

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
由于工具和方法不足,神经退行性变中的神经元-神经胶质相互作用尚未得到很好的理解。在这里,我们描述了从人类多能干细胞中获得诱导神经元,少突胶质细胞前体细胞和少突胶质细胞的优化方案,并提供这些方法在理解阿尔茨海默病细胞类型特异性贡献方面的价值示例。
Abstract
在阿尔茨海默病(AD)和其他神经退行性疾病中,少突胶质细胞衰竭是一种常见的早期病理特征,但它如何促进疾病的发展和进展,特别是在大脑的灰质中,仍然在很大程度上是未知的。少突胶质细胞谱系细胞的功能障碍以髓鞘形成缺陷和少突胶质细胞前体细胞(OPCs)自我更新受损为特征。这两种缺陷至少部分是由神经元和少突胶质细胞之间沿着病理学积累的相互作用的破坏引起的。OPC在中枢神经系统发育过程中产生髓鞘少突胶质细胞。在成熟的大脑皮层中,OPC是主要的增殖细胞(占总脑细胞的~5%),并以神经活动依赖性的方式控制新的髓磷脂形成。由于缺乏适当的工具,这种神经元到少突胶质细胞的通讯研究明显不足,特别是在AD等神经退行性疾病的背景下。近年来,我们的小组和其他人在改进目前可用的方案方面取得了重大进展,以从人类多能干细胞中单独生成功能性神经元和少突胶质细胞。在这篇手稿中,我们描述了我们的优化程序,包括建立一个共培养系统来模拟神经元 - 少突胶质细胞连接。我们的说明性结果表明,OPCs/少突胶质细胞对脑淀粉样变性和突触完整性做出了意想不到的贡献,并强调了该方法在AD研究中的实用性。这种还原论方法是从大脑内部固有的复杂性中剖析特定异细胞相互作用的有力工具。我们在这里描述的方案有望促进未来对神经变性发病机制中少突胶质细胞缺陷的研究。
Introduction
少突胶质细胞谱系细胞(包括少突胶质细胞前体细胞 (OPC)、髓鞘少突胶质细胞和介于两者之间的过渡型细胞)构成了人类脑细胞的主要群体1,它们积极参与许多关键功能,在整个神经发育和衰老过程中正常运作和维护我们的中枢神经系统2,3,4.虽然少突胶质细胞以产生髓磷脂以促进神经元活动传递并支持白质中的轴突健康而闻名,但OPC在髓鞘形成稀缺的灰质中含量丰富(~5%),并执行活动依赖性信号传导功能以控制学习行为和记忆形成5,6,7,8.少突胶质细胞在阿尔茨海默病(AD)和其他年龄相关神经退行性疾病的发病机制中的功能和功能障碍的研究不足9。适当的模型系统的不足和指导实验路径前进的一般知识的不足是造成这一差距的主要原因。
鉴于从多能干细胞(包括胚胎干(ES)和诱导多能干细胞(iPS)细胞)中提取人脑细胞的最新突破,这种细胞模型与现代基因编辑工具相结合已成为处理大脑中细胞相互作用的复杂联系的强大工具,并且能够证明人类特有的疾病表现10,11.考虑到单个脑细胞类型在面对相同的AD促进条件时可以表现出不同甚至相互矛盾的效果12,13,这种干细胞方法独特地提供了以前使用已建立的体内或体外模型错过的细胞类型特异性信息,这些模型仅提供来自脑细胞类型集合的汇总读数。在过去的十年中,已经开发了大量可靠的协议,以通过ES / iPS细胞的转分化或从其他终末分化细胞类型(例如成纤维细胞)的直接转化中产生人类神经元14,15。特别是,将关键的神经源性转录因子(例如,神经原蛋白2,Ngn2)16应用于人多能干细胞,可以为纯培养物产生表征良好的神经元细胞类型的均质群体,而无需与神经胶质细胞共培养12,17,18。对于诱导的人少突胶质细胞,有一些已发布的方案可以产生与其主要对应物高度相似的功能细胞,具有广泛的效率和时间和资源需求19,20,21,22,23,24,25,26,27,28.迄今为止,这些方案均未应用于研究少突胶质细胞如何响应和影响AD发病机制。
在这里,我们描述了我们改进的人类诱导神经元(iNs)和OPC /少突胶质细胞(iOPC / iOLs)的单一和混合培养方案。这里描述的iN协议基于广泛使用的Ngn2方法16,并且具有无胶质细胞的附加特征。所得iN是同质的,与皮质层2/3兴奋性神经元高度相似,具有特征性的锥体形态、基因表达模式和电生理特征17,18(图1)。为了克服多能干细胞定向分化的一些基本障碍,我们开发了一种简单有效的低剂量二甲基亚砜(DMSO)预处理方法29,30,并报告了人ES / iPS细胞转分化为iOPC和iOLs31的增强倾向,基于Douvaras和Fossati32广泛适应的方案.我们进一步简化了方案,并掺入了一种强大的分化促进化合物克立姆定7,33,34,以加速少突胶质细胞成熟过程。结果(图2),iOPCs可以在2周内产生(标记物O4的~95%阳性),iOLs可以在四周内产生(表达成熟的标志物MBP和PLP1)。有趣的是,我们发现单独iOPCs分泌大量淀粉样蛋白-β(Aβ),与独立的转录组学数据一致,数据显示淀粉样蛋白前体蛋白(APP)和加工蛋白酶β分泌酶(BACE1)在少突胶质细胞谱系细胞中的丰富表达35,36。此外,我们的iN-iOPC共培养系统通过MBP阳性iOL过程促进轴突的鞘化,并为突触形成提供重要支持(图3)。因此,我们在下面详细介绍的方案与先前编目的神经元-少突胶质共培养方法相比具有技术和生物学优势,并有望更好地模拟AD中的神经变性。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 人多能干细胞对人神经元的诱导
