

Summary
I denne studien ble en nano-mikrofluidstrømning kammer ansatt for å visualisere og funksjonelt karakterisere rykninger motilitet av Xylella fastidiosa, en bakterie som forårsaker Pierce sykdom i Grapevine.
Abstract
Xylella fastidiosa er en Gram-negative ikke-flagellert bakterie som forårsaker en rekke økonomisk viktige sykdommer hos planter. Den rykninger motilitet gir X. fastidiosa et middel for langdistanse intra-anlegget bevegelse og kolonisering, bidra mot patogenitet i X. fastidiosa. Den rykninger motilitet av X. fastidiosa opereres av type IV pili. Type IV pili av Xylella fastidiosa er regulert av pilG, en chemotaxis regulator i Pil-Chp operon som koder for proteiner som er involvert med signaltransduksjonsveiene. Å belyse rollene pilG i rykninger motilitet av X. fastidiosa, en pilG -deficient mutant Xf ΔpilG og dens komplementære belastning XfΔpilG- C inneholder mors pilG ble utviklet. En microfluidic kamre integrert med en time-lapse bildeopptakssystem ble brukt til å observere rykninger bevegelighet i XfΔpIlg, XfΔpilG- C og dens villtypestammen. Ved hjelp av denne innspillingen systemet, tillater det langsiktige romlige og tidsmessige observasjoner av aggregering, migrasjon av individuelle celler og bestander av bakterier via rykninger motilitet. X. fastidiosa villtype og utfyllende XfΔpilG- C belastning viste typiske rykninger motilitet egenskaper direkte observert i microfluidic flyt kamre, mens mutant XfΔpliG utstilt rykninger mangel fenotype. Denne studien viser at pilG bidrar til rykninger motilitet av X. fastidiosa. Den mikrofluidstrømningskammeret anvendes som et middel for å observere rykninger motilitet.
Introduction
Xylella fastidiosa er en Gram-negative ikke-flagellert, sykdomsfremkallende bakterie som forårsaker en rekke økonomisk viktige avlingssykdommer, inkludert Pierce sykdom i Grapevine (Vitis vinifera L.) 1,2, 3. Denne bakterie er begrenset til vann-ledende Margen fartøy. Infeksjon av grapevine fører til blokkering av Xylem fartøy og resultater i vannstress og ernæringsmessige mangler 3. Vellykket koloniseringen er avhengig av evnen til bakterien til å bevege seg fra den initiale infeksjonsstedet til resten av anlegget 3. Rykninger bevegeligheten er et middel for å flagellær uavhengig bakteriell bevegelse gjennom utvidelsen, vedlegg, og tilbaketrekking av polar type IV pili 4 som er karakterisert ved X. fastidiosa 5,6,7.
Den rykninger bevegeligheten har blitt observert av laser pinsett og atomic force mikroskopi (AFM) 8,9,10. Ved å bruke disse teknikkene, tWitching motilities generert av type IV pilus av N. gonorrhoeae og P. aeruginosa ble preget av fl uorescently merking pili og fange deres bevegelser mikroskopisk. Selv om begge metoder har beskrevet adhesjonskraften av de enkelte bakterier, fremgangsmåten er komplisert og tidkrevende 9,10. Mikro fl uidic kamre ble anvendt for å observere lang avstand migrering av individuelle celler så vel som små aggregater av bakterieceller 5,6. Disse kamrene ble utformet som en microfabricated-nano-kanal i en plate integrert med en time-lapse bildeopptakssystem 11,12,13,14. Micro fl uidic kammer enheter tilbyr flere fordeler for å studere bevegelse atferd og celle-celle interaksjoner av bakterier: (i) det gir en integrert plattform med flere kanaler evner; (Ii) det kan undersøke bevegelser og samlinger av enkeltceller i nano-skala funksjoner bakterier; (Iii) det tillater direkte microscopic bildeopptak av bakterieceller og time-lapse-analyse, (iv) det gir langsiktige romlige og tidsmessige observasjoner av individuelle og / eller bestander av bakterier i en mikro-miljø; (V) strømningshastigheten av kulturmedium i en kanal kan kontrolleres nøyaktig og (vi) bare et meget lite volum (1 ml) av kulturmedium er nødvendig for hvert forsøk.
