

Summary
I denne undersøgelse blev en nano-mikrofluid flow kammer anvendes til at visualisere og funktionelt karakterisere twitching motiliteten af Xylella fastidiosa, en bakterie, der forårsager Pierce sygdom i vinstok.
Abstract
Xylella fastidiosa er en gram-negativ ikke-flagellated bakterie, der forårsager en række økonomisk vigtige sygdomme i planter. Den trækninger motilitet giver X. fastidiosa et middel til lange afstande inden anlægget bevægelse og kolonisering, der bidrager mod patogenicitet i X. fastidiosa. Den trækninger motilitet af X. fastidiosa drives af type IV pili. Type IV pili af Xylella fastidiosa reguleres af pilG, en kemotaksi regulator i Pil-CHP operon koder for proteiner, der er involveret med signaltransduktionsveje. For at belyse de roller pilG i twitching motilitet af X. fastidiosa, en pilG deficiente mutant Xf ΔpilG og dets komplementære stamme XfΔpilG- C indeholder indfødte pilG blev udviklet. Mikrofluidapparat kamre integreret med en tidsforskudt billedoptagelse system blev anvendt til at observere trækninger motilitet i XfΔpiLG, XfΔpilG- C og dets vildtype-stamme. Ved hjælp af denne optagelse system, det tillader langsigtede rumlige og tidslige observationer af sammenlægning, migration af individuelle celler og populationer af bakterier via trækninger motilitet. X. fastidiosa vildtype og supplerende XfΔpilG- C-stamme viste typiske trækninger motilitet egenskaber direkte observeret i de mikrofluide flow kamre, mens mutant XfΔpliG udstillet den trækninger mangelfuld fænotype. Denne undersøgelse viser, at pilG bidrager til twitching motiliteten af X. fastidiosa. Den mikrofluid flow kammeret anvendes som et middel til iagttagelse trækninger motilitet.
Introduction
Xylella fastidiosa er en Gram-negativ ikke-pisket, patogen bakterie, der forårsager en række økonomisk vigtige afgrøder sygdomme, herunder Pierce sygdom i vinstok (Vitis vinifera L.) 1,2, 3. Denne bakterie er begrænset til det vandledende xylem fartøjer. Infektion af Grapevine forårsager blokering af xylem skibe og resultater i vand stress og ernæringsmæssige mangler 3. Vellykket kolonisering afhænger af evnen af bakterien til at bevæge sig fra den oprindelige sted for infektion til resten af planten 3. Trækninger motilitet er et middel til flagellar-uafhængig bakteriel bevægelse gennem udvidelse, tilknytning, og tilbagetrækning af den polære type IV pili 4, som er blevet kendetegnet ved X. fastidiosa 5,6,7.
Den trækninger motilitet er blevet observeret af laser pincet og atomic force mikroskopi (AFM) 8,9,10. Ved anvendelse af disse teknikker, twitching motilities genereret af type IV pilus af N. gonorrhoeae og P. aeruginosa var præget af fl uorescently mærkning pili og erobre deres bevægelser mikroskopisk. Selv om begge metoder har detaljeret limen kraft enkelte bakterier, procedurerne er komplicerede og tidskrævende 9,10. Mikro fl uidic kamre blev anvendt til at observere langdistance migration af individuelle celler samt små aggregater af bakterieceller 5,6. Disse kamre blev udformet som en mikrofabrikeret-nano-kanal i en plade integreret med en tidsforskudt billede registreringssystem 11,12,13,14. Micro fl uidic kammer enheder tilbyder flere fordele for at studere bevægelse adfærd og celle-celle interaktioner af bakterier: (i) det giver en integreret platform med flere kanal kapaciteter; (Ii) det kan undersøge de bevægelser og aggregater af enkelte celler i nanoskala funktioner i bakterier; (Iii) det giver mulighed for direkte microscopic billedoptagelse af bakterieceller og time-lapse-analyse, (iv) det giver langsigtede rumlige og tidslige observationer af individuelle og / eller populationer af bakterier i en mikro-miljø; (V) strømningshastigheden dyrkningsmedium i en kanal kan styres præcist og (vi) kun en meget lille volumen (1 ml) af dyrkningsmedium er påkrævet for hvert eksperiment.
