The ability to quantify birefringence of skeletal muscle provides a non-invasive but highly reproducible method to examine and compare levels of muscle damage, and examine muscle regeneration in vivo. Birefringence results from the diffraction of polarised light through the pseudo-crystalline array of the muscle sarcomeres15, and following injury or damage to the muscle, a reduction in birefringence is evident. Likewise, the activation and differentiation of stem cells results in the formation of new muscle fibres within the injury site, subsequently increasing birefringence intensity within this region. Using this system, we have examined muscle regeneration in a zebrafish model of congenital muscular dystrophy type 1A (MDC1A), caused by a deficiency in Lama216. A clutch of embryos from a cross between two lama2+/- zebrafish was collected, and at 3 dpf, the embryos were transferred to a DNA extraction chip (Figure 1A) and subsequently genotyped using an embryo genotyping technology. Having identified the genotype, lama2-/- larvae, which model MDC1A16, and lama2+/+ siblings were injured using a needle stab as per Figure 1B, and imaged on a polarizing microscope at 1 dpi, and 3 dpi, and the birefringence intensities were quantified. While muscle injury results in a reduction in birefringence intensity at 1 dpi (Figure 1Ci and Di), the successful regeneration of muscle results in increased birefringence in the same region (Figure 1Cii and Dii). It is also noteworthy that while lama2+/+ larvae display uniform birefringence intensity (Figure 1C), due to normal muscle patterning, the birefringence intensity in the muscle of lama2-/- was uneven and highly sporadic (Figure 1D), attributed to reduced muscle integrity.
Using this approach, we reveal that both wildtype larvae (lama2+/+; Figure 2A), and larvae deficient in lama2 (lama2-/-; Figure 2B), show significantly increased birefringence intensity in the wound site at 3 dpi compared to 1 dpi (Figure 2C), indicating that the muscle has regenerated. To compare the regenerative potential of larvae in each genotype, the regenerative index was determined, and we reveal that lama2-/- larvae displayed a striking increase in muscle regeneration compared to lama2+/+ larvae (Figure 2D; mean in lama2-/- = 1.30 ±- 0.251; mean in lama2-/- = 1.83 ± 0.439). To further validate the improved regeneration in lama2-/- larvae, we stained the muscle with an antibody against F-Actin (Supplementary Figure 1B-D). While these results confirm that lama2-/- do indeed regenerate, evident by the presence of differentiated muscle fibres within the wound site, the inability to examine the same fish at 1 dpi and 3 dpi limits the ability to quantify and compare the regenerative response between lama2+/+ and lama2-/- fish. Although the mechanistic basis for this improved regeneration capacity in lama2-/- larvae remains elusive, we believe that the loss of lama2 increases the number of activated stem cells, which subsequently results in improved muscle regeneration. However, further studies are needed to determine this. Collectively, these results highlight that ability of the described technique to identify changes in muscle regeneration in zebrafish models of muscle disease.

Figure 1: Overview of the genotyping and muscle regeneration protocol. (A) Image of a DNA extraction chip containing 24, 3 dpf zebrafish larvae. (B) Schematic of the orientation in which the 4 dpf larvae should be placed to perform the needle stab, with the head on the left, tail on the right, dorsal region up and ventral region down. The needle stab should be performed using a 30-gauge needle, targeting 1-2 somites of epaxial muscle. Created with BioRender.com. (C-D) Images of birefringence in a lama2+/+ and lama2-/- larvae at 1 dpi and 3 dpi. Regions shown in white and red reflect the areas used to quantify the birefringence in the wound site and uninjured somites respectively. Please click here to view a larger version of this figure.
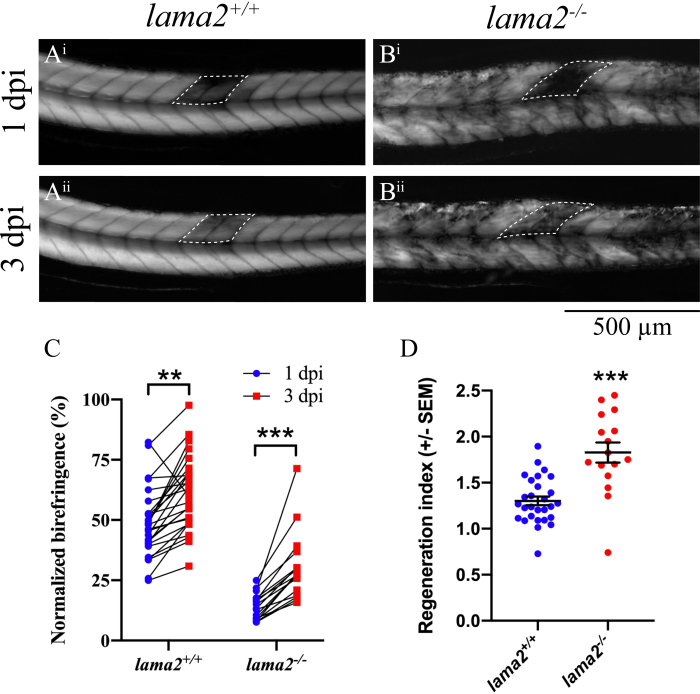
Figure 2: Quantification of muscle regeneration in lama2 deficient zebrafish larvae. Images of birefringence in lama2+/+ (A), and lama2-/- (B) larvae at 1 dpi and 3 dpi. The wound at 3 dpi in both, lama2+/+ and lama2-/- larvae is filled with new muscle. (C) Graph of the normalized birefringence of each larvae at 1 dpi and 3 dpi. The normalized birefringence in the wound site in lama2+/+ and lama2-/- larvae is significantly increased at 3dpi, as determined using a paired t-test. (D) Regenerative index in lama2+/+, and lama2-/- with the latter showing increased muscle regeneration, as determined using a t-test. Error bars represent SEM with larvae from three independent experiments (lama2+/+n=28, and lama2-/- n=16). Please click here to view a larger version of this figure.
Supplementary Figure 1: Examination of muscle regeneration in lama2 deficient larvae. Images of birefringence in lama2+/+ (Ai), and lama2-/- (Aii) larvae at 0 dpi demonstrating the presence of cellular debris within the wound site. Maximum projection confocal images of the larval myotome stained for F-actin at 0 dpi (B), 1 dpi (C) and 3 dpi (D). At 3 dpi, the wound site of lama2+/+ and lama2-/- larvae is characterized by the appearance of F-actin labelled muscle fibers. Please click here to download this file.
Supplementary Table 1: Template for the quantification of muscle regeneration. Please click here to download this file.
