

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Neuron-glial-interaktionerne i neurodegeneration forstås ikke godt på grund af utilstrækkelige værktøjer og metoder. Her beskriver vi optimerede protokoller til opnåelse af inducerede neuroner, oligodendrocytprecursorceller og oligodendrocytter fra humane pluripotente stamceller og giver eksempler på værdierne af disse metoder til forståelse af celletypespecifikke bidrag i Alzheimers sygdom.
Abstract
I Alzheimers sygdom (AD) og andre neurodegenerative lidelser er oligodendroglial svigt et almindeligt tidligt patologisk træk, men hvordan det bidrager til sygdomsudvikling og progression, især i hjernens grå stof, forbliver stort set ukendt. Dysfunktionen af oligodendrocyt-afstamningsceller er kendetegnet ved mangler i myelinering og nedsat selvfornyelse af oligodendrocytprecursorceller (OPC'er). Disse to defekter er forårsaget i det mindste delvist af forstyrrelsen af interaktioner mellem neuron og oligodendrocytter langs opbygningen af patologi. OPC'er giver anledning til myeliniserende oligodendrocytter under CNS-udvikling. I den modne hjernebark er OPC'er de vigtigste proliferative celler (bestående af ~ 5% af de samlede hjerneceller) og styrer ny myelindannelse på en neural aktivitetsafhængig måde. Sådan neuron-til-oligodendrocytkommunikation er signifikant undervurderet, især i forbindelse med neurodegenerative tilstande som AD, på grund af manglen på passende værktøjer. I de senere år har vores gruppe og andre gjort betydelige fremskridt for at forbedre de nuværende tilgængelige protokoller til at generere funktionelle neuroner og oligodendrocytter individuelt fra humane pluripotente stamceller. I dette manuskript beskriver vi vores optimerede procedurer, herunder etableringen af et co-kultursystem til at modellere neuron-oligodendrocytforbindelserne. Vores illustrative resultater tyder på et uventet bidrag fra OPC'er / oligodendrocytter til hjernens amyloidose og synapse integritet og fremhæver nytten af denne metode til AD-forskning. Denne reduktionistiske tilgang er et kraftfuldt værktøj til at dissekere de specifikke heterocellulære interaktioner ud af den iboende kompleksitet inde i hjernen. De protokoller, vi beskriver her, forventes at lette fremtidige undersøgelser af oligodendrogliale defekter i patogenesen af neurodegeneration.
Introduction
Oligodendrocyt-afstamningsceller - herunder oligodendrocytprecursorceller (OPC'er), myeliniserende oligodendrocytter og overgangstyper derimellem - udgør en hovedgruppe af humane hjerneceller1, der aktivt deltager i mange kritiske funktioner for korrekt drift og vedligeholdelse af vores centralnervesystem gennem neural udvikling og aldring 2,3,4 . Mens oligodendrocytter er velkendte for at producere myelin for at lette neuronal aktivitetsoverførsel og understøtte aksonal sundhed i hvidt stof, er OPC'er rigelige (~ 5%) i gråt stof, hvor myelinering er knap og udfører aktivitetsafhængige signalfunktioner til at styre læringsadfærd og hukommelsesdannelse 5,6,7,8 . Hvordan oligodendrogliale celler fungerer og dysfunktion i patogenesen af Alzheimers sygdom (AD) og andre aldersassocierede neurodegenerative tilstande er blevet undervurderet9. Utilstrækkeligheden af et passende modelsystem og mangler i den generelle viden til at styre en eksperimentel vej fremad er hovedårsagerne til denne kløft.
I lyset af de seneste gennembrud inden for udlede humane hjerneceller fra pluripotente stamceller, herunder embryonale stamceller (ES) og inducerede pluripotente stamceller (iPS), er sådanne cellulære modeller i forbindelse med moderne genredigeringsværktøjer opstået som robuste værktøjer til at håndtere den indviklede sammenhæng mellem cellulære interaktioner i hjernen og er i stand til at demonstrere humanspecifikke sygdomsmanifestationer10, 11. I betragtning af at individuelle hjernecelletyper kan udvise forskellige og endda modstridende virkninger i lyset af de samme AD-fremmende forhold12,13, tilbyder denne stamcellemetode unikt celletypespecifik information, der tidligere er blevet savnet ved hjælp af etablerede in vivo- eller in vitro-modeller, der kun giver aggregerede aflæsninger fra samlinger af hjernecelletyper. I det sidste årti er der udviklet et stort antal pålidelige protokoller til at generere humane neuroner fra transdifferentiering af ES / iPS-celler eller direkte konvertering fra andre terminalt differentierede celletyper (f.eks. Fibroblaster)14,15. Især kan anvendelsen af centrale neurogene transkriptionsfaktorer (f.eks. Neurogenin 2, Ngn2)16 på humane pluripotente stamceller generere en homogen population af velkarakteriserede neuronale celletyper til rene kulturer uden behov for cokulturering med gliaceller12,17,18. For inducerede humane oligodendrocytter er der et par offentliggjorte protokoller, der kan generere funktionelle celler, der ligner deres primære modstykker, med en bred vifte af effektivitet og efterspørgsel i tid og ressourcer 19,20,21,22,23,24,25,26,27,28 . Til dato er ingen af disse protokoller blevet anvendt til at undersøge, hvordan oligodendrogliale celler reagerer på og påvirker AD-patogenese.