For nylig er mikro fl uidic fl ow systemet blevet anvendt til at undersøge adfærd af bakterieceller under forskellige mikromiljøer 14,15,16. Klæbeevnen og overfladen fastgørelse af E. coli 15, X. fastidiosa 16, og Acidovorax citrulli 14 til glasoverflader blev vurderet ved hjælp af mikro fl uidic kamre. Den sammenlægning og bio fi lm dannelse medieret af type IV pili af Acidovorax citrulli blev analyseret 14. Endvidere bevægelsen af A. citrulli observeret under fl ow cETINGELSER viste, at type IV pili kan spille vigtige roller i kolonisering og spredning af A. citrulli i xylem fartøjer under sap fl ow betingelser. De trækninger motilities af Pseudomonas aeruginosa og X. fastidiosa celler lykkedes observeret mod en flydende strøm i en mikrofabrikeret flow kammer 5,6,17. Type IV pilus deficient pilB og pilQ mutanter af X. fastidiosa blev fundet dybt ændre hastigheden af trækninger motilitet under de fl ow forhold i mikro fl uidic enheder 5,6,18. Undersøgelserne udført på bakteriel adhæsion og motilitet i mikro fl uidic enheder viste, at de mikro fl uidic kamre er særlig egnede til analyse af trækninger motilitet og migrering af pili-medieret bakterier in vitro. Disse resultater forklarer trækninger-medieret migration mekanisme, som letter celle-cellebinding, aggregering og kolonisering indenværten, med tiden føre til systemisk infektion.
Pil-CHP operon af X. fastidiosa indeholder pilG, Pili, pilJ, pille, chpB og chpC som koder signaltransduktionsveje 20. De transmembrane kemoreceptorer binder kemiske stimuli i det periplasmiske område og aktivere en signaleringskaskade i deres cytoplasmiske del til i sidste ende at styre bakteriel trækninger motilitet. I Pil-CHP operon af X. fastidiosa, en phospho-shuttieprotein PilG er en homolog til Chey. I E. coli og P. aeruginosa, Chey er responsregulatorprotein i kemotaksi systemer, der interagerer med flageller motordrevne proteiner 19, 21. Selvom bidragene fra Pil-CHP operon mod virulens i X. fastidiosa blev undersøgt for nylig 20, rolle pilG i kemotaksi operon som reaktion på de miljømæssige signaler og til regulerede / motor type IV pili af X. fastidiosa er uncLear. At belyse indsigt af kemotaksi regulator pilG i aktiviteten af trækninger motilitet X. fastidiosa, er en mikro fl uidic kammer anvendes til at vurdere trækninger motilitet af X. fastidiosa. Den pilG af X. fastidiosa er karakteriseret ved at sammenligne fænotyper af en sletning mutant Xf ΔpliG, supplerende belastning XfΔpliG C og dets vildtype in vitro. Resultaterne fremhæve rolle pilG i twitching motilitet af X. fastidiosa.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Perifere Fringe af bakteriel Colony
- Grow X. fastidiosa (Xf) Temecula vildtype 22, pilG deletionsmutant Xf ΔpliG (under anvendelse af tidligere beskrevne deletionsstrategi 23), og dets komplementære XfΔpliG -C (anvendelse af tidligere beskrevne kromosom-baseret genetisk komplementering strategi 24) på PD2 medium agarplader 25 ved 28 ° C i 5-7 dage.
- Autoklave cellofan (1 x 1 cm2) i vand ved 121 ° C (249 ° F) i 15 min. Saml ét stykke af cellofan, dræne vandet ved at berøre et hjørne af cellofan på en tom petriskål, omhyggeligt lægge cellofan over 15% af agaroverfladen og lufttørre.