Nylig har mikro fl uidic fl ow system blitt ansatt for å undersøke atferd av bakterieceller under ulike microenvironments 14,15,16. Klebrighet og overflaten festing av E. coli 15, X. fastidiosa 16, og Acidovorax citrulli 14 til glassflater ble vurdert ved hjelp av mikro fl uidic kamre. Aggregering og bio fi lm dannelse mediert av type IV pili av Acidovorax citrulli ble analysert 14. Videre bevegelse av A. citrulli observert etter fl ow conditions vist at type IV pili kan spille viktige roller i koloniseringen og spredning av A. citrulli i Xylem fartøy i henhold saft fl ow forhold. De rykninger motilities av Pseudomonas aeruginosa og X. fastidiosa celler ble observert med hell mot en fluid strøm i et microfabricated strømningskammer 5,6,17. Type IV pilus mangel pilB og pilQ mutanter av X. fastidiosa ble funnet å dypt endre hastigheten på rykninger bevegelighet under de fl ow forholdene i mikro fl uidic enheter 5,6,18. De utført på bakteriell adhesjon og motilitet i mikro fl uidic enheter studier indikerte at mikro fl uidic kamre er spesielt egnet for analyse av rykning motilitet og migrering av pili-mediert bakterier in vitro. Disse resultatene forklare rykninger-mediert migrasjon mekanisme som muliggjør celle-celle vedlegg, aggregering og kolonisering innenforverten, til slutt føre til systemisk infeksjon.
Pil-Chp operon av X. fastidiosa inneholder pilG, pili, pilJ, pille, chpB og chpC som koder signaltransduksjonsveiene 20. De trans chemoreceptors binde kjemiske stimuli i periplasmatiske domene og aktivere en signalkaskade i sin cytoplasmadelen til slutt kontrollere bakterie rykninger motilitet. I Pil-Chp operon av X. fastidiosa, er en fosfor-shuttle protein PilG en homolog til Chey. I E. coli og P. aeruginosa, er Chey responsen regulator i Chemotaxis systemer som samhandler med flag motor proteiner 19, 21. Selv om bidragene fra de Pil-Chp operon mot virulens i X. fastidiosa ble undersøkt nylig 20, rolle pilG i chemotaxis operon som respons på miljøsignaler og til kontrollert / motor type IV pili av X. fastidiosa er unclear. Å belyse innsikt chemotaxis regulator pilG i aktiviteten til rykninger motilitet av X. fastidiosa, er en mikro fl uidic kammer brukes til å vurdere rykninger motilitet av X. fastidiosa. Den pilG av X. fastidiosa kjennetegnes ved å sammenligne fenotyper av en sletting mutert Xf ΔpliG, utfyllende belastning XfΔpliG C og dens naturlig in vitro. Resultatene markere rollen pilG i rykninger motilitet av X. fastidiosa.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Peripheral Fringe av bakteriell Colony
- Grow X. fastidiosa (Xf) Temecula villtype 22, pilG delesjonsmutant Xf ΔpliG (ved hjelp av tidligere beskrevne slettestrategi 23), og dens komplementære XfΔpliG-C (ved bruk av tidligere beskrevne kromosom basert genetisk komplementering strategi 24) på PD2-mediumagarplater 25 ved 28 ° C i 5-7 dager.
- Autoklaven cellofan (1 x 1 cm 2) i vann ved 121 ° C (249 ° F) i 15 min. Plukke opp en del av cellofan, tømme vannet ved å trykke på ett hjørne av cellofan på en tom petriskål, forsiktig lå cellofan over 15% av agaroverflaten og lufttørk.