- 慢病毒制备(~5天,详细方案如前所述16)
- 在每个T75烧瓶中接种~100万个HEK293T细胞,使其在进行转染时~40%汇合。用表达四环素诱导型Ngn2和嘌呤霉素抗性基因(PuroR;在相同的TetO启动子对照下)、rtTA和三种辅助质粒pRSV-REV、pMDLg/pRRE和VSV-G(12μg慢病毒载体DNA和6μg辅助质粒DNA)的质粒转染它们。每种慢病毒制剂至少准备三个烧瓶。按照制造商的说明使用PEI进行转染。16小时后更换培养基并丢弃。
- 通过每天收集培养基来收获释放的病毒颗粒,并用新鲜培养基替换3天。汇集收集的含有病毒颗粒的培养基进行纯化。通过0.22μm过滤器过滤病毒,并以49,000× g 离心90分钟。将沉淀重悬于适当体积的PBS-葡萄糖(~150μL)中。
- 神经元诱导(~5天)
注意:该诱导方案(图1A;流程图)对于经过验证的多能性的iPS和ES细胞都非常有效(可以通过对表征良好的多能性标志物进行免疫组织化学染色来测定; 图 1B)。- 在52代时使用市售的H1人ES细胞(参见材料表)。使用ES细胞维持培养基(参见材料表)在包被的6孔板(每6孔板~0.5mg基质溶液;参见材料表)的细胞外基质溶液上培养细胞,并将板在37°C与5%CO2孵育。
- 在第-2天,用1mL细胞分离溶液(参见 材料表)分离ES细胞(80%汇合),并在室温下孵育10分钟。将细胞转移到试管中;用 2 mL 培养基清洗孔并在同一管中混合。以300× g 离心5分钟,将沉淀重悬于培养基中,并以每孔1 x 105 个细胞的接种密度将细胞接种到基质包被的6孔板上。
- 在第-1天,将表达Ngn2的慢病毒加上PuroR和rtTA与聚溴乙烯(8μg/ ml)一起添加到新鲜ES细胞维持培养基中的ES细胞中(参见 材料表)。病毒的确切数量应由实际滴度或滴定确定。我们通常在 6 孔板中每孔添加 5 μL 每种病毒。
- 在第 0 天,在 DMEM-F12 培养基中加入多西环素(2 μg/mL,以激活 Ngn2 表达),加入不含形态原的 N2 补充剂。
- 在第 1 天,在 DMEM-F12 加 N2 和多西环素的新鲜培养基中加入嘌呤霉素,至终浓度为 1 μg/mL 培养基。选择嘌呤霉素中的转导细胞至少24小时,如果病毒滴度低,可能需要更高的嘌呤霉素浓度(高达5μg/ mL)和更长的选择期(长达48小时)以充分去除转导不足的细胞。
- 在第2天,用细胞分离溶液分离分化神经元(见 材料表),并将它们重新铺在涂有基质溶液的24孔板(80,000-200,000个细胞/孔之间)上(见 材料表),并将它们保存在不含强力霉素的NBA / B27培养基中。播种密度至关重要。
- 在这个阶段,分离的神经元可以在专门的商业冷冻培养基中冷冻(见 材料表)并在液氮中储存长达3个月。纯神经元可以铺板,占解冻后典型的~15%-20%细胞死亡,单独培养或与其他脑细胞类型共培养(参见步骤3.2.3.与OPC共培养)。
- 按照制造商的说明,在涂有细胞外基质溶液的平板上培养纯iN(参见 材料表)。特征性锥体形态应在第 4 天(和第 6 天; 图 1C)。突触形成最早可在第 14 至 16 天检测到,并在第 24 天通过用标准突触前和突触后标志物进行免疫组织化学染色而突出。(图1D;用突触前标记物突触1和树突标记物Map2标记)。
2. 多能干细胞诱导人少突胶质细胞前体细胞(OPCs)和少突胶质细胞成熟
- 神经祖细胞(NPC)生成:单层方案(~7天)。流程图见 图2A 。
- 如前所述培养H1人ES细胞(参见步骤1.2.1.),并通过称为双SMADi的既定方法将其转分化为神经祖细胞(NPC),并具有用于多种信号通路的小分子抑制剂。在这里,我们使用广泛接受的商业试剂盒,并遵循制造商提供的单层协议(见 材料表)。
- 在第-1天,在涂有生长因子还原基质溶液(参见材料表;每6孔板~0.5mg基质溶液)的6孔板中板0.5–1 x 106个细胞,并用ES细胞维持培养基(参见材料表)。该生长因子还原基质溶液用于涂覆将在以下步骤中使用的所有板。
- 在第0天,用补充2%DMSO的ES细胞维持培养基(参见 材料表)处理细胞24小时。
- 在第1-6天,用含有商业试剂盒中SMAD抑制剂的温热(37°C)神经诱导培养基更换全培养基(见 材料表)。如果细胞在第7天之前分裂并达到汇合,则传代至0.5-1 x 106的接种密度,如前面的步骤2.1.2中所述。
- 在第7天,使用细胞分离溶液(参见 材料表)传代NPC,并以1-2×105个细胞 /孔的接种密度在24孔板中接种。
- 通过免疫组织化学 (IHC) 染色测定缺乏多能性标志物(例如 OCT4)和 NPC 标志物(例如 PAX6、Nestin 和 Sox1)的分化效率。
