

Summary
本研究では、ナノマイクロ流体フローチャンバーは、視覚化し、機能的にXylella fastidiosa、グレイプバインのピアース病を引き起こす細菌の単収縮運動を特徴づけるために使用しました。
Abstract
Xylellaのfastidiosaは、植物の経済的に重要な多くの疾患の原因となるグラム陰性非鞭 毛細菌です。けいれん運動性は、Xを提供しますXの病原性に寄与する、長距離イントラ植物の動きと植民地化のための手段をfastidiosa fastidiosa。X.のけいれん運動fastidiosaは、IV型線毛によって運営されています。 XylellaのfastidiosaのIV型線毛はpilG、シグナル伝達経路に関与しているピル-Chpのオペロンをコードするタンパク質における走化性の調節因子によって調節されています。 Xの攣縮運動にpilGの役割を解明するために、 fastidiosa、pilG欠損変異体のXfΔpilGとネイティブpilGを含むその相補株XfΔpilG-Cを開発しました。タイムラプス画像記録システムと統合マイクロ流体チャンバはXfΔpに痙攣運動を観察するために使用されましたILG、XfΔpilG-Cおよびその野生型株。 この記録システムを使用して、長期の空間的および時間的な凝集の観察結果、個々の細胞および単収縮運動を介して細菌の集団の移動を可能にする。X.変異XfΔpliGがけいれん欠損表現型を示し、一方fastidiosa野生型および相補XfΔpilG-C株は、直接的にマイクロ流体フローチャンバーにおいて観察された典型的な単収縮運動の特性を示しました。この研究は、pilGは Xのけいれん運動性に寄与することを実証していますfastidiosa。マイクロ流体フローチャンバーは、単収縮運動性を観察するための手段として使用されます。
Introduction
Xylellaのfastidiosaはブドウでピアース病を含む、経済的に重要な作物の多数の疾患を引き起こすグラム陰性非鞭 毛、病原性細菌である( ヴィティスヴィニフェラ L.)1,2、3。この細菌は導水木部に限定されています船。ブドウの感染は、水ストレスや栄養不足3における木部船と結果の閉塞を引き起こします。成功植民地化は、植物3の残りの部分に感染の初期のサイトから移動する細菌の能力に依存します。運動性をけいれんすると、鞭毛に依存しない細菌の延長を経て動き、添付ファイル、およびXに特徴付けられている極性IV型線毛4の後退の手段でありますfastidiosa 5,6,7。
単収縮運動性は、レーザーピンセット及び原子間力顕微鏡(AFM)8,9,10により観察されました。これらの技術を用いて、TN.のIV型線毛によって生成された魔術の運動能淋菌およびP.緑膿菌は、FL uorescentlyラベリング線毛と微視的にその動きを捕捉することによって特徴づけられました。両方の方法は、個々の細菌の接着力を詳述しているが、手順が複雑で時間9,10を消費しています。微小流体チャンバは、長距離個 々の細胞の遊走、ならびに細菌細胞5,6の小さな凝集物を観察するために使用しました。これらのチャンバは、タイムラプス画像記録システム11,12,13,14と統合プレートに微細加工・ナノチャンネルとして設計されました。マイクロuidic室デバイスが移動行動や細菌の細胞間相互作用を研究するためのいくつかの利点を提供FL:(ⅰ)それは、複数のチャネル機能を備えた統合プラットフォームを提供します。 (ⅱ)それは細菌のナノスケールの特徴における単一細胞の運動や集計を調べることができます。 (ⅲ)それが直接メートルを可能にしますicroscopic細菌細胞の画像記録とタイムラプス解析、(iv)は、それは微小環境中の細菌の個々のおよび/または集団の長期的、空間的および時間的な観測を提供します。 (V)チャネルにおける培地の流量を正確に制御することができ、(VI)培地の非常に少量(1 ml)を各実験について必要とされます。
最近では、微小流体流動システムは、様々な微小環境14,15,16の下で細菌細胞の挙動を調査するために採用されています。接着性およびE.の表面付着大腸菌 15、X. fastidiosa 16、およびアシドボラックスは、ガラス表面への14の微小流体チャンバーを用いて評価したcitrulli。 アシドボラックスcitrulliのIV型線毛によって媒介凝集し、バイオフィルムの形成は、14を分析しました。 A.のさらに、運動流Cの下で観察citrullionditionsは、IV型線毛は、Aの植民地化と普及に重要な役割を果たしていることを実証しました樹液流条件下での木部船でcitrulli。 緑膿菌およびXのけいれんの運動能fastidiosa細胞が正常に微細加工フローチャンバー5,6,17内の流体流れに逆らって観察しました。 IV型線毛は、XのpilBとpilQ変異体を欠損しましたfastidiosaは深く微小流体デバイス5,6,18における流動条件でけいれん運動の速度を変更することが見出されました。微小流体デバイスにおける細菌の接着および運動性について行った研究は、微小流体チャンバが、インビトロで線毛媒介細菌の単収縮運動性および遊走を分析するのに特に適していることが示されました。これらの結果は、内の細胞 - 細胞付着、凝集・定着を促進するけいれん媒介移行メカニズムを説明しますホストは、最終的には全身感染につながります。