- Plukk opp enkelte X. fastidiosa kolonier med sterile runde tannpirkere og spot celler aseptisk mot en sterilisert ark av cellofan kledde på 15% av agaroverflaten i agar plater. Inkuber platens ved 28 ° C i 2-3 dager.
- Undersøk kanten morfologi av koloniene med en dissekere mikroskop med en 2X objektiv og en 10X okulær linse. Fotografere perifer utkant rundt koloniene.
2. mikroskopi og mikrofluid Flow Chambers
- Dikte microfluidic enheter ved hjelp av foto-litografisk lignende prosedyrer som beskrevet tidligere 5,18. Design fire parallelle kanaler med dataassistert design software på mester silisium wafer ved hjelp av standard litografiske metoder 26.
- Opprett microfluidic kamre fra silisium wafer master med polydimetylsiloksan (PDMS). Hell upolymeriserte PDMS over silikonplaten master og herde den ved 60 ° C i 1 time. Skrelle av PDMS kopi fra wafer master og trim PDMS replika med et blad inn i en 22 mm x 40 mm som samme størrelse på et dekkglass.
- Utsett PDMS kopi, et glass dekkglass (22 x 40 mm 2), og en mikroskopope sklie (51 x 76 mm 2) til luft plasma på 30 W i 2 min 27. Sandwich PDMS legemet mellom dekkglass og glass mikroskop for å bygge mikro fl uidic kammer.
- Bore et hull (5,5 mm diameter) gjennom PDMS ved hver ende av det mønstrede-kanalen. Skjær silikon gummi slangen inn 12-20 cm lang. Sette inn en ende av silikongummirør (5,1 mm utvendig diameter, 2,1 mm indre diameter, 0,8 mm vegg) i hver åpne ende av kanalene i PDMS kopi, og forsegle den med upolymeriserte PDMS ved 60 ° C i 1 time.
- Koble til en annen ende av slangen til pigg enden av plast luerkonnektorer. Pakk den sammensatte microfluidic kamre med aluminiumsfolie og autoklav dem i 20 min.
- Samle bakterieceller av X. fastidiosa villtype, mutant Xf ΔpliG, og utfyllende XfΔpliG Cs via skraping, bruk av engangs inoculating looper fra PD2 mellomplater. Juster celletetthettil en optisk densitet på 0,05 ved 600 nm i PD2 buljong som beskrevet tidligere 23. Samle de bakterielle celleoppløsning i en 1 ml gasstett sprøyte.
- Monter microfluidic enheten på en invertert mikroskop scenen. Koble en innløpsrøret til 5 ml gasstett sprøyte som inneholder PD2 buljong. Monter 5 ml gasstett sprøyte med sprøytepumper.
- Koble utløpsrøret til en avfallsreservoaret. Opprettholde et medium strømningshastighet på 0,2 mL min -1 i 30 min for å stabilisere systemet.
- Koble rørene side-innløpet til en 1 ml sprøyte inneholdende gasstett bakteriecelleløsning. Spyl bakteriecelleløsning gjennom gummislangen inntil kanalen er nådd. Opprettholde et medium strømningshastighet på 0,2 mL min -1 i ytterligere 30 minutter for å spyle-bundne cellene fra kammeret før bildeopptak.
- Monter mikroskop lukkeren under feltet justerte del av mikroskopet for å styre lyset. Koble lukkeren til lukkeren control systemet og koble lukkeren kontrollsystemet til datamaskinen.
- Monter et digitalt kamera til videoporten av mikroskopet og koble den til datamaskinen. Kjør time-lapse opptak programvare, velger du "lukker" funksjon fra menyen, og gjenkjenne automatisk installert skodde som standard i programvaren for å opprette forbindelser til lukkeren med programvaren.
- Velg "digitalkamera" funksjon fra menyen i intervallopptak programvare for å automatisk gjenkjenne det digitale kameraet som standard digitaliseringsenhet i programvaren og etablere kommunikasjon med det digitale kameraet med programvaren.