- 在此阶段,分离的NPC可以在专门的商业NPC冷冻介质中冷冻(见 材料表)并在液氮中储存长达3个月。在一次冻融后,NPC仍然保留多能性,以产生具有可靠协议的神经元,星形胶质细胞和OPC。
- 少突胶质细胞前体细胞(OPC)生成(~7天)。流程图请参见 图 2A 。
- 在第7天,使用细胞分离溶液(参见 材料表)传代NPC,并以每孔1-2×10 5个细胞的 接种密度将它们接种在24孔板中,在温暖(37°C)的神经诱导培养基中加上来自商业试剂盒的SMAD抑制剂(参见 材料表)。
- 在第8天,在OPC分化培养基中制备1%DMSO的溶液,并将铺板的NPC处理24小时。OPC 分化培养基由:DMEM/F12 培养基、1% N2 补充剂、1% B27 添加剂、20 ng/mL 的 bFGF、1 μM 的 SAG、10 ng/mL 的 PDGF-AA 组成(参见 材料表)。
- 在第9天,用不含DMSO的新鲜OPC分化培养基替换培养基。每隔一天喂一次细胞,直到第15天。如果细胞在第15天之前达到汇合,则按照步骤2.2.1中所述,将它们传代至每孔1-2×105 个细胞的接种密度。
- 在第 14 天,将 OPC 以 1-2 x 105 个细胞 /孔的密度在 OPC 分化培养基中接种 24 孔板。
- 在此阶段(第 15 天),通过 IHC 染色或 qPCR 测试细胞是否存在 OPC 特异性标志物(例如,O4、Olig1/2、CSPG4/Ng2、NKX2.2、PDGFRa; 图2B)以及缺乏鼻咽癌标志物(Pax6或Nestin; 图 2D)。我们通常在第15天检测到超过95%的细胞中的O4免疫反应性。与阿尔茨海默病特别相关的是, APP (淀粉样蛋白前体蛋白)、 BACE1 (加工蛋白酶β-秒化酶1)和肽淀粉样蛋白-β(Aβ)的表达在OPCs中含量丰富(图2F)。
- 少突胶质细胞 (OL) 成熟(~7-20 天)
- 在第15天,用OL成熟培养基替换培养基:神经基础-A培养基,2%B27补充剂,1μM cAMP,200ng / mL T3三碘甲状腺原氨酸和1μM的氯马斯汀(参见 材料表)。如有必要,每隔一天或每天更换培养基。
- 当细胞达到90%汇合时,以1:3的比例分裂至最多2代或直到细胞分裂显着减慢。如果OPCs分裂太快并在不到3天的时间内达到汇合,则以2-5μM的浓度添加Ara-C(参见 材料表)1-3天。活性增殖表明成熟效率降低。
- 通过评估OL标志物的表达来检查少突胶质细胞成熟的效率,例如CLDN11,PLP1,通过qPCR,IHC染色或免疫印迹的MBP。高度复杂结构的特征形态(图2C)和OL标志物的表达(图2E)应在第28天之前轻松检测到。
3. 人诱导神经元 (iN) 和少突胶质细胞前体细胞 (iOPC) 的共培养
- iOPC电镀(~3天)
- 在第14天以1 x 105个细胞 的密度在OPC分化培养基中的24孔板(如上文步骤2.2.4所述)中接种iOPC。
- iN-iOPC共培养建立
- 在第15天,在嘌呤霉素选择后第2天的步骤中分离诱导的人神经元(如步骤1.2.6中所述)与细胞分离溶液(参见 材料表)。
- 将神经元添加到培养的OPC上,以每孔2 x 105 个细胞的接种密度在具有生长OPC的24孔板中接种(从步骤3.1.1开始)。使用含有神经基础-A 培养基、2% B27 补充剂和 100 ng/mL T3 三碘甲状腺原氨酸的共培养基。第二天更换培养基,之后每隔一天更换一次。如果OPCs增殖太快并在不到3天的时间内达到汇合,则以2-5μM的浓度添加Ara-C。7天后神经元共培养生长的iN和iOPC的代表性图像如图 3A所示。
- 使用上述步骤1.2.7中制备的冷冻神经元与OPC共培养。以每孔 3 x 105 个细胞的更高密度冻融神经元。
- 在共培养的第14-16天之后,可以通过突触前和突触后标记物的IHC染色来观察iN中的突触形成,到第21天,突触点应该丰富(图3C),并且可以可靠地记录神经元活动。
- 从第 21 天开始,测试细胞的 OL 特异性标志物(例如,MBP 和 PLP1)。到第28天,我们通常观察到iOL过程对iN轴突的鞘现象,通过IHC染色标记特定标志物(图3B;iN轴突的神经丝NF和iOPC过程的MBP)。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
从人多能干细胞直接产生人诱导神经元
非常重要的是,起始人多能干细胞表现出高度的多能性,以成功生成iN或iOPC / iOLs。因此,在开始本手稿中描述的任一诱导方案之前,应针对特定标记物(例如Oct4和SOX2)对细胞进行染色(图1A)。按照Zhang等人先前发表的方案,使用人H1细胞获得诱导的兴奋性前脑神经元,并进行一些修改(图1C)12,16,17,18。在这里,我们提出了一个方案,其中在第2天的iN在基质溶液的纯培养物中重新铺板(参见材料表),没有任何饲养层:胶质细胞或成纤维细胞。除了先前发表的方案外,我们观察到在第2天冷冻iN不会显着影响细胞活力(解冻后~15%-20%细胞死亡)。培养物中的纯神经元将在第14-16天开始表达突触蛋白1(图1D)。建立纯神经元培养非常重要,因为某些因素,例如领先的AD危险因素ApoE,可以由饲养层中的细胞表达,这可能会显着混淆结果。
