

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Нейронно-глиальные взаимодействия при нейродегенерации недостаточно изучены из-за неадекватных инструментов и методов. Здесь мы описываем оптимизированные протоколы для получения индуцированных нейронов, клеток-предшественников олигодендроцитов и олигодендроцитов из плюрипотентных стволовых клеток человека и приводим примеры значений этих методов в понимании специфических для типа клеток вклада в болезнь Альцгеймера.
Abstract
При болезни Альцгеймера (БА) и других нейродегенеративных расстройствах олигодендроглиальная недостаточность является общей ранней патологической особенностью, но как она способствует развитию и прогрессированию заболевания, особенно в сером веществе мозга, остается в значительной степени неизвестным. Дисфункция клеток линии олигодендроцитов характеризуется недостатками миелинизации и нарушением самообновления клеток-предшественников олигодендроцитов (ОПК). Эти два дефекта вызваны, по крайней мере частично, нарушением взаимодействия между нейронами и олигодендроцитами вдоль накопления патологии. ОПК дают начало миелинизации олигодендроцитов во время развития ЦНС. В зрелой коре головного мозга OPC являются основными пролиферативными клетками (составляющими ~ 5% от общего количества клеток мозга) и контролируют образование новых миелинов в зависимости от нейронной активности. Такие нейронно-олигодендроцитарные коммуникации значительно недоизучены, особенно в контексте нейродегенеративных состояний, таких как БА, из-за отсутствия соответствующих инструментов. В последние годы наша группа и другие добились значительного прогресса в улучшении доступных в настоящее время протоколов для генерации функциональных нейронов и олигодендроцитов индивидуально из плюрипотентных стволовых клеток человека. В этой рукописи мы описываем наши оптимизированные процедуры, включая создание системы кокультуры для моделирования нейронно-олигодендроцитарных связей. Наши иллюстративные результаты свидетельствуют о неожиданном вкладе OPCs / олигодендроцитов в амилоидоз мозга и целостность синапсов и подчеркивают полезность этой методологии для исследований БА. Этот редукционистский подход является мощным инструментом для препарирования специфических гетероклеточных взаимодействий из присущей сложности внутри мозга. Ожидается, что протоколы, которые мы описываем здесь, облегчат будущие исследования олигодендроглиальных дефектов в патогенезе нейродегенерации.
Introduction
Клетки линии олигодендроцитов, включая клетки-предшественники олигодендроцитов (OPCs), миелинирующие олигодендроциты и переходные типы между ними, составляют основную группу клеток головного мозга человека1, которые активно участвуют во многих критических функциях для правильной работы и поддержания нашей центральной нервной системы на протяжении всего нервного развития и старения 2,3,4 . В то время как олигодендроциты хорошо известны тем, что производят миелин для облегчения передачи активности нейронов и поддержки здоровья аксонов в белом веществе, OPC в изобилии (~ 5%) в сером веществе, где миелинизация скудна, и выполняют зависящие от активности сигнальные функции для управления поведением обучения и формированием памяти 5,6,7,8 . Как функционируют олигодендроглиальные клетки и дисфункция в патогенезе болезни Альцгеймера (БА) и других возрастных нейродегенеративных состояний, было недостаточно изучено9. Недостатки соответствующей модельной системы и недостатки в общих знаниях, позволяющих направлять экспериментальный путь вперед, являются основными причинами этого разрыва.
В свете последних прорывов в получении клеток человеческого мозга из плюрипотентных стволовых клеток, включая эмбриональные стволовые (ES) и индуцированные плюрипотентные стволовые (iPS) клетки, такие клеточные модели в сочетании с современными инструментами редактирования генов стали надежными инструментами для обработки сложной связи клеточных взаимодействий в мозге и способны демонстрировать специфические для человека проявления заболеваний10, 11. Учитывая, что отдельные типы клеток мозга могут проявлять различные и даже противоречивые эффекты перед лицом одних и тех же условий, способствующих АД12,13, эта методология стволовых клеток однозначно предлагает информацию, специфичную для типа клеток, которая ранее была пропущена с использованием установленных моделей in vivo или in vitro, которые обеспечивают только агрегированные показания из коллекций типов клеток мозга. В последнее десятилетие было разработано большое количество надежных протоколов для генерации нейронов человека из трансдифференцировки ES/iPS-клеток или прямого преобразования из других терминально дифференцированных типов клеток (например, фибробластов)14,15. В частности, применение ключевых нейрогенных факторов транскрипции (например, нейрогенина 2, Ngn2)16 к плюрипотентным стволовым клеткам человека может генерировать однородную популяцию хорошо охарактеризованных типов нейрональных клеток для чистых культур без необходимости кокультурирования с глиальными клетками 12,17,18. Для индуцированных человеческих олигодендроцитов существует несколько опубликованных протоколов, которые могут генерировать функциональные клетки, очень похожие на их первичные аналоги, с широким диапазоном эффективности и спроса во времени и ресурсах 19,20,21,22,23,24,25,26,27,28 . На сегодняшний день ни один из этих протоколов не был применен для исследования того, как олигодендроглиальные клетки реагируют и влияют на патогенез БА.