- Finn de bakterielle celler i en av kanalene som bruker 20X fase kontrast optikk, deretter bytte til 40X objektiv før bildet tas.
- Kjør time-lapse opptak programvare, velger du "image oppkjøpet" funksjon ved hjelp av standardparameterne fra menyen for å skaffe bildene fra mikroskopet.Deretter åpner funksjonen "Hent time-lapse" og sette tidsintervallet til 30 sek 5,18,28 for varigheten av 6-24 timer avhengig av eksperiment for å observere rykninger bevegelighet av X. fastidiosa 5,18,28. Klikk "OK" for å starte intervallopptak. Klikk "Stack funksjon" fra menyen, velg "lagre som" for å stable bildene i en mappe på datamaskinen etter endt innspilling.
- For flere kanaler, ta time-lapse bilder fra den første kanalen hvert 30 sek i 6 timer. Bevege objektivlinsen av mikroskopet til den neste kanal for å lokalisere målcellene. Gjenta time-lapse funksjon som beskrevet ovenfor for å ta bilder i hver av fire kanaler sekvensielt hvis forsøket er satt opp til å utnytte fire kanaler. Fortsett time-lapse bildeopptak for så lenge som tre påfølgende dager. Alle forsøkene ble utført ved romtemperatur (23 ± 2 ° C).
- Kompilere time-lapse bilder til envideofil med intervallopptak bildebehandlingsprogrammer. Run time-lapse opptak programvare, kan du klikke "Stack funksjon" fra menyen og velg "åpne stabelen funksjon" for å åpne stablet filer fra datamaskinen.
- Start "Make filmen" funksjon fra stabelen modul, velge alle bildene og velge "AVI" output format. Klikk på "lagre som" for å lagre videofilen i målmappen på datamaskinen.
- Velg de kompilerte filmer fra målmappen på datamaskinen og spille dem. Deretter observere bevegeligheten av de enkelte cellene gjennom den resulterende visualisering av rykninger motilitet aktivitet av bakterieceller i den genererte videofiler.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
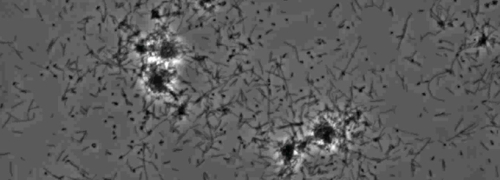
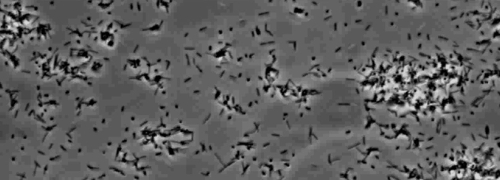
Tilstedeværelsen av en perifer koloni frynse indikativ for type IV pilus-mediert rykninger motilitet, ble observert i kolonier av X. fastidiosa villtype og komplementære Xf ΔpliG -C-stammen (figur 1). Mutant XfΔpliG, imidlertid ikke oppviser en ytterkant rundt periferien av koloniene (figur 1). Time-lapse avbildning av bakterieceller i nano-mikrofluidstrømnings kamre avdekket at rykninger motilitet ble observert i både villtype X. fastidiosa og utfyllende XfΔpliG C (Supplemental V1, V3), mens XfΔpilG mutante celler ikke vise rykninger bevegelighet gjennom hele forsøket (Supplemental V2). Celler av mutant XfΔpilG dannet relativt små løse aggregater i PD2 buljong (Supplemental V2). I kontrast, celler av X. fastidiosa villtype og utfyllende XfΔpilG- C d eveloped større aggregater i PD2 buljong (Figur 2, (Supplemental V1, V3).

Figur 1:. Den perifer utkant av bakteriekoloni Colony margin egenskapene til X. fastidiosa fra villtype, mutant Xf ΔpilG, og utfyllende XfΔpilG- C dyrket på PD2 agar dekket med en sterilisert ark av cellofan. Med unntak av mutant XfΔpilG, alle koloniene utviste en perifer frynser, som indikerer type IV pilus-mediert rykninger motilitet. Bilder ble tatt etter 5 dager med vekst på dyrkingsmedium. Forstørrelse bar, 0,5 mm. Klikk her for å se en større versjon av dette tallet.