通过DMSO处理改善iOPC的生成和iOL成熟
在这里,我们提出了一种快速有效的协议,可以在 2 周内生成 iOPC,并在 4-5 周内生成成熟的 iOL(图 2A)。我们利用我们之前开发的瞬时DMSO处理方法来增强ES和iPS细胞29,30,31的分化效率。DMSO处理丰富了早期G1阶段的细胞数量,以实现更好的信号整合,有利于分化。我们在诱导人ES细胞产生NPC之前进行了第一次治疗,在将NPC分化为iOPC之前进行了第二次治疗。我们最早可以在ES H1细胞接种后2周检测到特定的OPC标志物(Olig2,CSPG4,NKX2.2和PDGFRA)(图2B,E)。这个阶段的iOPC群体相当均匀,>95%的细胞对O4染色呈阳性,对其他标志物具有高水平的免疫反应性(图2B)。在第15天OL成熟开始后,我们通常可以从第28天开始检测特定的OL标志物(MBP,O1,CLDN11和PLP1)(图2C,E)。这些阶段特异性标志物的表达与少突胶质细胞的发育过程相关,并表明速度加快,NPC标志物逐渐下降,OPC标志物在第二周左右达到峰值,OL标志物在第三周升高(图2D,E)37。请注意,这种成熟过程使细胞群多样化。连续体中的亚群,包括OPC和成熟髓鞘少突胶质细胞之间的多个中间阶段,可以存在并占总细胞的不同百分比,稍后更成熟的细胞占主导地位。
作为比较,我们购买了高度引用的iOPC,并按照制造商协议在iOL中成熟它们。我们测试了上述标记物在iOPC和iOL制剂以及我们购买的细胞中的表达。我们确定按照我们的方案生成的细胞在所有测试基因中具有更高的表达(图2E)。有趣的是,当我们测试iNs与iOPCs中淀粉样蛋白-β的两种主要亚型(Aβ40和Aβ42)的分泌水平时,我们注意到iOPCs分泌了更多的两个片段,但比例保持不变(图2F)。
iN和iOPCs的共培养
该协议专门针对共培养iN和iOPC进行了优化,并允许我们在神经发育过程中实时监测这两种细胞类型之间的细胞间通信。两种细胞类型的理想镀层密度需要通过一系列细胞数滴定来确定,以实现适当的分化(图3A)。在共培养4周后,iOPCs有望充分分化为对特定标志物(如MBP)呈阳性的OL,并将过程扩展到鞘轴突(图3B)。共培养系统可以有力地增加突触的数量,表明iOPCs通过物理接触或营养因子的释放提供神经元支持(图3C)。我们可以将共培养物保持在可接受的健康状况长达 6 周,并观察到突触数量和其他神经元属性在第五周左右趋于平稳。值得注意的是,星形胶质细胞和小胶质细胞在我们的制剂中不存在,可以通过检查特定标志物的表达来记录它们的缺失(图3D)。iOPCs表达大量表征良好的基因,这些基因可以潜在地响应和介导来自邻近神经元的活动依赖性信号,以旁分泌(例如神经营养因子和代谢物)和/或突触方式(图3E 和 3F)。

图 1:从 hPSC 直接生成人类诱导神经元 (iN)。 (A)iN生成流程图。(B)人多能干细胞(H1)起始培养物的代表性明场和免疫荧光图像,以确认多能性。Oct4 显示为红色,Sox2 显示为绿色。(C)第4天和第6天iN的代表性明场图像。(D)在纯培养物中生长24天的iN中树枝状树枝状树状化和突触点的特征形态,并通过树突状标记物Map2和突触前标记突触1(Syn1)的免疫荧光染色染色。 请点击此处查看此图的大图。

图 2:iOPC 生成和 iOL 成熟。 (A) iOPC和iOL生成流程图。(B)第15天iOPCs的代表性明场和免疫荧光图像。Olig2(泛少突胶质标记)显示为绿色,O4(OPC 标记)显示为红色,DAPI 显示为蓝色。成像显示,>95%的iOPCs对O4呈阳性,25%对Olig2呈阳性。(C)第28天iOLs的代表性明场和免疫荧光图像。MBP 显示为绿色,O1 显示为红色,DAPI 显示为蓝色。(D)NPC标志物PAX6的表达在第14天在iOPCs中显着降低,并在第28天进一步降低到OLs的背景,表明iOPC群体中具有强大的NPC转分化和高水平的同质性。(E)所述方案产生的培养物中常见OPC和OL标记基因的时程表达谱,无需(-DMSO)或(+DMSO)DMSO孵育步骤(步骤2.1.3和2.2.2),在不同时间点测定。作为比较,商业iOPC(见 材料表)根据制造商的说明成熟,并且两种iOPC(iOPC-Tempo)或iOL(iOL-Tempo)都针对相同的标记进行测试。正如预期的那样,在所有测试的iOPCs中,在分化的早期阶段未检测到MBP(一种成熟的少突胶质细胞标志物)。DMSO显著提高了OPC分化和OL成熟的效率。(F)纯iN和iOPCs培养物中Aβ40和Aβ42的产生和分泌,通过商业ELISA试剂盒(参见 材料表)在从纯iN和iOPCs培养物中获得的上清液测量,在第15天并通过细胞数归一化(均在24孔板中每孔200,000个细胞的密度下)。 请点击此处查看此图的大图。
条形图中的数据绘制为平均± SEM (n ≥ 3)。通过学生 t检验(*,p < 0.05; ***, p < 0.001)评估统计学意义;在(D)中,与NPC相比;在(E)中,与对照iOPC-Tempo相比;在 (F) 中,与 iN 相比。