XのPIL-のCHPオペロンfastidiosaは、エンコード信号の伝達が20を経路pilG、線毛、pilJ、ピル、chpBとchpCが含まれています 。膜貫通化学受容体は、ペリプラズムドメイン内の化学的刺激を結合して、最終的には細菌のけいれんの運動性を制御するために、それらの細胞質部分のシグナル伝達カスケードを活性化します。 Xのピル-のCHPオペロン中fastidiosa、ホスホシャトルタンパク質PilGはチェイに相同体です。 E.でcoliおよびP.緑膿菌、崔はべん毛モータータンパク質19、21と相互作用する走化性システムのレスポンスレギュレーターです。 Xの病原性に向けてピル-のCHPオペロンの貢献が、 fastidiosaは 、最近20の環境シグナルへとX.の規制/モータIV型線毛に応答した走化性オペロン中pilGの役割を調べましたfastidiosaは、UNCでありますリア。 Xの攣縮運動の活動に走化性レギュレータpilGの洞察を解明するために、 fastidiosa、微小流体チャンバーはXのけいれんの運動性を評価するために使用されますfastidiosa。 X.のfastidiosaのpilGは、 欠失変異体 のXfΔpliG、補完的な歪みXfΔpliG-Cおよびin vitroでその野生型の表現型を比較することを特徴としています。結果は、Xのけいれん運動中pilGの役割を強調表示しますfastidiosa。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1.細菌コロニーの周辺フリンジ
- Xを育てます28℃でfastidiosa( のXf)テメキュラ野生型22、pilG欠失変異体のXfΔpliG(前述削除戦略23を使用)、およびその相補XfΔpliG-C(以前染色体ベースの遺伝的相補戦略24を用いて説明した)PD2培地寒天プレート上で25 5-7日のために、C。
- 15分間121°C(249°F)で水にオートクレーブセロファン(1×1cm 2の)。 、セロハンの1枚をピックアップ空のペトリ皿にセロハンの一角に触れて水を抜く、慎重に寒天表面と空気乾燥の15%の上にセロハンを築きます。
- 個々のXを拾いますfastidiosaコロニー滅菌丸い楊枝で寒天プレートに寒天表面の15%に重ねセロハンの滅菌シート上に無菌的に細胞を発見。プレートをインキュベート2〜3日間28℃でS。
- 2X対物レンズと10倍の接眼レンズを解剖顕微鏡を用いて、コロニーのエッジ形態を調べます。コロニーの周辺のフリンジを撮影。
2.顕微鏡やマイクロ流体フローチャンバー
- 以前5,18に記載したものと同様のフォトリソグラフィ手順を使用したマイクロ流体デバイスを製造します。標準的なリソグラフィ方法26を使用して、マスターのシリコンウェハ上のコンピュータ支援設計ソフトウェアを持つ4つの並列チャネルを設計します。
- ポリジメチルシロキサン(PDMS)でシリコンウェハマスターからマイクロ流体チャンバを作成します。シリコンウエハーマスター上に未重合PDMSを注いで、1時間60℃でそれを硬化させます。ウェハマスターからPDMSのレプリカをはがし、ガラスカバースリップの同じサイズとして22ミリメートルX 40ミリメートルにブレードとPDMSレプリカをトリミングします。
- PDMSのレプリカ、ガラスカバースリップ(22×40 mm 2)とし、microscを公開2分27用30 Wで空気プラズマにOPEスライド(51のx 76ミリメートル2)。サンドイッチガラスカバースリップと微小流体室を構築するためのガラス顕微鏡の間にPDMS本体。
- パターン化チャネルの各端部にPDMS貫通孔(5.5ミリメートル直径)ドリル。長い12〜20センチメートルにシリコーンゴムチューブをカットします。 PDMSレプリカのチャネルの各開口端部に、シリコーンゴム管(外径5.1ミリメートル、内径2.1 mmのを0.8mmの壁)の一端を挿入し、1時間60℃で重合していないPDMSとそれを密封します。
- プラスチック製のルアーコネクタのとげの終わりにチューブの他端を接続します。アルミホイルで組み立てられたマイクロ流体チャンバをラップし、20分のためにそれらをオートクレーブ。