- Pick up individuel X. fastidiosa kolonier med steril afrundede tandstikkere og spot-celler aseptisk onto en steriliseret ark af cellofan overlejret på 15% agar overflade i agarpladerne. Inkubér pladens ved 28 ° C i 2-3 dage.
- Undersøg kanten morfologi af kolonierne under anvendelse af et dissektionsmikroskop med en 2X objektivlinse og en 10X okular linse. Fotografere perifere frynser omkring kolonierne.
2. Mikroskopi og Mikrofluid Flow Chambers
- Fabrikere mikrofluidenheder brug af foto-litografiske procedurer svarende til dem, der tidligere er beskrevet 5,18. Design fire parallelle kanaler med computer-assisteret design software på mester silicium wafer ved hjælp af standard litografiske metoder 26.
- Opret mikrofluide kamre fra silicium wafer master med polydimethylsiloxan (PDMS). Hæld-polymeriserede PDMS over siliciumskiven master og helbrede den ved 60 ° C i 1 time. Skrælle PDMS replika fra wafer master og trim PDMS replika med en klinge i en 22 mm x 40 mm som den samme størrelse med et dækglas.
- Udsætte PDMS replika, et dækglas (22 x 40 mm 2), en Microscope slide (51 x 76 mm 2) til luft plasma ved 30 W i 2 min 27. Sandwich PDMS organ mellem dækglas og glasset mikroskop for at opbygge mikro fl uidic kammer.
- Bore et hul (5,5 mm diameter) gennem PDMS ved hver ende af den mønstrede-kanal. Skær silikonegummi slangen i 12-20 cm lange. Sæt den ene ende af siliconegummirør (5,1 mm udvendig diameter, 2,1 mm indvendig diameter, 0,8 mm wall) i hver åbning ende af kanalerne i PDMS replika, og forsegle det med upolymeriserede PDMS ved 60 ° C i 1 time.
- Tilslutte en anden ende af slangen til den med modhager forsynede ende af plast luer stik. Pak de forsamlede mikrofluide kamre med aluminiumsfolie og autoklaveres dem i 20 min.
- Indsamle bakterieceller af X. fastidiosa vildtype, mutant Xf ΔpliG, og supplerende XfΔpliG C via skrabning, hjælp engangs inokulerende sløjfer fra PD2 mellemstore plader. Juster celledensitettil en optisk densitet på 0,05 ved 600 nm i PD2 bouillon som tidligere 23 beskrevet. Saml bakteriecellen opløsning i en 1 ml gastæt sprøjte.
- Monter mikrofluidanordning på et omvendt mikroskop stadium. Tilslut et indgangsrør til 5 ml gastæt sprøjte indeholdende PD2 bouillon. Monter 5 ml gastæt sprøjte med sprøjtepumper.
- Tilslut udløbsrøret til et reservoir affald. Oprethold et medium strømningshastighed på 0,2 ul min -1 i 30 min for at stabilisere systemet.
- Tilslut sidegang indløbsrør til en 1 ml gastæt sprøjte indeholdende den bakterielle celle opløsning. Skyl bakteriecellen opløsningen gennem gummislanger indtil kanalen er nået. Oprethold et medium strømningshastighed på 0,2 ul min -1 i yderligere 30 minutter for at skylle ubundne celler fra kammeret før billedoptagelse.
- Monter mikroskopet lukkeren under field-justeret del af mikroskop for at styre lyset. Slut lukkeren til lukkeren control systemet og tilslut lukkeren styresystem til computeren.
- Monter et digitalt kamera til video-porten af mikroskopet og slutte den til computeren. Kør tidsforskudt optagelse software skal du vælge "shutter" funktionen fra menuen, og automatisk genkende den installerede lukkeren som standard i softwaren til at oprette forbindelser til lukkeren med softwaren.
- Vælg "digitalkamera" funktionen fra menuen af tidsforskudt optagelse software til automatisk at genkende det digitale kamera som standard capture enhed i softwaren og etablere kommunikation med det digitale kamera med softwaren.