图 3:iN 和 iOPC 的共培养。 (A)第7天共培养的iN和iOPCs的代表性明场图像,显示进一步成熟的适当密度。(B)共培养28天的iN和iOPCs的代表性免疫荧光图像。轴突标志物神经丝NF显示为绿色,少突胶质细胞标志物MBP显示为红色。右图,一段通过iOL工艺(MBP+)包裹的iN轴突。(C)在4周大的共培养物中测定的突触形成。对细胞进行突触素1(Syn1,绿色)和MAP2(红色)染色,并通过沿树突状片段的密度共聚焦分析来量化突触点,如17,18所述。(D)在我们的iN和iOPCs共培养物(共培养7天)中,星形胶质细胞标志物ALDHL1和GFAP的表达最小(顶部),并且小胶质细胞标志物TMEM119,TREM2和CD33的表达未通过qPCR检测到(N.D.)。因此,这两种神经胶质细胞类型的污染被排除在外。(E)iOPC与iN共培养导致神经元-OPC突触的形成。荧光标记的突触后标志物PSD95-mCherry仅在OPC中表达,在单个培养物中显示弥漫模式(左),但在共培养物中聚集形成点状(右,用箭头表示;Tuj1,神经元标记)。(F)在第14天表达特征明确的少突胶质细胞基因,这些基因可以在iOPCs的纯培养物中感知和响应神经元活动。请点击此处查看此图的大图。
条形图中的数据绘制为 SEM ±平均值 (n ≥ 3)。通过学生 t检验(**,p < 0.005; ***,p < 0.001)评估统计学意义;在(C)中,与无OPC条件相比;在(D)中,与顶部图中的原代星形胶质细胞相比。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
除了通过髓鞘形成稳定突触结构和促进盐信号传导的物理和代谢支持外,少突胶质细胞谱系细胞还可以通过与神经元的快速和动态串扰来塑造神经元活动模式5,6,7。虽然在AD病理学中,少突胶质反应最初被认为仅继发于炎症和氧化应激,但现在有有希望的证据表明,髓鞘完整性受损是Aβ聚集和tau过度磷酸化出现之前的早期致病事件9。此外,通过OPC的自我更新修复髓鞘形成在AD38中特别脆弱,这一过程在很大程度上依赖于神经元活动。因此,了解支持健康神经元 - 少突胶质细胞信号传导的机制代表了确定新治疗靶点的绝佳机会。
单转录因子Ngn2方案是用于生成干细胞衍生的人类神经元最常用的技术之一,此处概述的程序是获得纯神经元培养物的进一步改进。我们的iOPC/iOL方案的诱导期比之前发表的研究短(4至24周),其产量和纯度可与其他常用方案相媲美19,20,22,23,24,25,26,27,28.我们的协议通过表征的模式线索将ES细胞逐步分化为NPC,OPC,最后是少突胶质细胞,并产生可用于研究髓鞘形成稳态的调节和体外或体内修复的功能细胞(例如,通过植入颤抖小鼠模型)如前工作中所述。DMSO孵育极大地促进了我们方案的改进,DMSO孵育激活视网膜母细胞瘤蛋白并延长细胞周期的G1期,以更好地整合定向分化的刺激,并且还增强了终末分化为功能衍生物29,30。最后,使用克立姆定(一种通过药物筛选鉴定的毒蕈碱和抗组胺化合物)用于髓鞘再生治疗剂33,另外缩短了少突胶质细胞的成熟,如在iPS细胞制备和活体动物7,28中观察到的那样。
该技术的局限性主要在于简化的体外设置与大脑体内环境之间的内在差异;这种差异导致个体脑细胞类型在晚期阶段完全发育潜力的折扣。对于iN,最近的研究能够在相当长的一段时间内保持培养物的良好突触健康,但仍显示出一些相对不成熟,表现为“旧”iN培养物(即使是25个月大的)中的脊柱样结构减少和自发突触传递受损39。虽然iOPCs在移植到转基因小鼠大脑后已在体内可重复地显示对髓鞘轴突,但具有电子显微镜评估的体外髓鞘化测定仍然是一个技术挑战,几乎所有已发表的方案的效率都不令人满意19,28,包括这个。因此,我们的神经元-OPC共培养系统预计不会忠实地模仿大脑衰老过程以及AD病理学的晚期。相反,它的独特之处在于解开神经元与OPCs或早期少突胶质细胞之间复杂的细胞间相互作用,这些相互作用独立于髓鞘形成,但对于适当的神经发育和疾病发病机制至关重要。
这里描述的三个过程中的每一个在协议中都有其关键步骤,可能需要修改和故障排除。对于iN方案,有两个关键步骤:嘌呤霉素选择(步骤1.2.5)和铺板密度(步骤1.2.6)。不完全去除转导不足的细胞会导致低分化细胞的污染,并损害神经元的存活和功能。必须考虑步骤1.2.5中所述的具有更高浓度和更长孵育时间的更强嘌呤霉素选择的修饰。合适的接种密度应通过滴定每个多能细胞系来确定,因为低密度会导致培养物塌陷,而高密度会促进细胞聚集并阻碍神经元生长。对于iOPC / iOL方案,两个关键步骤是控制OPC分化中的细胞增殖(步骤2.2.3)和OL成熟的铺板密度(步骤2.2.4和2.3.2)。分化鼻咽癌的过度生长表明对OPC分化刺激的反应不佳,需要通过适当剂量的Ara-C治疗(在指示范围内)来抑制。在电镀OPC进行成熟时,这里优选较低的细胞密度范围,因为稀疏分布可以促进复杂结构的生理形态的诱导(如图 2C所示)。对于iN-iOPC共培养方案,我们想提请注意为两种细胞类型以适当密度进行接种的关键步骤(步骤3.1.1和3.2.2)。具体来说,iN可能无法很好地附着在生长的OPC之间的表面,并且当培养物达到汇合时倾向于首先分离。最佳比例必须通过滴定细胞数量来确定。