Здесь мы описываем наши улучшенные протоколы для одиночных и смешанных культур индуцированных человеком нейронов (iNs) и OPCs / олигодендроцитов (iOPCs / iOL). Протокол iN, описанный здесь, основан на широко используемом подходеNgn2 16 и имеет дополнительную особенность быть свободным от глии. Полученные iNs однородны и очень напоминают кортикальный слой 2/3 возбуждающих нейронов, с характерной пирамидальной морфологией, паттерном экспрессии генов и электрофизиологическими особенностями17,18 (Рисунок 1). Чтобы преодолеть некоторые из фундаментальных барьеров в направленной дифференцировке плюрипотентных стволовых клеток, мы разработали простой и эффективный метод предварительного лечения низкими дозами диметилсульфоксида (DMSO)29,30 и сообщили о повышенной склонности человеческих ES/iPS-клеток к трансдифференциации в iOPCs и iOL31, основанный на широко адаптированном протоколе Douvaras и Fossati32. . Мы еще больше упростили протокол и включили надежное соединение, способствующее дифференцировке, клемастин 7,33,34, для ускорения процесса созревания олигодендроглии. В результате (рисунок 2) iOPC могут быть сгенерированы за 2 недели (~95% положительных для маркера O4) и iOL за четыре недели (экспрессируя зрелые маркеры MBP и PLP1). Интересно, что мы обнаружили, что только iOPCs секретируют значительное количество β амилоида (Aβ), что согласуется с независимыми транскриптомными данными, показывающими обильную экспрессию белка-предшественника амилоида (APP) и обработку протеазы β-секретазы (BACE1) в клетках линии олигодендроцитов35,36. Кроме того, наша система кокультуры iN-iOPC способствует оболваниванию аксонов MBP-положительными процессами iOL и обеспечивает значительную поддержку образования синапсов (рисунок 3). Таким образом, протоколы, которые мы подробно описали ниже, имеют технические и биологические преимущества по сравнению с ранее каталогизированными методами совместного культивирования нейронов и олигодендроглии и имеют перспективу в лучшем моделировании нейродегенерации при БА.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Индукция нейронов человека из плюрипотентных стволовых клеток человека
- Препарат лентивируса (~5 дней, подробный протокол, как описано ранее16)
- Пластина ~ 1 миллион HEK293T клеток в каждой колбе T75, чтобы иметь их ~ 40% слива при выполнении трансфекции. Трансфектируйте их плазмидами, экспрессирующими тетрациклин-индуцируемый Ngn2 и пуромицин-резистентный ген (PuroR; под тем же контролем промотора TetO), rtTA и тремя вспомогательными плазмидами pRSV-REV, pMDLg/pRRE и VSV-G (12 мкг лентивирусной векторной ДНК и 6 мкг каждой из хелперных плазмидных ДНК). Приготовьте не менее трех колб на препарат лентивируса. Используйте PEI для трансфекции в соответствии с инструкцией производителя. Смените носитель через 16 ч и выбросьте.
- Собирают высвобожденные вирусные частицы путем сбора питательных сред каждый день и заменяют свежими средами в течение 3 дней. Объедините собранные среды, содержащие вирусные частицы, для очистки. Фильтруйте вирус через фильтр 0,22 мкм и центрифугу при 49 000 х г в течение 90 мин. Повторно суспендируют гранулу в соответствующем объеме PBS-глюкозы (~150 мкл).
- Индукция нейронов (~5 дней)
ПРИМЕЧАНИЕ: Этот протокол индукции (рисунок 1А; блок-схема) является высокоэффективным как для iPS, так и для ES клеток валидированной плюрипотентности (которые могут быть проанализированы иммуногистохимическим окрашиванием хорошо охарактеризованных маркеров плюрипотентности; Рисунок 1B).- Используйте коммерчески доступные H1 человеческие ES-клетки при прохождении 52 (см. Таблицу материалов). Культивируйте клетки на внеклеточном матричном растворе, покрытом 6-луночными пластинами (~0,5 мг матричного раствора на 6-луночную пластину; см. Таблицу материалов) с использованием среды для поддержания клеток ES (см. Таблицу материалов) и инкубируйте пластины при 37 °C с 5% CO2.