Her beskriver vi vores forbedrede protokoller for enkelt- og blandede kulturer af humaninducerede neuroner (iN'er) og OPC'er / oligodendrocytter (iOPC'er / iOL'er). Den her beskrevne iN-protokol er baseret på den meget anvendte Ngn2-tilgang16 og har den ekstra funktion at være gliafri. De resulterende iN'er er homogene og ligner meget det kortikale lag 2/3 excitatoriske neuroner med karakteristisk pyramidemorfologi, genekspressionsmønster og elektrofysiologiske træk17,18 (figur 1). For at overvinde nogle af de grundlæggende barrierer i rettet differentiering af pluripotente stamceller har vi udviklet en enkel og effektiv metode til lavdosis dimethylsulfoxid (DMSO) forbehandling29,30 og rapporteret en forbedret tilbøjelighed hos humane ES/iPS-celler til at transdifferentiere til iOPC'er og iOL'er31, baseret på en bredt tilpasset protokol af Douvaras og Fossati 32 . Vi har yderligere forenklet protokollen og indarbejdet en robust differentieringsfremmende forbindelse, clemastin 7,33,34, for at fremskynde processen med oligodendroglial modning. Som et resultat (figur 2) kan iOPC'erne genereres på 2 uger (~ 95% positive for markøren O4) og iOL'erne på fire uger (udtrykker modne markører MBP og PLP1). Interessant nok fandt vi, at iOPC'er alene udskiller en bemærkelsesværdig mængde amyloid-β (Aβ), i overensstemmelse med de uafhængige transkriptomiske data, der viser den rigelige ekspression af amyloidprecursorproteinet (APP) og behandlingsprotease β-sekretase (BACE1) i oligodendrocyt-afstamningsceller35,36. Desuden fremmer vores iN-iOPC-samkultursystem ensheathing af axoner ved MBP-positive iOL-processer og giver betydelig støtte til synapsedannelse (figur 3). Således har de protokoller, vi har beskrevet nedenfor, tekniske og biologiske fordele i forhold til tidligere katalogiserede neuron-oligodendroglia co-dyrkningsmetoder og har et løfte om bedre modellering af neurodegenerationen i AD.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Induktion af humant neuron fra humane pluripotente stamceller
- Lentivirus forberedelse (~ 5 dage, detaljeret protokol som beskrevet tidligere16)
- Plade ~ 1 million HEK293T-celler hver T75-kolbe for at få dem ~ 40% sammenløbende, når de udfører transfektion. Transficere dem med plasmider, der udtrykker tetracyklininducerbart Ngn2 og puromycinresistent gen (PuroR; under samme TetO-promotorkontrol), rtTA og de tre hjælperplasmider pRSV-REV, pMDLg/pRRE og VSV-G (12 μg lentiviralt vektor-DNA og 6 μg af hvert af hjælperplasmid-DNA'et). Forbered mindst tre kolber pr. Lentiviruspræparat. Brug PEI til transfektion efter producentens anvisninger. Skift mediet efter 16 timer, og kassér.
- Høst frigav virale partikler ved at indsamle kulturmedier hver dag og erstatte med friske medier i 3 dage. Saml de indsamlede medier, der indeholder virale partikler, til rensning. Filtrer virussen gennem et 0,22 μm filter og centrifuge ved 49.000 x g i 90 min. Resuspender pelleten i det passende volumen PBS-glucose (~ 150 μL).
- Neuron induktion (~ 5 dage)
BEMÆRK: Denne induktionsprotokol (figur 1A; flowdiagram) er yderst effektiv til både iPS- og ES-celler med valideret pluripotens (som kan analyseres ved immunhistokemifarvning af velkarakteriserede pluripotensmarkører; Figur 1B).- Brug kommercielt tilgængelige H1 humane ES-celler ved passage af 52 (se materialetabel). Dyrkning af cellerne på ekstracellulære matrixopløsningsbelagte 6-brøndsplader (~ 0,5 mg matrixopløsning pr. 6-brøndsplade; se Materialetabel) ved hjælp af ES-cellevedligeholdelsesmedium (se Materialetabel) og inkubere pladerne ved 37 ° C med 5% CO2.
- På dag -2 afmonteres ES-celler (80% sammenflydende) med 1 ml celleløsningsopløsning (se materialetabel) og inkuberes ved stuetemperatur i 10 min. Overfør cellerne til et rør; vask brønden med 2 ml medier og kombiner i samme rør. Centrifuge ved 300 x g i 5 minutter, suspender pelleten i medier og plade cellerne på matrixbelagte 6-brøndsplader ved såtætheden på 1 x 105 celler pr. Brønd.
- På dag -1 tilsættes lentivirus, der udtrykker Ngn2 plus PuroR og rtTA sammen med polybren (8 μg / ml) til ES-cellerne i frisk ES-cellevedligeholdelsesmedium (se Materialetabel). Den nøjagtige mængde vira skal bestemmes af faktiske titere eller titreringen. Vi tilføjer typisk 5 μL hver virus pr. brønd i en 6-brøndsplade.
- På dag 0 tilsættes Doxycyclin (2 μg/ml for at aktivere Ngn2-ekspression) i DMEM-F12-medium med N2-tilskud uden morfogener.
- På dag 1 tilsættes Puromycin i frisk substrat af DMEM-F12 plus N2 og doxycyclin til den endelige koncentration på 1 μg / ml medium. Udvalgte transducerede celler i Puromycin i mindst 24 timer. Højere Puromycinkoncentration (op til 5 μg/ml) og længere selektionsperiode (op til 48 timer) kan være påkrævet for at fjerne de undertransducerede celler tilstrækkeligt, hvis virustiteren er lav.
- På dag 2 skal du løsne differentierende neuroner med celleløsningsopløsning (se Materialetabel) og genplade dem på 24-brøndsplader (mellem 80.000-200.000 celler / brønd) belagt med matrixopløsning (se Materialetabel) og vedligeholde dem i NBA / B27-medium uden doxycyclin. Såtætheden er kritisk.
- På dette stadium kan løsrevne neuroner fryses i specialiseret kommercielt frysemedium (se Materialetabel) og opbevares i flydende nitrogen i op til 3 måneder. Rene neuroner kan belægges og tegner sig for den typiske ~ 15% -20% celledød efter optøning, dyrket alene eller co-kultiveret med andre hjernecelletyper (se trin 3.2.3. for samdyrkning med OPC'er).
- Kultur ren iNs på pladerne belagt med ekstracellulære matrixbaserede opløsninger som instrueret af producenten (se Materialetabel). Den karakteristiske pyramidemorfologi skal være tydelig på dag 4 (og dag 6; Figur 1C). Synapsedannelsen kan detekteres allerede i dag 14 til 16 og er fremtrædende på dag 24 ved immunhistokemisk farvning med standard præ- og postsynaptiske markører. (Figur 1D; mærket med den præsynaptiske markør Synapsin 1 og dendritisk markør Map2).
2. Induktion af humane oligodendrocytprecursorceller (OPC'er) fra pluripotente stamceller og oligodendrocytmodning
- Neural Progenitor Cell (NPC) generation: monolagsprotokol (~ 7 dage). Se figur 2A for flowdiagrammet.