总体而言,存在于我们协议中的这种还原论方法是从人脑固有的复杂性中剖析特定异细胞相互作用的有力工具,并有助于揭示健康和AD中的少突胶质细胞生物学。因此,我们认为,就现有方法而言,其重要性是相当明显的。这里为未来应用开发的方法的另一个用途是基于细胞的脱髓鞘治疗,例如放疗后40 和脊髓损伤41,42。此外,这种基于干细胞的系统的高通量能力还可以在更大范围内用于筛选小分子文库,以寻找可以保护或恢复神经元、OPC、少突胶质细胞及其相互作用的生理状态的化合物。因此,我们相信这里描述的协议将有助于未来开发更好的建模工具和AD和其他神经退行性疾病的有效治疗方法。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者没有什么可透露的。
Acknowledgments
这项工作得到了美国国立卫生研究院(R00 AG054616 至 YAH,T32 GM136566 至 K.C.)、斯坦福大学医学院和 Siebel 奖学金(授予 SC)的资助。YAH是布朗转化科学研究所转化神经科学中心的GFL转化教授。
Materials
| Name | Company | Catalog Number | Comments |
| Accutase | STEMCELL Technologies | 7920 | |
| B27 supplement | ThermoFisher | 17504044 | |
| bFGF | ThermoFisher | PHG 0266 | |
| cAMP | MilliporeSigma | A9501 | |
| Clemastine | MilliporeSigma | SML0445 | |
| DMEM/F12 medium | STEMCELL Technologies | 36254 | |
| DMSO | ThermoFisher | D12345 | |
| Doxycycline | MilliporeSigma | D3072 | |
| Fetal Bovine Serum | ScienCell | 10 | |
| H1 human ES cells | WiCell | WA01 | |
| Matrigel | Corning | 354234 | |
| mTeSR plus | STEMCELL Technologies | 5825 | |
| N2 supplement | ThermoFisher | 17502001 | |
| Neurobasal A medium | ThermoFisher | 10888-022 | |
| Non Essential Amino Acids | ThermoFisher | 11140-050 | |
| PDGF-AA | R&D Systems | 221-AA-010 | |
| PEI | VWR | 71002-812 | |
| pMDLg/pRRE | Addgene | 12251 | |
| Polybrene | MilliporeSigma | TR-1003-G | |
| pRSV-REV | Addgene | 12253 | |
| Puromycin | ThermoFisher | A1113803 | |
| ROCK Inhibitor Y-27632 | STEMCELL Technologies | 72302 | |
| SAG | Tocris | 4366 | |
| STEMdiff Neural Progenitor Freezing Media | STEMCELL Technologies | 5838 | |
| STEMdiff SMADi Neural Induction Kit | STEMCELL Technologies | 8581 | |
| T3 triiodothyronine | MilliporeSigma | T6397 | |
| Tempo-iOlogo: Human iPSC-derived OPCs | Tempo BioScience | SKU102 | |
| TetO-Ng2-Puro | Addgene | 52047 | |
| VSV-G | Addgene | 12259 |
References
- Pelvig, D. P., Pakkenberg, H., Stark, A. K., Pakkenberg, B. Neocortical glial cell numbers in human brains. Neurobiology of Aging. 29 (11), 1754-1762 (2008).