- На 2-й день отделяют ЭС-клетки (80% сливающиеся) с 1 мл раствора клеточной отслойки (см. Таблицу материалов) и инкубируют при комнатной температуре в течение 10 мин. Перенесите клетки в трубку; промыть лунку 2 мл среды и соединить в одной трубе. Центрифугу при 300 х г в течение 5 мин, повторно суспендируют гранулу в среде и выкладывают ячейки на покрытые матрицей 6-луночные пластины с плотностью посева 1 х 105 ячеек на лунку.
- На 1-й день добавляют лентивирусы, экспрессирующие Ngn2 плюс PuroR и rtTA вместе с полибреном (8 мкг/мл) к клеткам ES в свежей среде поддержания клеток ES (см. Таблицу материалов). Точное количество вирусов должно определяться фактическими титрами или титрованием. Обычно мы добавляем 5 мкл каждого вируса на скважину в 6-луночную пластину.
- На 0-й день добавляют доксициклин (2 мкг/мл, для активации экспрессии Ngn2) в среду DMEM-F12 с добавкой N2 без морфогенов.
- На 1-й день добавляют Пуромицин в свежую среду DMEM-F12 плюс N2 и доксициклин до конечной концентрации 1 мкг/мл среды. Отбирают трансдуцированные клетки в Пуромицине в течение не менее 24 ч. Для адекватного удаления недотрансдуцированных клеток может потребоваться более высокая концентрация Пуромицина (до 5 мкг/мл) и более длительный период отбора (до 48 ч).
- На 2-й день отсоедините дифференцирующие нейроны раствором для отсоединения клеток (см. Таблицу материалов) и повторно нанесите их на 24-луночные пластины (от 80 000 до 200 000 клеток / лунка), покрытые матричным раствором (см. Таблицу материалов), и сохраните их в среде NBA / B27 без доксициклина. Плотность посева имеет решающее значение.
- На этом этапе отсоединенные нейроны могут быть заморожены в специализированной коммерческой морозильной среде (см. Таблицу материалов) и храниться в жидком азоте до 3 месяцев. Чистые нейроны могут быть покрыты, что составляет типичный ~ 15-20% гибели клеток после оттепели, культивироваться отдельно или совместно культивироваться с другими типами клеток мозга (см. шаг 3.2.3. для совместного культивирования с OPC).
- Культивируйте чистые iNs на пластинах, покрытых растворами на основе внеклеточного матрикса в соответствии с инструкциями производителя (см. Таблицу материалов). Характерная пирамидальная морфология должна быть очевидна к 4-му дню (и 6-му дню; Рисунок 1С). Образование синапсов может быть обнаружено уже на 14-16 день и заметно на 24-й день путем иммуногистохимического окрашивания стандартными пре- и постсинаптическими маркерами. (Рисунок 1D; помечен предсинаптическим маркером Synapsin 1 и дендритным маркером Map2).
2. Индукция клеток-предшественников олигодендроцитов человека (OPCs) из плюрипотентных стволовых клеток и созревание олигодендроцитов
- Генерация нейронных клеток-предшественников (NPC): однослойный протокол (~7 дней). Блок-схему см. на рисунке 2A .
- Культивируйте H1 человеческие ES-клетки, как описано ранее (см. шаг 1.2.1.), и трансдифференцируйте их в нейронные клетки-предшественники (NPC) с помощью установленного подхода, называемого двойным SMADi, с ингибиторами малых молекул для нескольких сигнальных путей. Здесь мы используем широко распространенный коммерческий комплект и следуем монослойному протоколу, предоставленному производителем (см. Таблицу материалов).
- В день -1 пластина 0,5–1 х 106 ячеек на скважину в 6-луночной пластине, покрытой раствором с пониженным коэффициентом роста (см. Таблицу материалов; ~0,5 мг матричного раствора на 6-луночную пластину) средой для поддержания клеток ES (см. Таблицу материалов). Этот матричный раствор с пониженным фактором роста используется для покрытия всех пластин, которые будут использоваться на следующих этапах.
- На 0-й день обрабатывают клетки в течение 24 ч поддерживающей средой для клеток ES (см. Таблицу материалов), дополненной 2% ДМСО.