- Kultur H1 humane ES-celler som beskrevet tidligere (se trin 1.2.1.) og transdifferentiere dem til neurale stamceller (NPC'er) ved en etableret tilgang kaldet dobbelt SMADi med små molekylehæmmere til flere signalveje. Her bruger vi et bredt accepteret kommercielt sæt og følger monolagsprotokollen fra producenten (se Materialetabel).
- På dag -1, plade 0,5-1 x 10 6 celler pr. brønd i en 6-brønds plade belagt med en vækstfaktor reduceret matrixopløsning (se Materialetabel; ~ 0,5 mg matrixopløsning pr. 6-brøndsplade) med ES-cellevedligeholdelsesmedium (se Materialetabel). Denne vækstfaktorreducerede matrixopløsning bruges til at belægge alle de plader, der vil blive brugt i de følgende trin.
- På dag 0 behandles celler i 24 timer med ES-cellevedligeholdelsesmedium (se Materialetabel) suppleret med 2% DMSO.
- På dag 1-6 skal du skifte hele mediet med varmt (37 °C) neuralt induktionsmedium, der indeholder SMAD-hæmmere fra det kommercielle kit (se Materialetabel). Hvis celler deler sig og når sammenløb før dag 7, skal de føre dem til såtætheden på 0,5-1 x 106, som beskrevet tidligere i trin 2.1.2.
- På dag 7 passerer NPC'er ved hjælp af celleløsrivelsesopløsning (se Materialetabel) og plade ved en såtæthed på 1-2 x 105 celler / brønd af en 24-brøndsplade.
- Assay differentieringseffektiviteten ved immunohistokemisk (IHC) farvning for fravær af pluripotensmarkør, OCT4 for eksempel, og tilstedeværelse af NPC-markører såsom PAX6, Nestin og Sox1.
- På dette stadium kan løsrevne NPC'er fryses i det specialiserede kommercielle NPC-frysemedie (se Materialetabel) og opbevares i flydende nitrogen i op til 3 måneder. Efter frysning og optøning for en gangs skyld bevarer NPC'er stadig multipotensen til at give anledning til neuroner, astrocytter og OPC'er med pålidelige protokoller.
- Oligodendrocyt precursor celle (OPC) generation (~ 7 dage). Se figur 2A for flowdiagrammet.
- På dag 7 passerer NPC'er ved hjælp af celleløsrivelsesopløsning (se Materialetabel) og plade dem ved en såtæthed på 1-2 x 105 celler pr. Brønd i en 24-brøndsplade i varmt (37 ° C) neuralt induktionsmedium plus SMAD-hæmmere fra det kommercielle kit (se Materialetabel).
- På dag 8 fremstilles en opløsning af 1% DMSO i OPC-differentieringsmediet og behandles de belagte NPC'er i 24 timer. OPC-differentieringsmediet består af: DMEM / F12-medium, 1% N2-supplement, 1% B27-supplement, bFGF ved 20 ng / ml, SAG ved 1 μM, PDGF-AA ved 10 ng / ml (se materialetabel).
- På dag 9 skal du udskifte medier med frisk OPC-differentieringsmedium uden DMSO. Foder cellerne hver anden dag indtil dag 15. Hvis cellerne når sammenløb før dag 15, føres de til såtætheden på 1-2 x 105 celler pr. samt beskrevet i trin 2.2.1.
- På dag 14, plade OPC'er i OPC-differentieringsmedium ved en densitet på 1-2 x 105 celler / brønd i en 24-brøndsplade.
- På dette stadium (dag 15) testes celler for tilstedeværelsen af OPC-specifikke markører ved IHC-farvning eller qPCR (f.eks. O4, Olig1/2, CSPG4/Ng2, NKX2.2, PDGFRa; Figur 2B) og for fraværet af NPC-markører (Pax6 eller Nestin; Figur 2D). Vi registrerer typisk O4-immunreaktiviteten i mere end 95% af cellerne på dag 15. Af særlig relevans for Alzheimers sygdom er ekspressionen af APP (amyloidprecursorprotein), BACE1 (behandlingsprotease β-secreatase 1) og peptidamyloid-β (Aβ) rigelig i OPC'er (figur 2F).
- Oligodendrocyt (OL) modning (~ 7-20 dage)
- På dag 15 udskiftes medier med OL-modningsmedium: Neurobasal-A-medium, 2% B27-supplement, 1 μM cAMP, 200 ng / ml T3-triiodothyronin og klemasin på 1 μM (se materialetabel). Skift mediet hver anden dag eller hver dag, hvis det er nødvendigt.
- Når celler når 90% sammenløb, opdeles ved et 1: 3-forhold op til 2 passager, eller indtil celledelingen sænkes væsentligt. Hvis OPC'er deler sig for hurtigt og når sammenløb på mindre end 3 dage, tilsættes Ara-C (se Materialetabel) i en koncentration på 2-5 μM i 1-3 dage. Aktiv spredning indikerer nedsat modningseffektivitet.
- Undersøg effektiviteten af oligodendroglial modning ved at vurdere ekspressionen af OL-markører, fx CLDN11, PLP1, MBP ved qPCR, IHC-farvning eller immunblotting. Den karakteristiske morfologi af meget komplekse strukturer (figur 2C) og ekspressionen af OL-markører (figur 2E) bør let detekteres på dag 28.
3. Co-dyrkning af humant inducerede neuroner (iN'er) og oligodendrocytprækursorceller (iOPC'er)
- iOPC-plettering (~ 3 dage)
- Plade iOPC'er på dag 14 ved en massefylde på 1 x 105 celler pr. brønd i en 24-brøndsplade (som beskrevet ovenfor i trin 2.2.4.) i OPC-differentieringsmedium (som beskrevet i trin 2.2.2.).
- iN-iOPC co-kultur oprettet
- På dag 15 løsnes de inducerede humane neuroner på trin 2 efter Puromycin-udvælgelsen (som beskrevet i trin 1.2.6.) med celleløsningsopløsning (se Materialetabel).
- Tilsæt neuroner på de dyrkede OPC'er, plettering ved såtætheden på 2 x 105 celler pr. Brønd i 24-brøndspladen med voksende OPC'er (fra trin 3.1.1). Brug co-kulturmediet indeholdende Neurobasal-A medium, 2% B27 supplement og 100 ng / ml T3 triiodothyronin. Skift mediet den næste dag og derefter hver anden dag bagefter. Hvis OPC'er spredes for hurtigt og når sammenløb på mindre end 3 dage, tilsættes Ara-C i en koncentration på 2-5 μM. Et repræsentativt billede af iNs og iOPC'er dyrket i samkultur efter 7 dage neuroner er vist i figur 3A.