_upload / 53816 / 53816fig2.jpg "/>
Figur 2: rykninger motilitet av X. fastidiosa celler i nano-mikrofluidstrømning kammeret. Den rykninger bevegelighet av alle testede belastningsskader celler ble spilt inn i løpet av 6 dager med observasjon. Vurderingene ble utført fra tre uavhengige videosegmentene. Forstørrelse bar, 20 mikrometer.
Merk: rykninger motilitet av X. fastidiosa celler er preget av enkelt celle bevegelse over glassflater gjennom utvidelsen, vedlegg, og tilbaketrekking av polar type IV pili. Den eneste celle ble observert i migrasjonsfortrinnsvis mot en væskestrøm i en microfabricated flyt kammer. Klikk her for å se en større versjon av dette tallet.
6 / 53816supfig1.jpg "/>
Supplerende Figur 1: En firekanals mikrofluidstrømningskammeret. Hver kanal med media i og media ut kontakter i hver ende. Klikk her for å se en større versjon av dette tallet.
Supplemental Movie 1: Rykninger motilitet. (Høyreklikk for å laste ned). Rykninger av vill type X. fastidiosa i et mikrofluidstrømning kammer.
Supplemental Movie 2: Nedsatt twitching motilitet. (Høyreklikk for å laste ned). Motilitet av Xf mutant. XfΔpilG observert i amicrofluidic flyt kammer.
Supplemental Movie 3: Restaurert rykninger motilitet. (Høyreklikk for å laste ned). Rykninger motilitet av Xf utfyllende belastning. XfΔpilG-C observert i et mikrofluidstrømningskammer.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
I denne studien, karakterisert vi bevegelse oppførselen til X. fastidiosa PilG mutant Xf ΔpilG og dets utfyllende XfΔpilG- C stammer i nydesignede flere parallell nano-kanals mikro fl uidic kamre. De nydesignede mikro fl uidic kamre kan ha opptil fire parallelle kamre med 100 mikrometer nano-kanal i bredden i forhold til tidligere design med bare en enkelt 50 mikrometer bred kanal 18. Den forbedrede bredere nano-kanal letter innføring av bakterieceller med rennende av mediet. I tillegg er denne MicroFluidics kammeret 1) enkelt å bygge og montere; 2) relativt billig; og 3) lett anvendelig for varierende eksperimentelle krav. Som et resultat av dette kammer konstruksjon tillater den langsiktige observasjon av bevegelser av bakteriecellene i henhold til varianter eksperimentelle mikromiljøet.
Stabilisere fl ow av strømmene igjennomgh mikro fl uidic kanal er det kritiske trinn for å skape et intakt strømmende-mikro fl uidic mikromiljø for observasjon av motiliteten av bakteriecellene under en rekke eksperimentelle omgivelser. Spylingen av mikro fl uidic kamre og som forbinder produksjonsrøret med media før innføringen av bakterieceller er også et viktig skritt for å stabilisere strømningen i kammeret. Imidlertid vil den høye hastigheten av strømnings spyle bakteriecellene ut av kammeret uten å beholde celler i løpet av kammeret. Den riktige hastigheten på media som strømmer gjennom mikro fl uidic kanaler må justeres ved hjelp av en sprøytepumpe. Under eksperimenter i denne studien var den strømmende satt og stabilisert ved hjelp av pumpen på 0,2 til 1 pl min -1 i minst 30 minutter før innføring av bakteriecellene inn i kanalen. Når cellene ble innført i mikro fl uidic kanaler, mediet strømning ble opprettholdt ved 0,2 ul mi -1 i 30 til 60 min for å stabilisere systemet og ta nonattached celler. Det er meget viktig for å stabilisere strømningen og for å holde den klar bakgrunn for å observere bevegelsen av bakteriecellene. Bilder ble tatt hvert 30 sek å lure fi rm rykninger motilitet aktiviteten av de innførte cellene opprettholde en konstant flyt hastighet på 0,2 mL min -1. Ved forsøkene krever samling av bakteriecellene i kammeret, kan cellene bli spylt ut ved gradvis å øke strømning ratene media 0,2 til 110 ul min -1 ved å justere sprøytepumpehastigheten.