- Find de bakterielle celler i en af kanalerne ved hjælp 20X fase kontrast optik, derefter skifte til 40X objektiv før billedoptagelse.
- Kør tidsforskudt optagelse software skal du vælge "erhvervelse billede" funktion ved hjælp af standardparametre fra menuen til at erhverve billederne fra mikroskop.Næste åbne funktionen "Acquire time-lapse" og indstille tidsintervallet til 30 sek 5,18,28 for varighed af 6-24 timer afhængig af eksperimentet er nødvendig for at observere trækninger motilitet af X. fastidiosa 5,18,28. Klik på "OK" for at starte tidsforskudt optagelse. Klik på "Stack-funktion" fra menuen, vælg "gem som" for at stable billederne i destinationsmappen på computeren efter endt optagelse.
- For flere kanaler, indfange time-lapse-billeder fra den første kanal hver 30 sek i 6 timer. Flyt objektivlinse af mikroskopet til den næste kanal at lokalisere målcellerne. Gentag time-lapse funktion som beskrevet ovenfor til at tage billeder i hver af fire kanaler sekventielt hvis forsøget er indstillet til at benytte fire kanaler. Fortsæt time-lapse billedoptagelse så længe som tre på hinanden følgende dage. Alle eksperimenter blev udført ved stuetemperatur (23 ± 2 ° C).
- Udfyld de time-lapse billeder i envideofil ved hjælp af tidsforskudt optagelse imaging software. Kør tidsforskudt optagelse software ved at klikke på "Stack-funktion" i menuen, og vælg "åben stack-funktion" for at åbne de stablede filer fra computeren.
- Start "Gør filmen" funktionen fra stakken modulet, vælge billeder og vælge "AVI" output format. Klik på "gem som" for at gemme videofil i destinationsmappen på computeren.
- Vælg de indsamlede film fra destinationsmappen på computeren og afspille dem. Derefter observere motilitet af de enkelte celler gennem den resulterende visualisering af trækninger motilitet aktivitet af bakterieceller i de genererede videofiler.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
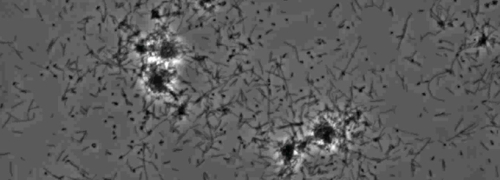
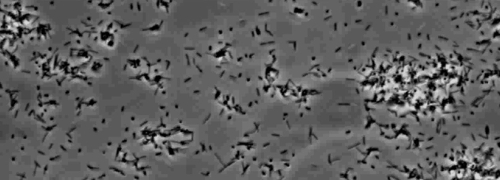
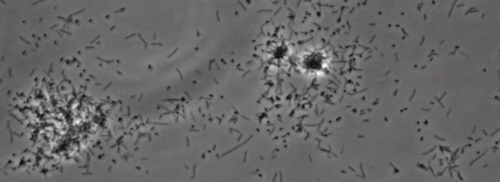
Tilstedeværelsen af en perifer koloni frynser indikerer type IV pilus-medieret trækninger motilitet, blev observeret i kolonier af X. fastidiosa vildtype og komplementære Xf ΔpliG -C stamme (figur 1). Mutant XfΔpliG imidlertid udviste ikke en bræmme omkring periferien af kolonierne (figur 1). Time-lapse billeddannelse af bakterieceller i nano-mikrofluid strømningskamre afslørede, at trækninger motilitet blev observeret i både vildtype X. fastidiosa og den supplerende XfΔpliG -C (Supplemental V1, V3), hvorimod XfΔpilG mutant celler ikke udviste trækninger motilitet hele eksperimentet (Supplemental V2). Celler af mutant XfΔpilG dannet relativt små løse aggregater i PD2 bouillon (Supplemental V2). I modsætning hertil celler af X. fastidiosa vildtype og supplerende XfΔpilG- C d eveloped større aggregater i PD2 bouillon (Figur 2, (Supplemental V1, V3).