- Brug frosne neuroner fremstillet som beskrevet ovenfor i trin 1.2.7 til samdyrkning med OPC'er. Plade fryser og optøer neuroner ved en højere densitet på 3 x 105 celler pr. Brønd.
- Efter dag 14-16 i co-kulturer kan synapsedannelsen i iNs observeres ved IHC-farvning af præ- og postsynaptiske markører, og på dag 21 skal den synaptiske puncta være rigelig (figur 3C), og neuronale aktiviteter kan registreres pålideligt.
- Fra dag 21 testes celler for OL-specifikke markører (for eksempel MBP og PLP1). På dag 28 observerer vi normalt fænomenet indkapsling af iN-axoner ved iOL-processer, mærket med IHC-farvning til specifikke markører (figur 3B; neurofilament NF til iN-axoner og MBP til iOPC-processer).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Direkte generering af humant inducerede neuroner fra humane pluripotente stamceller
Det er meget vigtigt, at de begyndende humane pluripotente stamceller udviser en høj grad af pluripotens for en vellykket generering af iN'er eller iOPC'er / iOL'er. Derfor bør celler farves for specifikke markører, såsom Oct4 og SOX2, inden en af induktionsprotokollerne beskrevet i dette manuskript (figur 1A påbegyndes). Humane H1-celler blev brugt til at opnå inducerede excitatoriske forhjerneneuroner efter den tidligere offentliggjorte protokol af Zhang et al. med nogle ændringer (figur 1C) 12,16,17,18. Her præsenterer vi en protokol, hvor iNs på dag 2 genbelægges i ren kultur på matrixopløsning (se Materialetabel), i mangel af noget feederlag: glia eller fibroblaster. Ud over de tidligere offentliggjorte protokoller observerer vi, at frysning af iNs på dag 2 ikke påvirker cellens levedygtighed væsentligt (~ 15% -20% celledød efter optøning). Rene neuroner i kultur vil begynde at udtrykke synapsin1 på dag 14-16 (figur 1D). Etablering af en ren neuronal kultur er meget vigtig, fordi visse faktorer, for eksempel den førende AD-risikofaktor ApoE, kan udtrykkes af celler i feederlaget, og dette kan forvirre resultaterne betydeligt.
iOPC-generering og iOL-modning forbedres ved DMSO-behandling
Her præsenterer vi en hurtig og effektiv protokol, der muliggør generering af iOPC'er på 2 uger og modne iOL'er på 4-5 uger (figur 2A). Vi udnyttede metoden til forbigående DMSO-behandling, som vi tidligere har udviklet, til at øge differentieringseffektiviteten for ES- og iPS-celler 29,30,31. DMSO-behandling beriger antallet af celler i den tidlige G1-fase for bedre signalintegration, hvilket favoriserer differentiering. Vi udførte den første behandling, før vi inducerede humane ES-celler til at generere NPC'er, og den anden behandling, før vi differentierede NPC'er til iOPC'er. Vi kan detektere specifikke OPC-markører (Olig2, CSPG4, NKX2.2 og PDGFRA) så tidligt som 2 uger efter plettering af ES H1-celler (figur 2B, E). iOPC-populationen på dette stadium er ret homogen, med >95% af cellerne positive for O4-farvning og et højt immunreaktivitetsniveau for andre markører (figur 2B). Efter starten af OL-modning på dag 15 kan vi typisk registrere specifikke OL-markører (MBP, O1, CLDN11 og PLP1) startende ved dag 28 (figur 2C, E). Udtrykket af disse fasespecifikke markører korrelerer med udviklingsforløbet af oligodendroglialceller og antyder et accelereret tempo, hvor NPC-markørerne gradvist går ned, OPC-markører topper omkring den anden uge og OL-markører hæver sig i den tredje uge (figur 2D, E)37. Bemærk, at denne modningsproces diversificerer cellepopulationerne. Delpopulationerne i kontinuum, der omfatter flere mellemstadier mellem OPC'er og modne myeliniserende oligodendrocytter, kan være til stede og tegne sig for varierende procentdel af de samlede celler, hvor mere modne celler dominerer på et senere tidspunkt.
Til sammenligning købte vi de meget refererede iOPC'er og modnede dem i iOL efter producentprotokollen. Vi testede ekspressionen af de ovennævnte markører i både vores iOPC- og iOL-præparater og i de celler, vi købte. Vi fastslog, at de celler, der blev genereret efter vores protokol, havde højere ekspression af alle de testede gener (figur 2E). Interessant nok, da vi testede de udskillede niveauer af to store isoformer af amyloid-β (Aβ40 og Aβ42) i iNs versus iOPC'er, bemærkede vi, at iOPC'er udskilte mere af begge fragmenter, men forholdet forblev det samme (figur 2F).
Samdyrkning af iNs og iOPC'er
Denne protokol er optimeret specifikt til co-dyrkning af iNs og iOPC'er og tillader vores realtidsovervågning af den intercellulære kommunikation mellem disse to celletyper i løbet af neural udvikling. De ideelle belægningstætheder for begge celletyper skal bestemmes med en række celletalsiditrering for at opnå korrekt differentiering (figur 3A). Efter 4 uger i samkulturer forventes iOPC'erne at blive tilstrækkeligt differentieret til OL'er, der er positive for specifikke markører såsom MBP og udvider processer til at ensheath axoner (figur 3B). Co-kultursystemet kan robust øge antallet af synapser, hvilket indikerer, at iOPC'erne giver en neuronal støtte gennem fysiske kontakter eller frigivelse af trofiske faktorer (figur 3C). Vi kan opretholde co-kulturerne i acceptabel helbredstilstand i op til 6 uger og observere, at synapsetallet og andre neuronale egenskaber plateau omkring den femte uge. Det skal bemærkes, at astrocytter og mikroglia ikke er til stede i vores præparater, og deres fravær kan dokumenteres ved at kontrollere ekspressionen af specifikke markører (figur 3D). IOPC'erne udtrykker et stort antal velkarakteriserede gener, der potentielt kan reagere på og formidle de aktivitetsafhængige signaler fra naboneuroner på en parakrin (f.eks. Neurotrofiner og metabolitter) og / eller en synaptisk måde (figur 3E og 3F).