- Xの細菌細胞を収集fastidiosa野生型、変異型のXfΔpliG、および相補XfΔpliG-C PD2培地プレートから使い捨て接種ループを使用して、掻き介して。細胞密度を調整しますPD2ブロス中の600nmで0.05の光学密度を、以前23記載されているように。 1ミリリットルガスタイトシリンジに細菌の細胞溶液を収集します。
- 倒立顕微鏡のステージ上にマイクロ流体デバイスをマウントします。 PD2ブロスを含む5ミリリットルガスタイトシリンジに導入管を接続します。シリンジポンプで5ミリリットルガスタイトシリンジを取り付けます。
- 廃棄物容器に排出管を接続します。 0.2μL分の媒体流量を維持-1で30分間、システムを安定化します。
- 細菌細胞溶液を含む1ミリリットルガスタイトシリンジに側入口管を接続します。チャネルに到達するまで、ゴム管を介して細菌の細胞溶液を洗い流します。チャンバーからの事前の画像キャプチャに結合していない細胞をフラッシュするために0.2μlの分の媒体流量-1別の30分間の維持。
- 光を制御するために、顕微鏡の視野-調整部分の下顕微鏡シャッターをマウントします。シャッター共同にシャッターを接続しますntrolシステムやコンピュータにシャッター制御システムを接続します。
- 顕微鏡のビデオポートにデジタルカメラをマウントし、それをコンピュータに接続します。タイムラプス撮影のソフトウェアを実行して、メニューから「シャッター」機能を選択して、ソフトウェアとシャッターへの接続を確立するためのソフトウェアでデフォルトとして自動的にインストールシャッターを認識しています。
- 自動的にソフトウェアのデフォルトのキャプチャデバイスとしてデジタルカメラを認識し、ソフトウェアとデジタルカメラとの通信を確立するためにタイムラプスレコーディングソフトウェアのメニューから「デジタルカメラ」機能を選択します。
- 20X位相差光学系を用いて、チャネルのいずれかで細菌細胞を見つけ、その後、画像キャプチャの前に40X対物レンズに切り替えます。
- 顕微鏡から画像を取得するには、メニューからデフォルトパラメータを使用して、「画像取得」機能を選択し、タイムラプス録画ソフトウェアを実行します。次に、「タイムラプスを取得」機能を開き、6-24時間は、Xのけいれん運動性を観察するために必要な実験に応じての持続時間30秒5,18,28に時間間隔を設定fastidiosa 5,18,28。タイムラプス撮影を開始するために「OK」をクリックします。メニューから「スタック機能」をクリックして、録音を終えた後、コンピュータ上の保存先フォルダ内の画像をスタックに「名前を付けて保存」を選択します。
- 複数のチャネルについては、6時間、最初のチャンネルから30秒ごとにタイムラプス画像をキャプチャします。標的細胞を見つけるために次のチャネルに顕微鏡の対物レンズを移動します。実験は4つのチャネルを利用するように設定されている場合は、4つのチャネルを順次それぞれの画像をキャプチャするために上記のようにタイムラプス機能を繰り返します。限り3として日間連続タイムラプス画像キャプチャを続けます。全ての実験は、室温(23±2℃)で行いました。
- タイムラプス画像をコンパイルしますタイムラプス撮影イメージングソフトウェアを使用して、ビデオファイル。タイムラプス撮影のソフトウェアを実行して、メニューから「スタック機能」をクリックして、コンピュータから積み重ねられたファイルを開くには、「オープンスタック機能」を選択します。
- すべての画像を選択し、「AVI」出力フォーマットを選択し、スタックモジュールから「映画を作る」機能を起動します。コンピュータ上の保存先フォルダ内のビデオファイルを保存するために「名前を付けて保存」をクリックします。
- コンピュータ上の保存先のフォルダからコンパイルされたムービーを選択し、それらを再生。そして、生成されたビデオファイルの中の細菌細胞の単収縮運動の活動の結果の可視化を通じて単一細胞の運動性を観察します。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
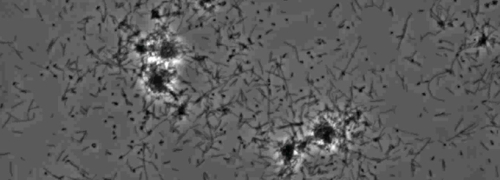
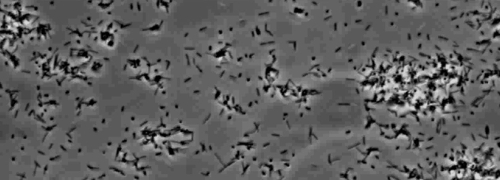
IV型線毛によって媒介される単収縮運動を示す周辺コロニー縞の存在は、Xのコロニーが観察されましたfastidiosa野生型および相補性のXfΔpliG-C株( 図1)。ミュータントXfΔpliGは 、しかし、コロニーの周囲( 図1)の周りにフリンジを示しませんでした。ナノマイクロ流体フローチャンバー内の細菌細胞のタイムラプスイメージングは、けいれん運動性は、両方の野生型X.で観察されたことを明らかにしましたfastidiosa相補XfΔpliG-C(補足V1、V3)、XfΔpilG変異細胞は、実験(補足V2)を通して運動性をけいれん示さなかったのに対し。変異XfΔpilGの細胞は、PD2ブロス(補足V2)に比較的小さな緩い凝集体を形成しました。 Xのとは対照的に、細胞fastidiosa野生型および相補XfΔpilG-C D PD2ブロス中のevelopedより大きな凝集体( 図2、(補足V1、V3)。

