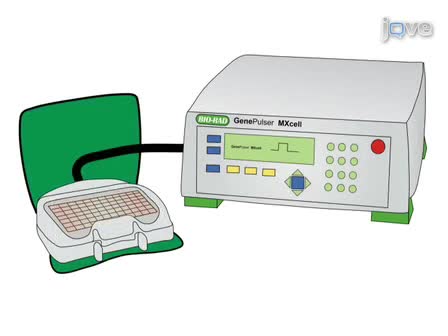
We describe a set of assays to analyze expression levels of H1 linker histones. mRNA of individual H1 genes are quantitatively measured by random primer based reverse transcription followed by real-time PCR, whereas protein quantification of H1 histones is achieved by HPLC analysis.