Figur 1: Direkte generering af humaninducerede neuroner (iN'er) fra hPSC'er. (A) Flowdiagram over iN-generering. (B) Repræsentative lysfelt- og immunfluorescensbilleder af startkulturen af humane pluripotente stamceller (H1) for at bekræfte pluripotensen. Oct4 er vist med rødt og Sox2 i grønt. (C) Repræsentative lyse feltbilleder af iNs på dag 4 og dag 6. (D) Den karakteristiske morfologi for dendritisk arborisering og synapse puncta i iNs dyrket i ren kultur i 24 dage og farvet ved immunfluorescensfarvning til dendritisk markør Map2 og præsynaptisk markør Synapsin 1 (Syn1). Klik her for at se en større version af denne figur.

Figur 2: iOPC-generering og iOL-modning. (A) Flowdiagram over iOPC- og iOL-generering. (B) Repræsentative lysfelt- og immunfluorescensbilleder af iOPC'er på dag 15. Olig2 (pan-oligodendroglia markør) er vist med grønt, O4 (OPC markør) i rødt og DAPI i blåt. Billeddannelsen afslørede, at >95% af iOPC'erne er positive for O4 og 25% for Olig2. (C) Repræsentative lysfelt- og immunfluorescensbilleder af iOL'er på dag 28. MBP er vist med grønt, O1 med rødt og DAPI med blåt. (D) Ekspressionen af NPC-markøren PAX6 falder dramatisk i iOPC'er på dag 14 og sænkes yderligere til baggrund i OL'er på dag 28, hvilket indikerer en robust NPC-transdifferentiering og en høj grad af homogenitet i iOPC-populationen. (E) Tidsforløbsekspressionsprofilen for fælles OPC- og OL-markørgener i kulturer genereret af den beskrevne protokol uden (-DMSO) eller med (+DMSO) DMSO-inkubationstrinnet (trin 2.1.3 og 2.2.2), analyseret på forskellige tidspunkter. Til sammenligning blev kommercielle iOPC'er (se Materialetabel) modnet i henhold til producentens anvisninger, og både iOPC'er (iOPC-Tempo) eller iOL'er (iOL-Tempo) blev testet for de samme markører. Som forventet blev MBP (en moden oligodendrocytmarkør) ikke påvist (N.D.) i de tidlige stadier af differentiering i alle de testede iOPC'er. DMSO forbedrede effektiviteten af OPC-differentiering og OL-modning betydeligt. (F) Produktion og sekretion af Aβ40 og Aβ42 i rene iN- og iOPC-kulturer målt ved kommercielle ELISA-sæt (se materialetabel) på supernatant opnået fra rene iNs- og iOPC-kulturer både på dag 15 og normaliseret med celletal (begge ved densitet på 200.000 celler pr. brønd i en 24-brøndsplade). Klik her for at se en større version af denne figur.
Data i søjlediagrammer afbildes som gennemsnit ± SEM (n ≥ 3). Statistisk signifikans blev evalueret ved Student's t-test (*, p < 0,05; ***, s < 0,001); i (D) sammenlignet med NPC i litra E) sammenlignet med kontrol-iOPC-Tempo i (F) sammenlignet med iN.

Figur 3: Samkultur mellem iN'er og iOPC'er. (A) Repræsentativt lyst feltbillede af samkulturerede iN'er og iOPC'er på dag 7, der viser en passende tæthed for yderligere modning. (B) Repræsentativt immunfluorescensbillede af iNs og iOPC'er samdyrket i 28 dage. Axonal markør neurofilament NF er vist i grøn og oligodendrocytisk markør MBP i rødt. Til højre, et segment af iN-axon indkapslet af iOL-proces (MBP +). (C) Synapsedannelse analyseret i 4 uger gamle samkulturer. Celler blev farvet for Synapsin 1 (Syn1, grøn) og MAP2 (rød), og synaptisk puncta blev kvantificeret ved konfokal analyse af densitet langs de dendritiske segmenter som beskrevet17,18. (D) I vores co-kulturer af iNs og iOPC'er (7 dages co-kultur) er ekspressionen af astrocytmarkører, ALDHL1 og GFAP, minimal (øverst), og ekspressionen af microglia-markører, TMEM119, TREM2 og CD33, detekteres ikke (N.D.) af qPCR. Forureningen fra disse to gliacelletyper er således udelukket. (E) Cokulturering af iOPC med iN fører til dannelsen af neuron-OPC-synapser. Den fluorescensmærkede postsynaptiske markør PSD95-mCherry udtrykkes kun i OPC'er og viser et diffust mønster i enkeltkulturer (venstre), men aggregeres for at danne puncta i cokulturer (højre, angivet med pile; Tuj1, neuronal markør). (F) Ekspressionen af velkarakteriserede oligodendrogliale gener, der kan mærke og reagere på neuronale aktiviteter i de rene kulturer af iOPC'er på dag 14. Klik her for at se en større version af denne figur.
Data i søjlediagrammer afbildes som middel ± SEM (n ≥ 3). Statistisk signifikans blev evalueret ved Student's t-test (**, p < 0,005; ***, s < 0,001); i (C) sammenlignet med betingelsen om ingen OPC i (D) sammenlignet med primære astrocytter i toppanelet.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Ud over den fysiske og metaboliske støtte til at stabilisere synapsestrukturerne og lette saltsignalledningen ved myelinering, kan oligodendrocytlinjeceller forme neuronalt aktivitetsmønster via hurtige og dynamiske krydssamtaler med neuroner 5,6,7. Mens oligodendroglial-reaktionerne i AD-patologi oprindeligt blev betragtet som blot sekundære i forhold til inflammation og oxidative belastninger, er der nu lovende beviser, der hævder, at kompromitteret myelinintegritet er en tidlig patogen begivenhed forud for udseendet af Aβ-aggregering og tau-hyperphosphorylering9. Desuden er reparation af myelinering gennem selvfornyelse af OPC'er særligt sårbar iAD 38, en proces, der i høj grad afhænger af neuronale aktiviteter. At forstå mekanismen til at understøtte sund neuron-oligodendrocytsignalering repræsenterer således en glimrende mulighed for at identificere nye terapeutiske mål.