Figur 1:. Den perifere yderkant bakteriekoloni Colony margin karakteristika X. fastidiosa fra vildtype, mutant Xf ΔpilG, og supplerende XfΔpilG- C dyrket på PD2 agar dækket med en steriliseret ark cellofan. Med undtagelse af mutant XfΔpilG, alle kolonier udviste en perifer yderkant, hvilket indikerer type IV pilus-medieret trækninger motilitet. Billeder blev taget efter 5 dages vækst på dyrkningsmediet. Forstørrelse bar, 0,5 mm. Klik her for at se en større version af dette tal.
_upload / 53816 / 53816fig2.jpg "/>
Figur 2: trækninger motilitet X. fastidiosa celler i nano-mikrofluid flow kammer. Det trækninger motilitet alle testede strain celler blev optaget under 6 dages observation. Vurderingerne blev gennemført fra tre uafhængige video segmenter. Forstørrelse bar, 20 pm.
Bemærk: trækninger motilitet X. fastidiosa celler er karakteriseret ved en enkelt celle bevægelighed på tværs glasoverflader gennem forlængelsen, tilknytning, og tilbagetrækning af den polære type IV pili. Den enkelt celle blev observeret i migrationen fortrinsvis mod en flydende strøm i en mikrofabrikeret flow kammer. Klik her for at se en større version af dette tal.
6 / 53816supfig1.jpg "/>
Supplerende Figur 1: En fire-kanals mikrofluid strømningskammeret. Hver kanal med medier i og medier ud stik i hver ende. Klik her for at se en større version af dette tal.
Supplerende Movie 1: Trækninger motilitet. (Højreklik for at downloade). Krampetrækning af vildtype X. fastidiosa i en mikrofluid flow kammer.
Supplerende Movie 2: Nedsat Twitching motilitet. (Højreklik for at downloade). Motilitet Xf mutant. XfΔpilG observeret i amicrofluidic strømningskammeret.
Supplerende Movie 3: Restaureret trækninger motilitet. (Højreklik for at downloade). Trækninger motilitet Xf komplementære stamme. XfΔpilG-C observeret i en mikrofluid flow kammer.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
I denne undersøgelse har vi kendetegnet bevægelse adfærd X. fastidiosa PilG mutant Xf ΔpilG og dens komplementære XfΔpilG- C stammer i nydesignede multiple parallel-nano-kanals mikro fl uidic kamre. De nydesignede mikro fl uidic kamre kan have op til fire parallelle kamre med 100 um nano-kanal i bredden i forhold til tidligere design med kun en enkelt 50 um bred kanal 18. Den forbedrede bredere nano-kanal letter indførelsen af bakterieceller med strømmende medier. Desuden denne mikrofluidik kammer er en) enkel at bygge og samle; 2) relativt billig; og 3) let anvendelse på forskellige eksperimentelle betingelser. Som et resultat, dette kammer design tillader langvarig observation af bevægelserne af de bakterielle celler under sorter eksperimentel mikromiljø.
Stabilisering af fl ow af strømmene through mikro fl uidic kanal er det kritiske trin til at skabe en intakt strømmende-mikro fl uidic mikromiljø til observation af motiliteten af bakteriecellerne under forskellige eksperimentelle miljøer. Træk af mikro fl uidic kamre og forbinder slangen med før indførelsen af bakterieceller medierne er også et vigtigt skridt til at stabilisere strømningen i kammeret. den høje hastighed af strømmen, vil imidlertid skylle bakteriecellerne ud af kammeret uden at bevare celler i løbet af kammeret. Den rette hastighed medierne flyder gennem mikro fl uidic kanaler skal justeres ved hjælp af en sprøjte pumpe. Under forsøgene i denne undersøgelse, den strømmende blev indstillet og stabiliseret ved pumpen ved 0,2 til 1 pi min -1 i mindst 30 minutter før indføring bakteriecellerne i kanalen. Når cellerne blev indført i mikro fl uidic kanaler, mediet fl ow blev holdt på 0,2 pi mi -1 i 30 til 60 minutter for at stabilisere systemet og fjerne nonattached celler. Det er meget vigtigt at stabilisere strømmen, og at holde baggrunden klart for at observere bevægelsen af bakteriecellerne. Billeder blev taget til fange hver 30 sek at bekræfte den trækninger motilitet aktiviteten af de indførte celler opretholder et konstant flow hastighed på 0,2 pi min -1. Hvis forsøgene kræver samling af de bakterielle celler i kammeret, kan cellerne skylles ud ved gradvist at øge de fl ow satser medier 0,2-110 pi min -1 ved at justere sprøjtepumpen hastighed.