Bakteriecellen aktiviteter i mikro fl uidic kamre ble vurdert gjennom invertert mikroskopet med 40X fase kontrast optikk og time-lapse bilder som er tatt med et digitalt kamera, som ble styrt av bildebehandlingsprogrammer. Flyten fart og intervallet tid for bildeopptak kan justeres tilsvarende med experimental krav. Men i de fleste tilfeller, strømningshastighet for et medium strømningshastigheten er innstilt på 0,2 mL min -1 med time-lapse bilder registreres hvert 30. sekund for de pilus-mediert bakterieceller. I andre tilfeller, hvis mediet fl ow hastigheten økes til 0,1 mL min -1 i løpet av eksperimentering, oppførselen til bakterieceller vil bli registrert ved å ta bilder hvert 10 til 15 sek tilsvarende. Time-lapse bildene ble tatt hver 30 sek varighet på 6-24 timer og lagret som en kilde bildefiler for hvert testet belastning. Videoene ble deretter overholdt fra bilder tatt 6-8 timer fra hver stamme, som vist kontrast fenotyper i rykninger motilities mellom mutant og villtype / utfylle stammer (Supplemental V1, V2, V3).
Betydningen av nano-mikro fl uidic kammer enheter over macroscale parallell-plate fl ow kamre omfatter direkte undersøkelse av bevegelser og samlinger av syngele celler av bakterier. I tillegg til lave kostnader 5 og lave reagens og prøve volum krav, fordelene av mikro fl uidic kamre er den enkle konstruksjonen av en fl ow mikromiljøet for bakteriekultur og nøyaktig kontroll over fl uid fl ow rate. De mange parallelle kanaler tillate observasjon av differensialbakteriestammer i et enkelt eksperiment-oppsett som gir kompatible data for analyse. Den flagell-uavhengig rykninger motilitet av bakterier med den polare pili er spesielt egnet for analyse i dette nano-mikro fl uidic kammer. Imidlertid er denne mikro fl uidic kammer mindre egnet til flagell-avhengige bakteriell bevegelse, hvor bevegelsen av bakterier er vanligvis altfor raskt og utviser tilfeldige retninger. Denne begrensningen noen ganger kan bli kompromittert ved å justere mellomstrømningshastigheten til 0,05 mL min -1 og endring av time-lapse bildefotografering hastigheten hver 1-2 sek for en time til ANALYZe bevegelsene til flagellpol-mediert bakterier i nano-skala miljø.
Microfluidic kammer enheter som brukes her gir direkte visuell bevis for en undersøkelse av PilG ansvarlig for bevegelse oppførsel in vitro. I tillegg denne studien viser også at celle til celle aggregering gjennom rykninger bevegelighet er viktig for biofilmdannelse, alarm og patogenitet i X. fastidiosa. Visualisering av bakterier rykninger motilitet ved microfluidic anordninger tilveiebringer en ny tilnærming for å studere genfunksjon som involverer oppførsel i bakterier som ikke er lett måles av andre analysemetoder. Denne tilnærmingen kan brukes for andre bakterielle systemer. De microfluidic kammer enheter gir et strømningssystem for å karakterisere den fysiologiske virkemåten av bakterieceller i forbindelse med celle-celle vedlegg, celle-aggregater, bevegelsesmønstre og biofilmdannelse.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne hevder at de ikke har noen konkurrerende økonomiske interesser.