Den enkelte transkriptionsfaktor Ngn2-protokol er en af de mest anvendte teknikker til generering af stamcelleafledte humane neuroner, og de procedurer, der er skitseret her, er yderligere forbedringer til opnåelse af rene neuronale kulturer. Vores iOPC/iOL-protokol har en induktionsperiode, der er kortere end de tidligere offentliggjorte undersøgelser (4 til 24 uger), med et robust udbytte og renhed, der kan sammenlignes med andre almindeligt anvendte protokoller 19,20,22,23,24,25,26,27,28 . Vores protokol introducerer trinvis differentiering af ES-celler til NPC'er, OPC'er og endelig oligodendrocytter ved karakteriserede mønstersignaler og genererer funktionelle celler, der kan bruges til at studere reguleringen af myelineringshomeostase og reparere in vitro eller in vivo (f.eks. ved indkapsling i shiverer musemodellen) som beskrevet i det foregående arbejde. Forbedringen i vores protokol fremmes i høj grad af DMSO-inkubationen, som aktiverer retinoblastomproteinet og forlænger G1-fasen af cellecyklussen for bedre at integrere stimuli for rettet differentiering og forbedrer også terminal differentiering i funktionelle derivater29,30. Endelig forkorter brugen af clemastin, en muskarin og antihistaminisk forbindelse, der er identificeret gennem lægemiddelskærme til remyelineringsterapi33, yderligere oligodendrocytmodningen, som observeret i iPS-celleforberedelse og levende dyr 7,28.
Teknikkens begrænsninger ligger hovedsageligt i den iboende uoverensstemmelse mellem de forenklede in vitro-indstillinger og in vivo-miljøerne i hjernen; Denne uoverensstemmelse fører til en rabat i det fulde udviklingspotentiale på de avancerede stadier for individuelle hjernecelletyper. For iN'er var nylige undersøgelser i stand til at opretholde kulturerne i god synaptisk sundhed i en betydelig lang periode, men afslørede stadig en vis relativ umodenhed manifesteret som reducerede rygsøjlelignende strukturer og nedsat spontan synaptisk transmission i "gamle" iN-kulturer (selv de 25 måneder gamle)39. Mens iOPC'er er blevet reproducerbart vist til myelinataxoner in vivo efter transplantation i transgene musehjerner, udgør in vitro myelineringsassays med elektronmikroskopisk evaluering stadig en teknisk udfordring med utilfredsstillende effektivitet for næsten alle offentliggjorte protokoller19,28, inklusive denne. Derfor forventes vores neuron-OPC-co-kultursystem ikke trofast at efterligne hjernens aldringsproces såvel som den sene fase af AD-patologi. Det er snarere unikt klar til at adskille de udførlige intercellulære interaktioner mellem neuroner og OPC'er eller oligodendrocyt i det tidlige stadium, som er uafhængige af myelinering og alligevel grundlæggende for korrekt neural udvikling og sygdomspatogenese.
Hver af de tre procedurer, der er beskrevet her, har sine c ritical trin inden for protokollen og kan kræve ændringer og fejlfinding. For iN-protokollen er der to kritiske trin: puromycinvalg (trin 1.2.5) og belægningstæthed (trin 1.2.6). Ufuldstændig fjernelse af undertransduceret celle resulterer i forurening af dårligt differentierede celler og kompromitterer neuronal overlevelse og funktioner. Ændringerne for stærkere puromycinselektion med højere koncentration og længere inkubation som beskrevet i trin 1.2.5 skal overvejes. Den passende belægningstæthed bør bestemmes ved titrering for hver pluripotent cellelinje, da lav densitet fører til sammenbrud af kulturerne, og høj densitet tilskynder til celleaggregering og hæmmer neuronal vækst. For iOPC/iOL-protokollen er de to kritiske trin kontrol af celleproliferation i OPC-differentiering (trin 2.2.3) og belægningstæthed for OL-modning (trin 2.2.4 og 2.3.2). Tilgroningen af differentierende NPC'er signalerer en dårlig reaktion på OPC-differentieringsstimuli og skal dæmpes ved en passende dosis af Ara-C-behandling (inden for det angivne interval). Mens der pletteres OPC'er til modning, foretrækkes et lavere interval af celletæthed her, da den sparsomme fordeling kan lette induktionen af en fysiologisk morfologi af komplekse strukturer (som vist i figur 2C). For iN-iOPC co-kulturprotokol vil vi gerne henlede opmærksomheden på det kritiske trin i plettering med en passende tæthed for begge celletyper (trin 3.1.1 og 3.2.2). Specifikt kan iN'erne muligvis ikke fastgøres godt til overfladen mellem voksende OPC'er og har tendens til at løsne sig først, når kulturen når sammenløb. Det optimale forhold skulle bestemmes ved at titrere cellenumrene.
Samlet set er denne reduktionistiske tilgang, der findes i vores protokoller, et kraftfuldt værktøj til at dissekere de specifikke heterocellulære interaktioner fra den menneskelige hjernes iboende kompleksitet og tjene til at afdække oligodendroglial biologi i sundhed og i AD. Betydningen i forhold til eksisterende metoder er således efter vores mening ret tydelig. En yderligere nytte af de metoder, der er udviklet her til de fremtidige applikationer, er cellebaseret terapi til demyeliniserende tilstande, såsom post-strålebehandling40 og rygmarvsskade41,42. Desuden kan den høje kapacitetskapacitet i dette stamcellebaserede system også udnyttes i større skala til at screene biblioteker af små molekyler for forbindelser, der kan beskytte eller genoprette den fysiologiske status for neuroner, OPC'er, oligodendrocytter og deres interaktioner. Således mener vi, at de protokoller, der er beskrevet her, vil lette det fremtidige arbejde med at udvikle bedre modelleringsværktøjer og effektive behandlinger for AD og andre neurodegenerative lidelser.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har intet at afsløre.
Acknowledgments
Dette arbejde blev støttet af tilskud fra National Institutes of Health (R00 AG054616 til Y.A.H. og T32 GM136566 til K.C.), Stanford University School of Medicine og et Siebel Fellowship (tildelt SC). Y.A.H. er en GFL Translational Professor fra Center for Translational Neuroscience i Brown Institute for Translational Sciences.