De bakterielle celle aktiviteter i de mikro fl uidic kamre blev vurderet via inverteret mikroskop ved hjælp 40X fase kontrast optik og time-lapse billeder optaget med et digitalt kamera, som blev styret af imaging software. Strømmen hastighed og intervallet tid for billedoptagelse kan justeres i overensstemmelse hermed med experimEntal krav. Men i de fleste tilfælde, flow hastighed i et medium strømningshastighed er indstillet til 0,2 pi min -1 med time-lapse billeder optaget hver 30 sekunder ved pilus-medieret bakterieceller. I andre tilfælde, hvis mediet fl ow forøges til 0,1 pi min -1 under eksperimenteren adfærd af bakteriecellerne bliver optaget ved at opfange billeder hver 10 til 15 sek i overensstemmelse hermed. De time-lapse billeder blev taget hver 30 sek for varighed af 6-24 timer og gemt som en kilde billedfiler til hver testet stamme. Videoerne blev derefter fulgt fra billeder taget 6-8 timer fra hver stamme, som vist kontrast fænotyper i trækninger motilities mellem mutant og vildtype / supplere stammer (Supplemental V1, V2, V3).
Betydningen af nano-mikro fl uidic kammer enheder over makroskala parallel-plade fl ow kamre omfatter direkte undersøgelse af bevægelserne og samlinger af syngele celler af bakterier. Udover lave omkostninger 5 og lave reagens- og prøvevolumen krav, fordelene ved mikro fl uidic kamre er lette konstruktionen af et fl ow mikromiljø for bakteriekultur og nøjagtig kontrol over fl uid fl ow sats. De multiple parallelle kanaler tillader observation af differentielle bakteriestammer i et enkelt eksperiment setup, hvilket giver kompatible data til analyse. Den flagellar-uafhængige trækninger motilitet af bakterierne med den polære pili er særlig egnet til analyse i dette nano-mikro fl uidic kammer. Men denne mikro fl uidic kammer er mindre egnet til flagellar-afhængig bakteriel bevægelse, hvor bevægelsen af bakterier er normalt for hurtig og udviser tilfældige retninger. Denne begrænsning til tider kunne blive kompromitteret ved at justere medium flow til 0,05 pi min -1 og ændre time-lapse billedoptagelse hastighed hver 1 til 2 sek i 1 time for at ANALYze bevægelserne af flagel-medierede bakterier i nanoskala miljø.
Mikrofluide chamber anordninger, der anvendes heri tilvejebringer direkte visuel bevis for funktionel evaluering af PilG ansvarlig for motion adfærd in vitro. Desuden er denne undersøgelse viser også, at celle til celle sammenlægning gennem trækninger motilitet er afgørende for biofilmdannelse, signalering og patogenicitet i X. fastidiosa. Visualisering af bakterier trækninger motilitet ved mikrofluidenheder giver en ny tilgang til at studere genfunktion involverer adfærd i bakterier, der ikke er let måles ved andre analysemetoder. Denne fremgangsmåde kan anvendes på andre bakterielle systemer. De mikrofluide chamber enheder giver et strømningssystem til karakterisering den fysiologiske adfærd af bakterieceller associeret med celle-celle vedhæftede filer, celle aggregeringer, bevægelsesmønstre og biofilmdannelse.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne erklærer, at de ikke har nogen konkurrerende finansielle interesser.