Acknowledgments
Denne studien ble støttet av United States Department of Agriculture, Agricultural Research Service. Varemerker eller kommersielle produkter i denne publikasjonen er nevnt utelukkende for det formål å gi spesifikk informasjon og innebærer ikke en anbefaling eller godkjenning av United States Department of Agriculture. USDA er en lik mulighet leverandør og arbeidsgiver.
Materials
| Name | Company | Catalog Number | Comments |
| Biology materials | |||
| X. fastidiosa (Xf) Temecula wild type | Costa, H. S., et al., 2004 22 | ||
| pilG deletion mutant XfΔpliG | Shi, X. Y., et al., 2007 26 | ||
| pilG complementary strain XfΔpliG-C | Davis, M. J., et al. 1998 23 | ||
| Physical materials and equipment | |||
| Disposable inoculating loops | VWR international, Radnor, PA | #22-363-607 | quantitative procedures such as bacterial collection |
| Polydimethylsiloxane (PDMS) | Dow Corning Corporation | #0002709226 | Sylgard 184 silicone Elastomeric Kits |
| AmScope MD2000 digital camera | AmScope, Irvine, CA | SE305R-AZ-E | Image, video recording and measurement |
| Tubes line | Edgewood, NY | #T4300 | Connected to the syringe and microfluidic chamber |
| Plastic luer connectors | Edgewood, NY | Connected to the syringe and microfluidic chamber | |
| Syringe pumps | Pico Plus, Harvard Apparatus, MA | #702209 | The flow rate can be adjusted while the pump is running. |
| Syringes | Gastight, Hemilton Company, Reno, NV | #1005 | Provide the flowing broth |
| Inverted Olympus IMT-2 microscope | Olympus | IMT-2 FLuoro PHase | Image observation and recording |
| SPOT-RT digital camera | Diagnostic Instruments, Inc., MI | RT230 | Image, video recording and measurement |
| Microscope Shutter | The UNIBLITZ, US | #LS2T2 | Control camera’s exposure time |
| Microscope Shutter Control system | The UNIBLITZ, US | VCM-D1 | VCM-D1 Single Channel CE/UL/CSA Approved Shutter Driver |
| MetaMorph Image software | Universal Imaging Corp., PA | Real-time super-resolution image processing |
References
- Purcell, A. H., Hopkins, D. L. Fastidious xylem-limited bacterial plant pathogens. Annu. Rev. Phytopathol. 34, 131-151 (1996).
- Purcell, A. H. Xylella fastidiosa, a regional problem or global threat. J. Plant Pathology. 79, 99-105 (1997).
- Hopkins, D. L. Xylella fastidiosa: Xylem-limited bacterial pathogen of plants. Annu. Rev. Phytopathol. 27, 271-290 (1989).
- Mattick, J. S. Type IV pili and twitching motility. Annu. Rev. Microbiol. 56, 289-314 (2002).
- Meng, Y., et al. Upstream migration of Xylella fastidiosa via pilus-driven twitching motility. J. Bacteriol. 187, 5560-5567 (2005).
- Li, Y., et al. Type I and type IV pili of Xylella fastidiosa affect twitching motility, biofilm formation and cell-cell aggregation. Microbiology. 153, 719-726 (2007).
- Simpson, A. J. G., et al. The genome sequence of the plant pathogen Xylella fastidiosa. Nature. 406, 151-157 (2000).
- Maier, B., Potter, L., So, M., Long, C. D., Seifert, H. S., Sheetz, M. P. Single pilus motor forces exceed 100 pN. Proc. Natl. Acad. Sci. USA. 99, 16012-16017 (2002).
- Touhami, A., Jericho, M. H., Boyd, J. M., Beveridge, T. J. Nanoscale characterization and determination of adhesion forces of Pseudomonas aeruginosa pili by using atomic force microscopy. J. Bacteriol. 188, 370-377 (2006).
- Skerker, J. M., Berg, H. C. Direct observation of extension and retraction of type IV pili. Proc. Natl. Acad. Sci. USA. 98, 6901-6904 (2001).