Materials
| Name | Company | Catalog Number | Comments |
| Accutase | STEMCELL Technologies | 7920 | |
| B27 supplement | ThermoFisher | 17504044 | |
| bFGF | ThermoFisher | PHG 0266 | |
| cAMP | MilliporeSigma | A9501 | |
| Clemastine | MilliporeSigma | SML0445 | |
| DMEM/F12 medium | STEMCELL Technologies | 36254 | |
| DMSO | ThermoFisher | D12345 | |
| Doxycycline | MilliporeSigma | D3072 | |
| Fetal Bovine Serum | ScienCell | 10 | |
| H1 human ES cells | WiCell | WA01 | |
| Matrigel | Corning | 354234 | |
| mTeSR plus | STEMCELL Technologies | 5825 | |
| N2 supplement | ThermoFisher | 17502001 | |
| Neurobasal A medium | ThermoFisher | 10888-022 | |
| Non Essential Amino Acids | ThermoFisher | 11140-050 | |
| PDGF-AA | R&D Systems | 221-AA-010 | |
| PEI | VWR | 71002-812 | |
| pMDLg/pRRE | Addgene | 12251 | |
| Polybrene | MilliporeSigma | TR-1003-G | |
| pRSV-REV | Addgene | 12253 | |
| Puromycin | ThermoFisher | A1113803 | |
| ROCK Inhibitor Y-27632 | STEMCELL Technologies | 72302 | |
| SAG | Tocris | 4366 | |
| STEMdiff Neural Progenitor Freezing Media | STEMCELL Technologies | 5838 | |
| STEMdiff SMADi Neural Induction Kit | STEMCELL Technologies | 8581 | |
| T3 triiodothyronine | MilliporeSigma | T6397 | |
| Tempo-iOlogo: Human iPSC-derived OPCs | Tempo BioScience | SKU102 | |
| TetO-Ng2-Puro | Addgene | 52047 | |
| VSV-G | Addgene | 12259 |
References
- Pelvig, D. P., Pakkenberg, H., Stark, A. K., Pakkenberg, B. Neocortical glial cell numbers in human brains. Neurobiology of Aging. 29 (11), 1754-1762 (2008).
- Barres, B. A. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 60 (3), 430-440 (2008).
- De Strooper, B., Karran, E. The cellular phase of Alzheimer's disease. Cell. 164 (4), 603-615 (2016).
- Monje, M. Myelin plasticity and nervous system function. Annual Review of Neuroscience. 41, 61-76 (2018).
- Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., Bergles, D. E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nature Neuroscience. 21 (5), 696-706 (2018).
- Gibson, E. M., et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science. 344 (6183), 1252304 (2014).
- Pan, S., Mayoral, S. R., Choi, H. S., Chan, J. R., Kheirbek, M. A. Preservation of a remote fear memory requires new myelin formation. Nature Neuroscience. 23 (4), 487-499 (2020).
- Thornton, M. A., Hughes, E. G. Neuron-oligodendroglia interactions: Activity-dependent regulation of cellular signaling. Neuroscience Letters. 727, 134916 (2020).
- Ettle, B., Schlachetzki, J. C. M., Winkler, J. Oligodendroglia and myelin in neurodegenerative diseases: more than just bystanders. Molecular Neurobiology. 53 (5), 3046-3062 (2016).
- Essayan-Perez, S., Zhou, B., Nabet, A. M., Wernig, M., Huang, Y. A. Modeling Alzheimer's disease with human iPS cells: advancements, lessons, and applications. Neurobiology of Disease. 130, 104503 (2019).
- Li, L., et al. GFAP mutations in astrocytes impair oligodendrocyte progenitor proliferation and myelination in an hiPSC model of Alexander disease. Cell Stem Cell. 23 (2), 239-251 (2018).
- Lin, Y. T., et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer's disease phenotypes in human iPSC-derived brain cell types. Neuron. 98 (6), 1294 (2018).
- TCW, J., et al. Cholesterol and matrisome pathways dysregulated in human APOE ε4 glia. bioRxiv. , (2019).
- Ang, C. E., Wernig, M.
- Penney, J., Ralvenius, W. T., Tsai, L. H. Modeling Alzheimer's disease with iPSC-derived brain cells. Molecular Psychiatry. 25 (1), 148-167 (2020).
- Zhang, Y., et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron. 78 (5), 785-798 (2013).
- Huang, Y. A., Zhou, B., Nabet, A. M., Wernig, M., Sudhof, T. C. Differential signaling mediated by ApoE2, ApoE3, and ApoE4 in human neurons parallels Alzheimer's Disease risk. Journal of Neuroscience. 39 (37), 7408-7427 (2019).
- Huang, Y. A., Zhou, B., Wernig, M., Sudhof, T. C. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Abeta Secretion. Cell. 168 (3), 427-441 (2017).
- Yang, N., et al. Generation of oligodendroglial cells by direct lineage conversion. Nature Biotechnology. 31 (5), 434-439 (2013).
- Douvaras, P., et al. Efficient generation of myelinating oligodendrocytes from primary progressive multiple sclerosis patients by induced pluripotent stem cells. Stem Cell Reports. 3 (2), 250-259 (2014).
- Lee, E. H., Park, C. H. Comparison of reprogramming methods for generation of induced-oligodendrocyte precursor cells. Biomolecules & Therapeutics (Seoul). 25 (4), 362-366 (2017).
- Ehrlich, M., et al. Rapid and efficient generation of oligodendrocytes from human induced pluripotent stem cells using transcription factors. Proceedings of the National Academy of Sciences of the United States of America. 114 (11), 2243-2252 (2017).
- Rodrigues, G. M. C., et al. Defined and scalable differentiation of human oligodendrocyte precursors from pluripotent stem cells in a 3D culture system. Stem Cell Reports. 8 (6), 1770-1783 (2017).
- Hu, B. Y., Du, Z. W., Li, X. J., Ayala, M., Zhang, S. C. Human oligodendrocytes from embryonic stem cells: conserved SHH signaling networks and divergent FGF effects. Development. 136 (9), 1443-1452 (2009).
- Izrael, M., et al. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Molecular and Cellular Neuroscience. 34 (3), 310-323 (2007).
- Yamashita, T., et al. Differentiation of oligodendrocyte progenitor cells from dissociated monolayer and feeder-free cultured pluripotent stem cells. PLoS One. 12 (2), 0171947 (2017).
- Wang, S., et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell. 12 (2), 252-264 (2013).