Acknowledgments
Denne undersøgelse blev støttet af USA Department of Agriculture, Agricultural Research Service. Handel navne eller kommercielle produkter i denne publikation nævnes udelukkende med det formål at give specifikke oplysninger og indebærer ikke anbefaling eller godkendelse af United States Department of Agriculture. USDA er en lige muligheder udbyder og arbejdsgiver.
Materials
| Name | Company | Catalog Number | Comments |
| Biology materials | |||
| X. fastidiosa (Xf) Temecula wild type | Costa, H. S., et al., 2004 22 | ||
| pilG deletion mutant XfΔpliG | Shi, X. Y., et al., 2007 26 | ||
| pilG complementary strain XfΔpliG-C | Davis, M. J., et al. 1998 23 | ||
| Physical materials and equipment | |||
| Disposable inoculating loops | VWR international, Radnor, PA | #22-363-607 | quantitative procedures such as bacterial collection |
| Polydimethylsiloxane (PDMS) | Dow Corning Corporation | #0002709226 | Sylgard 184 silicone Elastomeric Kits |
| AmScope MD2000 digital camera | AmScope, Irvine, CA | SE305R-AZ-E | Image, video recording and measurement |
| Tubes line | Edgewood, NY | #T4300 | Connected to the syringe and microfluidic chamber |
| Plastic luer connectors | Edgewood, NY | Connected to the syringe and microfluidic chamber | |
| Syringe pumps | Pico Plus, Harvard Apparatus, MA | #702209 | The flow rate can be adjusted while the pump is running. |
| Syringes | Gastight, Hemilton Company, Reno, NV | #1005 | Provide the flowing broth |
| Inverted Olympus IMT-2 microscope | Olympus | IMT-2 FLuoro PHase | Image observation and recording |
| SPOT-RT digital camera | Diagnostic Instruments, Inc., MI | RT230 | Image, video recording and measurement |
| Microscope Shutter | The UNIBLITZ, US | #LS2T2 | Control camera’s exposure time |
| Microscope Shutter Control system | The UNIBLITZ, US | VCM-D1 | VCM-D1 Single Channel CE/UL/CSA Approved Shutter Driver |
| MetaMorph Image software | Universal Imaging Corp., PA | Real-time super-resolution image processing |
References
- Purcell, A. H., Hopkins, D. L. Fastidious xylem-limited bacterial plant pathogens. Annu. Rev. Phytopathol. 34, 131-151 (1996).
- Purcell, A. H. Xylella fastidiosa, a regional problem or global threat. J. Plant Pathology. 79, 99-105 (1997).
- Hopkins, D. L. Xylella fastidiosa: Xylem-limited bacterial pathogen of plants. Annu. Rev. Phytopathol. 27, 271-290 (1989).
- Mattick, J. S. Type IV pili and twitching motility. Annu. Rev. Microbiol. 56, 289-314 (2002).
- Meng, Y., et al. Upstream migration of Xylella fastidiosa via pilus-driven twitching motility. J. Bacteriol. 187, 5560-5567 (2005).
- Li, Y., et al. Type I and type IV pili of Xylella fastidiosa affect twitching motility, biofilm formation and cell-cell aggregation. Microbiology. 153, 719-726 (2007).
- Simpson, A. J. G., et al. The genome sequence of the plant pathogen Xylella fastidiosa. Nature. 406, 151-157 (2000).
- Maier, B., Potter, L., So, M., Long, C. D., Seifert, H. S., Sheetz, M. P. Single pilus motor forces exceed 100 pN. Proc. Natl. Acad. Sci. USA. 99, 16012-16017 (2002).
- Touhami, A., Jericho, M. H., Boyd, J. M., Beveridge, T. J. Nanoscale characterization and determination of adhesion forces of Pseudomonas aeruginosa pili by using atomic force microscopy. J. Bacteriol. 188, 370-377 (2006).
- Skerker, J. M., Berg, H. C. Direct observation of extension and retraction of type IV pili. Proc. Natl. Acad. Sci. USA. 98, 6901-6904 (2001).