- Barres, B. A. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 60 (3), 430-440 (2008).
- De Strooper, B., Karran, E. The cellular phase of Alzheimer's disease. Cell. 164 (4), 603-615 (2016).
- Monje, M. Myelin plasticity and nervous system function. Annual Review of Neuroscience. 41, 61-76 (2018).
- Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., Bergles, D. E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nature Neuroscience. 21 (5), 696-706 (2018).
- Gibson, E. M., et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science. 344 (6183), 1252304 (2014).
- Pan, S., Mayoral, S. R., Choi, H. S., Chan, J. R., Kheirbek, M. A. Preservation of a remote fear memory requires new myelin formation. Nature Neuroscience. 23 (4), 487-499 (2020).
- Thornton, M. A., Hughes, E. G. Neuron-oligodendroglia interactions: Activity-dependent regulation of cellular signaling. Neuroscience Letters. 727, 134916 (2020).
- Ettle, B., Schlachetzki, J. C. M., Winkler, J. Oligodendroglia and myelin in neurodegenerative diseases: more than just bystanders. Molecular Neurobiology. 53 (5), 3046-3062 (2016).
- Essayan-Perez, S., Zhou, B., Nabet, A. M., Wernig, M., Huang, Y. A. Modeling Alzheimer's disease with human iPS cells: advancements, lessons, and applications. Neurobiology of Disease. 130, 104503 (2019).
- Li, L., et al. GFAP mutations in astrocytes impair oligodendrocyte progenitor proliferation and myelination in an hiPSC model of Alexander disease. Cell Stem Cell. 23 (2), 239-251 (2018).
- Lin, Y. T., et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer's disease phenotypes in human iPSC-derived brain cell types. Neuron. 98 (6), 1294 (2018).
- TCW, J., et al. Cholesterol and matrisome pathways dysregulated in human APOE ε4 glia. bioRxiv. , (2019).
- Ang, C. E., Wernig, M.
- Penney, J., Ralvenius, W. T., Tsai, L. H. Modeling Alzheimer's disease with iPSC-derived brain cells. Molecular Psychiatry. 25 (1), 148-167 (2020).
- Zhang, Y., et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron. 78 (5), 785-798 (2013).
- Huang, Y. A., Zhou, B., Nabet, A. M., Wernig, M., Sudhof, T. C. Differential signaling mediated by ApoE2, ApoE3, and ApoE4 in human neurons parallels Alzheimer's Disease risk. Journal of Neuroscience. 39 (37), 7408-7427 (2019).
- Huang, Y. A., Zhou, B., Wernig, M., Sudhof, T. C. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Abeta Secretion. Cell. 168 (3), 427-441 (2017).
- Yang, N., et al. Generation of oligodendroglial cells by direct lineage conversion. Nature Biotechnology. 31 (5), 434-439 (2013).
- Douvaras, P., et al. Efficient generation of myelinating oligodendrocytes from primary progressive multiple sclerosis patients by induced pluripotent stem cells. Stem Cell Reports. 3 (2), 250-259 (2014).
- Lee, E. H., Park, C. H. Comparison of reprogramming methods for generation of induced-oligodendrocyte precursor cells. Biomolecules & Therapeutics (Seoul). 25 (4), 362-366 (2017).
- Ehrlich, M., et al. Rapid and efficient generation of oligodendrocytes from human induced pluripotent stem cells using transcription factors. Proceedings of the National Academy of Sciences of the United States of America. 114 (11), 2243-2252 (2017).
- Rodrigues, G. M. C., et al. Defined and scalable differentiation of human oligodendrocyte precursors from pluripotent stem cells in a 3D culture system. Stem Cell Reports. 8 (6), 1770-1783 (2017).
- Hu, B. Y., Du, Z. W., Li, X. J., Ayala, M., Zhang, S. C. Human oligodendrocytes from embryonic stem cells: conserved SHH signaling networks and divergent FGF effects. Development. 136 (9), 1443-1452 (2009).
- Izrael, M., et al. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Molecular and Cellular Neuroscience. 34 (3), 310-323 (2007).
- Yamashita, T., et al. Differentiation of oligodendrocyte progenitor cells from dissociated monolayer and feeder-free cultured pluripotent stem cells. PLoS One. 12 (2), 0171947 (2017).
- Wang, S., et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell. 12 (2), 252-264 (2013).
- Chanoumidou, K., Mozafari, S., Baron-Van Evercooren, A., Kuhlmann, T. Stem cell derived oligodendrocytes to study myelin diseases. Glia. 68 (4), 705-720 (2020).
- Chetty, S., et al. A simple tool to improve pluripotent stem cell differentiation. Nature Methods. 10 (6), 553-556 (2013).
- Li, J., et al. A transient DMSO treatment increases the differentiation potential of human pluripotent stem cells through the Rb family. PLoS One. 13 (12), 0208110 (2018).
- Sambo, D., Li, J., Brickler, T., Chetty, S. Transient treatment of human pluripotent stem cells with DMSO to promote differentiation. Journal of Visualized Experiments: JoVE. (149), (2019).
- Douvaras, P., Fossati, V. Generation and isolation of oligodendrocyte progenitor cells from human pluripotent stem cells. Nature Protocols. 10 (8), 1143-1154 (2015).