図1:細菌コロニーの周辺フリンジ Xのコロニーマージン特性。野生型、変異型のXfΔpilG、およびセロハンの滅菌シートで覆われたPD2寒天上で増殖させ、相補XfΔpilG-Cからfastidiosa。変異XfΔpilGを除いて、全てのコロニーは、IV型線毛媒介けいれん運動性を示す、周辺フリンジを示しました。写真は培養培地上での増殖の5日後に採取しました。倍率バー、0.5ミリメートル。 この図の拡大版をご覧になるにはこちらをクリックしてください。
_upload / 53816 / 53816fig2.jpg "/>
図2:Xのけいれん運動 ナノマイクロ流体フローチャンバー内 fastidiosa 細胞 。全ての試験された株細胞の単収縮運動は観察の6日間記録しました。評価は、3つの独立したビデオ・セグメントから行われました。倍率バーは20μm。
注意:Xのけいれん運動性をfastidiosa細胞を単一細胞拡張を介してガラス表面を横切る移動、結合、極性IV型線毛の退縮によって特徴付けられます。単一セルが微細加工フローチャンバー内の流体流れに逆らって優先的に移行が観察された。 この図の拡大版をご覧になるにはこちらをクリックしてください。
6 / 53816supfig1.jpg "/>
補足図1:4チャンネルマイクロ流体フローチャンバー 。各端の中でメディアおよびメディアアウトコネクタを持つ各チャネル。 この図の拡大版をご覧になるにはこちらをクリックしてください。
補足ムービー1:運動性をけいれん。 (右クリックしてダウンロード)。野生型X.のfastidiosaのけいれんをマイクロ流体フローチャンバーに。
補足ムービー2:減損トゥイッチングラムの運動性。 (右クリックしてダウンロード)。Xfの変異体の運動性を。 XfΔpilGはamicrofluidicフローチャンバー内で観察しました。
補足ムービー3:復元けいれん運動。 (右クリックしてダウンロード)。 Xfの補完的な株の運動性をけいれん。 XfΔpilG-Cは、マイクロ流体フローチャンバーで観察しました。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
本研究では、Xの動きの挙動を特徴付けfastidiosa PilG変異体のXfΔpilGと新たに設計された複数の並列-ナノチャネル微小流体チャンバ内のその相補XfΔpilG-C株。新たに設計された微小流体室は、単一の50μm幅のチャネル18と以前の設計に比べて幅が100μmのナノチャネルと最大4つの並列チャンバを持つことができます。改善され、より広いナノチャネルは、メディアが流れると細菌細胞の導入を容易にします。また、このマイクロ流体室は、構築し、組み立てるために1)簡単です。 2)比較的安価。 3)実験的な要件を変化させることに容易に適用できます。結果として、このチャンバ設計は、多様な実験微小環境下での細菌細胞の運動の長期的観察を可能にします。
電流throuのOW FLを安定させますGH微小流体チャネルは、実験環境の様々な環境下で細菌細胞の運動性の観察のための無傷の流入微小流体微小環境を作成するための重要なステップです。前の細菌細胞の導入にメディアとの微小流体室のフラッシングと接続するチューブは、チャンバ内の流れを安定させるための重要なステップです。しかし、流れの高速は、チャンバの過程で細胞を保持することなく、チャンバから細菌細胞をフラッシュします。微小流体チャネルを通って流れる媒体の適切な速度は、シリンジポンプを使用して調整する必要があります。本研究では実験中、流れるは、0.2〜1マイクロリットル分でポンプによって設定され、安定化した-1、少なくとも30分間、チャネルに細菌細胞を導入する前に。細胞は、微小流体チャネルに導入した後、媒体が流動0.2μlのメートルに維持しましたで-1 30〜60分間、システムを安定させ、非取付け細胞を除去します。流れを安定化させ、細菌細胞の動きを観察するために透明なバックグラウンドを維持することが非常に重要です。画像は、0.2μlの分で一定の流速を維持-1導入された細胞の単収縮運動の活動RM Fiを提供してコンために30秒毎に捕捉しました。実験は、チャンバ内の細菌細胞の収集を必要とする場合、細胞は徐々にシリンジポンプ速度を調整することにより、0.2μL分110 -1から媒体の流動速度を増加させることによって洗い流すことができます。