- Brown, D. C., Larson, R. S. Improvements to parallel plate flow chambers to reduce reagent and cellular requirements. BMC Immunol. 2, 9 (2001).
- Thomas, W. E., Nilsson, L. M., Forero, M., Sokurenko, E. V., Vogel, V. Shear-dependent 'stick-and-roll' adhesion of type 1 fimbriated Escherichia coli. Mol. Microbiol. 53, 1545-1557 (2004).
- Thomas, W. E., Trintchina, E., Forero, M., Vogel, V., Sokurenko, E. V. Bacterial adhesion to target cells enhanced by shear force. Cell. 109, 913-923 (2002).
- Bahar, O., Fuente, D. L., Burdman, S. Assessing adhesion, biofilm formation and motility of Acidovorax citrulli using microfluidic flow chambers. FEMS Microbiol. Lett. 312, 33-39 (2010).
- Thomas, W. E. Using a laminar flow system to explain shear-enhanced bacterial adhesion. Proceedings of ICMM2005, Third International Conference on Microchannels and Mini-channels. Toronto, Ontario, Canada, , 751-759 (2005).
- Fuente, D. L., et al. Assessing adhesion forces of type I and type IV pili of Xylella fastidiosa bacteria by use of a microfluidic flow chamber. Appl. Environ. Microbiol. 73, 2690-2696 (2007).
- DeLange, P. A., Collins, T. L., Pierce, G. E., Robinson, J. B. PilJ localizes to cell poles and is required for type IV pilus extension in Pseudomonas aeruginosa. Curr Microbiol. 55, 389-395 (2007).
- Fuente, D. L., Burr, T. J., Hoch, H. C. Mutations in type I and type IV pilus biosynthetic genes affect twitching motility rates in Xylella fastidiosa. J. Bacteriol. 189, 7507-7510 (2007).
- Ferandez, A., Hawkins, A. C., Summerfield, D. T., Harwood, C. S. Cluster II che genes from Pseudomonas aeruginosa are required for an optimal chemotactic response. J. Bacteriol. 184, 4374-4383 (2002).
- Cursino, L., et al. Identification of an Operon, Pil-Chp, That Controls Twitching Motility and Virulence in Xylella fastidiosa. Mol. Plant Microbe Interact. 10, 1198-1206 (2011).
- Hazelbauer, G. L., Falke, J. J., Parkinson, J. S. Bacterial chemoreceptors: High-performance signaling in networked arrays. Trends Biochem. Sci. 33, 9-19 (2008).
- Costa, H. S., et al. Plant hosts of Xylella fastidiosa in and near southern California vineyards. Plant Dis. 88, 1255-1261 (2004).
- Shi, X. Y., Dumenyo, C. K., Hernandez-Martinez, R., Azad, H., Cooksey, D. A. Characterization of regulatory pathways in Xylella fastidiosa: genes and phenotypes controlled by algU. Appl. Environ. Microbiol. 73, 6748-6756 (2007).
- Matsumoto, A., Young, G. M., Igo, M. M. Chromosome-Based Genetic Complementation System for Xylella fastidiosa. Appl. Environ. Microbiol. 75, 1679-1687 (2009).
- Davis, M. J., Purcell, A. H., Thomson, S. V. Isolation Media for the Pierce's Disease Bacterium. Phytopathology. 70, 425-429 (1980).
- Xia, Y. N., Whitesides, G. M.
- Chaudhury, M. K., Whitesides, G. M. Direct measurement of interfacial interactions between semispherical lenses and flat sheets of poly-(dimethylsiloxane) and their chemical derivatives. Langmuir. 7, 1013-1025 (1991).
- Cruz, L. F., Parker, J. K., Cobine, P. A., De La Fuente, L. Calcium-enhanced twitching motility in Xylella fastidiosa is linked to a single PilY1 homolog. Appl. Environ. Microbiol. 80, 7176-7196 (2014).

{kind=link}