- Brown, D. C., Larson, R. S. Improvements to parallel plate flow chambers to reduce reagent and cellular requirements. BMC Immunol. 2, 9 (2001).
- Thomas, W. E., Nilsson, L. M., Forero, M., Sokurenko, E. V., Vogel, V. Shear-dependent 'stick-and-roll' adhesion of type 1 fimbriated Escherichia coli. Mol. Microbiol. 53, 1545-1557 (2004).
- Thomas, W. E., Trintchina, E., Forero, M., Vogel, V., Sokurenko, E. V. Bacterial adhesion to target cells enhanced by shear force. Cell. 109, 913-923 (2002).
- Bahar, O., Fuente, D. L., Burdman, S. Assessing adhesion, biofilm formation and motility of Acidovorax citrulli using microfluidic flow chambers. FEMS Microbiol. Lett. 312, 33-39 (2010).
- Thomas, W. E. Using a laminar flow system to explain shear-enhanced bacterial adhesion. Proceedings of ICMM2005, Third International Conference on Microchannels and Mini-channels. Toronto, Ontario, Canada, , 751-759 (2005).
- Fuente, D. L., et al. Assessing adhesion forces of type I and type IV pili of Xylella fastidiosa bacteria by use of a microfluidic flow chamber. Appl. Environ. Microbiol. 73, 2690-2696 (2007).
- DeLange, P. A., Collins, T. L., Pierce, G. E., Robinson, J. B. PilJ localizes to cell poles and is required for type IV pilus extension in Pseudomonas aeruginosa. Curr Microbiol. 55, 389-395 (2007).
- Fuente, D. L., Burr, T. J., Hoch, H. C. Mutations in type I and type IV pilus biosynthetic genes affect twitching motility rates in Xylella fastidiosa. J. Bacteriol. 189, 7507-7510 (2007).
- Ferandez, A., Hawkins, A. C., Summerfield, D. T., Harwood, C. S. Cluster II che genes from Pseudomonas aeruginosa are required for an optimal chemotactic response. J. Bacteriol. 184, 4374-4383 (2002).
- Cursino, L., et al. Identification of an Operon, Pil-Chp, That Controls Twitching Motility and Virulence in Xylella fastidiosa. Mol. Plant Microbe Interact. 10, 1198-1206 (2011).
- Hazelbauer, G. L., Falke, J. J., Parkinson, J. S. Bacterial chemoreceptors: High-performance signaling in networked arrays. Trends Biochem. Sci. 33, 9-19 (2008).
- Costa, H. S., et al. Plant hosts of Xylella fastidiosa in and near southern California vineyards. Plant Dis. 88, 1255-1261 (2004).
- Shi, X. Y., Dumenyo, C. K., Hernandez-Martinez, R., Azad, H., Cooksey, D. A. Characterization of regulatory pathways in Xylella fastidiosa: genes and phenotypes controlled by algU. Appl. Environ. Microbiol. 73, 6748-6756 (2007).
- Matsumoto, A., Young, G. M., Igo, M. M. Chromosome-Based Genetic Complementation System for Xylella fastidiosa. Appl. Environ. Microbiol. 75, 1679-1687 (2009).
- Davis, M. J., Purcell, A. H., Thomson, S. V. Isolation Media for the Pierce's Disease Bacterium. Phytopathology. 70, 425-429 (1980).
- Xia, Y. N., Whitesides, G. M.
- Chaudhury, M. K., Whitesides, G. M. Direct measurement of interfacial interactions between semispherical lenses and flat sheets of poly-(dimethylsiloxane) and their chemical derivatives. Langmuir. 7, 1013-1025 (1991).
- Cruz, L. F., Parker, J. K., Cobine, P. A., De La Fuente, L. Calcium-enhanced twitching motility in Xylella fastidiosa is linked to a single PilY1 homolog. Appl. Environ. Microbiol. 80, 7176-7196 (2014).

{kind=link}