微小流体チャンバ内の細菌細胞活動は40X位相差光学系および画像化ソフトウェアによって制御されたデジタルカメラで記録されたタイムラプス画像を用いた倒立顕微鏡で評価しました。流速および画像キャプチャの時間間隔は、experimで適宜調整することができます内部の要件。媒体流率は0.2μlの分に設定されているためにしかし、ほとんどの場合、速度を流す-1タイムラプス画像と線毛媒介細菌細胞ごとに30秒を記録しました。他の例では、媒体であれば流率は0.1μL分に増加-1実験の過程の間に、細菌細胞の挙動は、それに応じて画像ごとに10〜15秒を捕捉することによって記録されます。タイムラプス画像は6-24時間の期間ごとに30秒を取り、試験した各菌株のためのソース画像ファイルとして保存されました。ビデオはその後、変異体および野生型/補完株(補足V1、V2、V3)の間の運動性をけいれんでコントラストの表現型を示し、各株から6-8時間から撮影した画像から従いました。
マクロスケールの平行平板流室以上のナノ微小流体チャンバーデバイスの意義は、動きの直接尋問と歌うの集計を含み細菌のル・セル。低コスト5と低い試薬およびサンプル容量の要件に加えて、微小流体チャンバの利点は、細菌培養および流体流速度に対する正確な制御のための流動微小環境の構築の容易されています。複数の並列チャネルは、分析のための互換性のあるデータを提供する単一の実験のセットアップに差動細菌株の観察を可能にします。極性線毛と細菌のべん毛に依存しないけいれん運動性は、このナノ微小流体室での分析のために特に適しています。しかし、この微小流体室は、細菌の運動は、通常、あまりにも迅速で、ランダムな方向性を示すに鞭毛依存細菌運動、にはあまり適していません。この制限は、時には0.05μLminに媒体の流量を調整することによって損なわ-1 ANALYに1時間毎に1〜2秒でタイムラプス画像キャプチャレートを変更することができナノスケール環境における鞭毛媒介細菌の動きをZE。
本明細書で使用されるマイクロ流体チャンバーデバイスは、in vitroでの運動行動に責任PilGの機能評価のための直接の視覚的証拠を提供します。さらに、この研究はまた、単収縮運動を介して細胞凝集細胞がXでシグナリングおよび病原性、バイオフィルム形成のために必須であることを明らかにfastidiosa。マイクロ流体デバイスによって運動性をけいれん細菌の可視化は、簡単に他のアッセイ法によって測定されない細菌中での挙動を伴う遺伝子機能を研究するための新しいアプローチを提供します。このアプローチは、他の細菌システムに適用することができます。マイクロ流体チャンバ装置は、細胞の添付、細胞集計、動きパターン及びバイオフィルム形成に関連した細菌細胞の生理的挙動を特徴づけるためのフローシステムを提供します。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
著者は、彼らが競合する金融利害関係を持たないことを宣言します。
Acknowledgments
この研究は、米国農務省、農業研究サービスによってサポートされていました。この資料に記載の商品名や商用製品は、特定の情報を提供する目的のためだけに記載されていると、米国農務省による推薦または保証を意味するものではありません。 USDAは機会均等プロバイダーおよび雇用主です。
Materials
| Name | Company | Catalog Number | Comments |
| Biology materials | |||
| X. fastidiosa (Xf) Temecula wild type | Costa, H. S., et al., 2004 22 | ||
| pilG deletion mutant XfΔpliG | Shi, X. Y., et al., 2007 26 | ||
| pilG complementary strain XfΔpliG-C | Davis, M. J., et al. 1998 23 | ||
| Physical materials and equipment | |||
| Disposable inoculating loops | VWR international, Radnor, PA | #22-363-607 | quantitative procedures such as bacterial collection |
| Polydimethylsiloxane (PDMS) | Dow Corning Corporation | #0002709226 | Sylgard 184 silicone Elastomeric Kits |