- Mei, F., et al. Micropillar arrays as a high-throughput screening platform for therapeutics in multiple sclerosis. Nature Medicine. 20 (8), 954-960 (2014).
- Madhavan, M., et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nature Methods. 15 (9), 700-706 (2018).
- Zhang, Y., et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 89 (1), 37-53 (2016).
- Grubman, A., et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer's disease reveals cell-type-specific gene expression regulation. Nature Neuroscience. 22 (12), 2087-2097 (2019).
- Goldman, S. A., Kuypers, N. J. How to make an oligodendrocyte. Development. 142 (23), 3983-3995 (2015).
- Behrendt, G., et al. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia. 61 (2), 273-286 (2013).
- Patzke, C., et al. Neuromodulator signaling bidirectionally controls vesicle numbers in human synapses. Cell. 179 (2), 498-513 (2019).
- Piao, J., et al. Human embryonic stem cell-derived oligodendrocyte progenitors remyelinate the brain and rescue behavioral deficits following radiation. Cell Stem Cell. 16 (2), 198-210 (2015).
- Keirstead, H. S., et al. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. Journal of Neuroscience. 25 (19), 4694-4705 (2005).
- Kim, D. S., et al. Rapid generation of OPC-like cells from human pluripotent stem cells for treating spinal cord injury. Experimental & Molecular Medicine. 49 (7), 361 (2017).
Tags
神经科学,第 165 期,诱导多能干细胞、iPS 细胞、人胚胎干细胞、ES 细胞、神经元、少突胶质细胞、少突胶质细胞祖细胞、OPC、阿尔茨海默病、淀粉样蛋白 β 肽、突触Erratum
Formal Correction: Erratum: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions
Posted by JoVE Editors on 12/29/2020.
Citeable Link.
An erratum was issued for: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions. The Representative Results section has been updated.
Figure 3 was updated from:
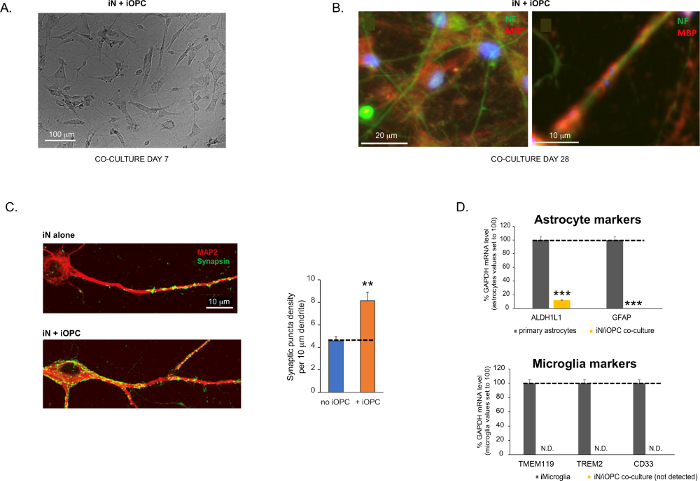
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. Please click here to view a larger version of this figure.
to:
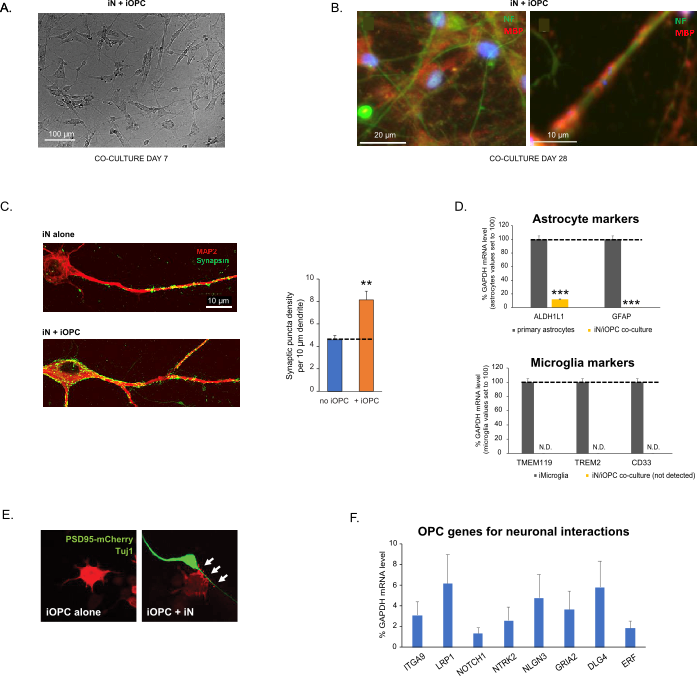
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. (E) Coculturing iOPC with iN leads to the formation of neuron-OPC synapses. The fluorescence-tagged post-synaptic marker PSD95-mCherry is expressed only in OPCs, and display a diffuse pattern in single cultures (left) but aggregate to form puncta in cocultures (right, indicated by arrows; Tuj1, neuronal marker). (F) The expression of well-characterized oligodendroglial genes that can sense and respond to neuronal activities in the pure cultures of iOPCs at Day 14. Please click here to view a larger version of this figure.
The fourth paragraph was updated from:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D).
to:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D). The iOPCs express a good number of well-characterized genes that can potentially respond to and mediate the activity-dependent signals from neighboring neurons, in a paracrine (e.g. neurotrophins and metabolites) and/or a synaptic manner (Figure 3E and 3F).