- Chanoumidou, K., Mozafari, S., Baron-Van Evercooren, A., Kuhlmann, T. Stem cell derived oligodendrocytes to study myelin diseases. Glia. 68 (4), 705-720 (2020).
- Chetty, S., et al. A simple tool to improve pluripotent stem cell differentiation. Nature Methods. 10 (6), 553-556 (2013).
- Li, J., et al. A transient DMSO treatment increases the differentiation potential of human pluripotent stem cells through the Rb family. PLoS One. 13 (12), 0208110 (2018).
- Sambo, D., Li, J., Brickler, T., Chetty, S. Transient treatment of human pluripotent stem cells with DMSO to promote differentiation. Journal of Visualized Experiments: JoVE. (149), (2019).
- Douvaras, P., Fossati, V. Generation and isolation of oligodendrocyte progenitor cells from human pluripotent stem cells. Nature Protocols. 10 (8), 1143-1154 (2015).
- Mei, F., et al. Micropillar arrays as a high-throughput screening platform for therapeutics in multiple sclerosis. Nature Medicine. 20 (8), 954-960 (2014).
- Madhavan, M., et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nature Methods. 15 (9), 700-706 (2018).
- Zhang, Y., et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 89 (1), 37-53 (2016).
- Grubman, A., et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer's disease reveals cell-type-specific gene expression regulation. Nature Neuroscience. 22 (12), 2087-2097 (2019).
- Goldman, S. A., Kuypers, N. J. How to make an oligodendrocyte. Development. 142 (23), 3983-3995 (2015).
- Behrendt, G., et al. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia. 61 (2), 273-286 (2013).
- Patzke, C., et al. Neuromodulator signaling bidirectionally controls vesicle numbers in human synapses. Cell. 179 (2), 498-513 (2019).
- Piao, J., et al. Human embryonic stem cell-derived oligodendrocyte progenitors remyelinate the brain and rescue behavioral deficits following radiation. Cell Stem Cell. 16 (2), 198-210 (2015).
- Keirstead, H. S., et al. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. Journal of Neuroscience. 25 (19), 4694-4705 (2005).
- Kim, D. S., et al. Rapid generation of OPC-like cells from human pluripotent stem cells for treating spinal cord injury. Experimental & Molecular Medicine. 49 (7), 361 (2017).
Tags
Neurovidenskab udgave 165 inducerede pluripotente stamceller iPS-celler humane embryonale stamceller ES-celler neuroner oligodendrocytter oligodendrocytprogenitorceller OPC'er Alzheimers sygdom amyloid-beta-peptider synapserErratum
Formal Correction: Erratum: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions
Posted by JoVE Editors on 12/29/2020.
Citeable Link.
An erratum was issued for: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions. The Representative Results section has been updated.
Figure 3 was updated from:
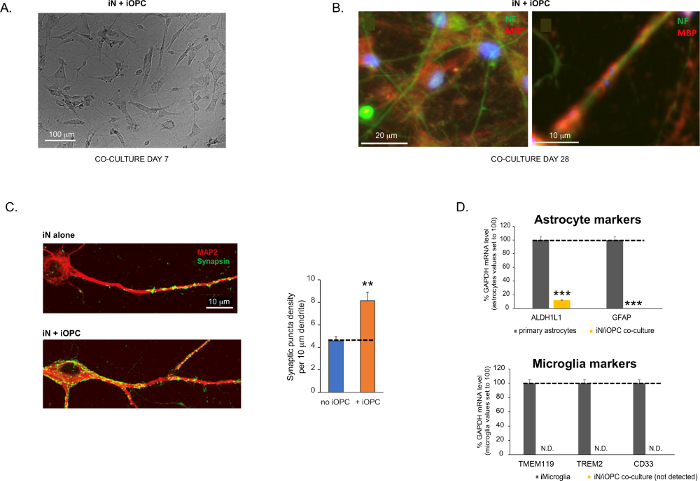
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. Please click here to view a larger version of this figure.
to:
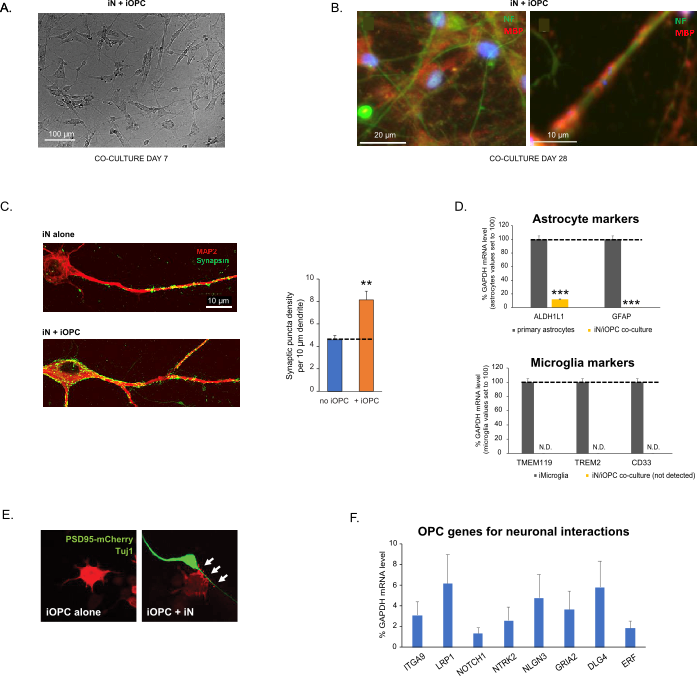
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. (E) Coculturing iOPC with iN leads to the formation of neuron-OPC synapses. The fluorescence-tagged post-synaptic marker PSD95-mCherry is expressed only in OPCs, and display a diffuse pattern in single cultures (left) but aggregate to form puncta in cocultures (right, indicated by arrows; Tuj1, neuronal marker). (F) The expression of well-characterized oligodendroglial genes that can sense and respond to neuronal activities in the pure cultures of iOPCs at Day 14. Please click here to view a larger version of this figure.
The fourth paragraph was updated from:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D).
to:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D). The iOPCs express a good number of well-characterized genes that can potentially respond to and mediate the activity-dependent signals from neighboring neurons, in a paracrine (e.g. neurotrophins and metabolites) and/or a synaptic manner (Figure 3E and 3F).