| AmScope MD2000 digital camera | AmScope, Irvine, CA | SE305R-AZ-E | Image, video recording and measurement |
| Tubes line | Edgewood, NY | #T4300 | Connected to the syringe and microfluidic chamber |
| Plastic luer connectors | Edgewood, NY | Connected to the syringe and microfluidic chamber | |
| Syringe pumps | Pico Plus, Harvard Apparatus, MA | #702209 | The flow rate can be adjusted while the pump is running. |
| Syringes | Gastight, Hemilton Company, Reno, NV | #1005 | Provide the flowing broth |
| Inverted Olympus IMT-2 microscope | Olympus | IMT-2 FLuoro PHase | Image observation and recording |
| SPOT-RT digital camera | Diagnostic Instruments, Inc., MI | RT230 | Image, video recording and measurement |
| Microscope Shutter | The UNIBLITZ, US | #LS2T2 | Control camera’s exposure time |
| Microscope Shutter Control system | The UNIBLITZ, US | VCM-D1 | VCM-D1 Single Channel CE/UL/CSA Approved Shutter Driver |
| MetaMorph Image software | Universal Imaging Corp., PA | Real-time super-resolution image processing |
References
- Purcell, A. H., Hopkins, D. L. Fastidious xylem-limited bacterial plant pathogens. Annu. Rev. Phytopathol. 34, 131-151 (1996).
- Purcell, A. H. Xylella fastidiosa, a regional problem or global threat. J. Plant Pathology. 79, 99-105 (1997).
- Hopkins, D. L. Xylella fastidiosa: Xylem-limited bacterial pathogen of plants. Annu. Rev. Phytopathol. 27, 271-290 (1989).
- Mattick, J. S. Type IV pili and twitching motility. Annu. Rev. Microbiol. 56, 289-314 (2002).
- Meng, Y., et al. Upstream migration of Xylella fastidiosa via pilus-driven twitching motility. J. Bacteriol. 187, 5560-5567 (2005).
- Li, Y., et al. Type I and type IV pili of Xylella fastidiosa affect twitching motility, biofilm formation and cell-cell aggregation. Microbiology. 153, 719-726 (2007).
- Simpson, A. J. G., et al. The genome sequence of the plant pathogen Xylella fastidiosa. Nature. 406, 151-157 (2000).
- Maier, B., Potter, L., So, M., Long, C. D., Seifert, H. S., Sheetz, M. P. Single pilus motor forces exceed 100 pN. Proc. Natl. Acad. Sci. USA. 99, 16012-16017 (2002).
- Touhami, A., Jericho, M. H., Boyd, J. M., Beveridge, T. J. Nanoscale characterization and determination of adhesion forces of Pseudomonas aeruginosa pili by using atomic force microscopy. J. Bacteriol. 188, 370-377 (2006).
- Skerker, J. M., Berg, H. C. Direct observation of extension and retraction of type IV pili. Proc. Natl. Acad. Sci. USA. 98, 6901-6904 (2001).
- Brown, D. C., Larson, R. S. Improvements to parallel plate flow chambers to reduce reagent and cellular requirements. BMC Immunol. 2, 9 (2001).
- Thomas, W. E., Nilsson, L. M., Forero, M., Sokurenko, E. V., Vogel, V. Shear-dependent 'stick-and-roll' adhesion of type 1 fimbriated Escherichia coli. Mol. Microbiol. 53, 1545-1557 (2004).
- Thomas, W. E., Trintchina, E., Forero, M., Vogel, V., Sokurenko, E. V. Bacterial adhesion to target cells enhanced by shear force. Cell. 109, 913-923 (2002).
- Bahar, O., Fuente, D. L., Burdman, S. Assessing adhesion, biofilm formation and motility of Acidovorax citrulli using microfluidic flow chambers. FEMS Microbiol. Lett. 312, 33-39 (2010).
- Thomas, W. E. Using a laminar flow system to explain shear-enhanced bacterial adhesion. Proceedings of ICMM2005, Third International Conference on Microchannels and Mini-channels. Toronto, Ontario, Canada, , 751-759 (2005).
- Fuente, D. L., et al. Assessing adhesion forces of type I and type IV pili of Xylella fastidiosa bacteria by use of a microfluidic flow chamber. Appl. Environ. Microbiol. 73, 2690-2696 (2007).
- DeLange, P. A., Collins, T. L., Pierce, G. E., Robinson, J. B. PilJ localizes to cell poles and is required for type IV pilus extension in Pseudomonas aeruginosa. Curr Microbiol. 55, 389-395 (2007).
- Fuente, D. L., Burr, T. J., Hoch, H. C. Mutations in type I and type IV pilus biosynthetic genes affect twitching motility rates in Xylella fastidiosa. J. Bacteriol. 189, 7507-7510 (2007).
- Ferandez, A., Hawkins, A. C., Summerfield, D. T., Harwood, C. S. Cluster II che genes from Pseudomonas aeruginosa are required for an optimal chemotactic response. J. Bacteriol. 184, 4374-4383 (2002).
- Cursino, L., et al. Identification of an Operon, Pil-Chp, That Controls Twitching Motility and Virulence in Xylella fastidiosa. Mol. Plant Microbe Interact. 10, 1198-1206 (2011).
- Hazelbauer, G. L., Falke, J. J., Parkinson, J. S. Bacterial chemoreceptors: High-performance signaling in networked arrays. Trends Biochem. Sci. 33, 9-19 (2008).
- Costa, H. S., et al. Plant hosts of Xylella fastidiosa in and near southern California vineyards. Plant Dis. 88, 1255-1261 (2004).
- Shi, X. Y., Dumenyo, C. K., Hernandez-Martinez, R., Azad, H., Cooksey, D. A. Characterization of regulatory pathways in Xylella fastidiosa: genes and phenotypes controlled by algU. Appl. Environ. Microbiol. 73, 6748-6756 (2007).
- Matsumoto, A., Young, G. M., Igo, M. M. Chromosome-Based Genetic Complementation System for Xylella fastidiosa. Appl. Environ. Microbiol. 75, 1679-1687 (2009).
- Davis, M. J., Purcell, A. H., Thomson, S. V. Isolation Media for the Pierce's Disease Bacterium. Phytopathology. 70, 425-429 (1980).
- Xia, Y. N., Whitesides, G. M.
- Chaudhury, M. K., Whitesides, G. M. Direct measurement of interfacial interactions between semispherical lenses and flat sheets of poly-(dimethylsiloxane) and their chemical derivatives. Langmuir. 7, 1013-1025 (1991).
- Cruz, L. F., Parker, J. K., Cobine, P. A., De La Fuente, L. Calcium-enhanced twitching motility in Xylella fastidiosa is linked to a single PilY1 homolog. Appl. Environ. Microbiol. 80, 7176-7196 (2014).

{kind=link}