- На 1–6 день замените полную среду теплой (37 °C) нейронной индукционной средой, содержащей ингибиторы SMAD из коммерческого набора (см. Таблицу материалов). Если клетки делятся и достигают слияния до 7-го дня, переведите их к плотности посева 0,5–1 х 106, как описано ранее на этапе 2.1.2.
- На 7-й день проход NPC с использованием клеточного отслоения раствора (см. Таблицу материалов) и пластины при плотности посева 1–2 х 105 клеток/лунку из 24-луночной пластины.
- Анализ эффективности дифференцировки иммуногистохимическим (IHC) окрашиванием на отсутствие маркера плюрипотентности, например OCT4, и наличие маркеров NPC, таких как PAX6, Nestin и Sox1.
- На этом этапе отсоединенные NPC могут быть заморожены в специализированной коммерческой морозильной среде NPC (см. Таблицу материалов) и храниться в жидком азоте до 3 месяцев. После замораживания и оттаивания на один раз NPC все еще сохраняют мультипотентность, чтобы дать начало нейронам, астроцитам и OPC с надежными протоколами.
- Генерация клеток-предшественников олигодендроцитов (OPC) (~7 дней). Схему блок-схемы см. на рисунке 2A .
- На 7-й день проходят NPC с помощью раствора для отсоединения клеток (см. Таблицу материалов) и обкладывают их с плотностью посева 1–2 х 105 клеток на лунку в 24-луночной пластине в теплой (37 °C) нейронной индукционной среде плюс ингибиторы SMAD из коммерческого набора (см. Таблицу материалов).
- На 8-й день готовят раствор 1% ДМСО в дифференцировочной среде OPC и обрабатывают покрытые NPC в течение 24 ч. Дифференцирующая среда OPC состоит из: среды DMEM/F12, 1% добавки N2, 1% добавки B27, bFGF при 20 нг/мл, SAG при 1 мкМ, PDGF-AA при 10 нг/мл (см. Таблицу материалов).
- На 9-й день замените носитель свежей средой дифференциации OPC без ДМСО. Кормите клетки через день до 15-го дня. Если клетки достигают слияния до 15-го дня, переведите их к плотности посева 1–2 х 105 клеток на лунку, как описано на этапе 2.2.1.
- На 14-й день пластинчатые OPC в среде дифференцировки OPC при плотности 1–2 х 105 клеток/лунка в 24-луночной пластине.
- На этом этапе (день 15) проверяют клетки на наличие OPC-специфических маркеров путем окрашивания IHC или qPCR (например, O4, Olig1/2, CSPG4/Ng2, NKX2.2, PDGFRa; Рисунок 2B) и за отсутствие маркеров NPC (Pax6 или Nestin; Рисунок 2D). Мы обычно обнаруживаем иммунореактивность O4 в более чем 95% клеток на 15-й день. Особое значение для болезни Альцгеймера имеет экспрессия APP (белок-предшественник амилоида), BACE1 (обработка протеазы β-секреатазы 1) и пептида амилоид-β (Aβ) в изобилии содержится в OPCs (рисунок 2F).
- Созревание олигодендроцитов (OL) (~7–20 дней)
- На 15-й день замените среду созревания OL: нейробазаль-А, 2% добавки B27, 1 мкМ цАМФ, 200 нг/мл T3 трийодтиронина и клемастина 1 мкм (см. Таблицу материалов). Меняйте среду через день или каждый день, если это необходимо.
- Когда клетки достигают 90% слияния, расщепляются в соотношении 1:3 до 2 проходов или до тех пор, пока деление клеток существенно не замедлится. Если ОПК делятся слишком быстро и достигают слияния менее чем за 3 дня, добавляют Ара-С (см. Таблицу материалов) в концентрации 2–5 мкМ в течение 1–3 дней. Активное пролиферация указывает на снижение эффективности созревания.
- Изучить эффективность созревания олигодендроглии путем оценки экспрессии маркеров OL, например, CLDN11, PLP1, MBP с помощью qPCR, окрашивания IHC или иммуноблоттинга. Характерная морфология очень сложных структур (рисунок 2C) и экспрессия маркеров OL (рисунок 2E) должны быть легко обнаружены к 28-му дню.
3. Совместное культивирование индуцированных человеком нейронов (iNs) и клеток-предшественников олигодендроцитов (iOPCs)
- Покрытие iOPC (~3 дня)
- Пластинчатые iOPCs на 14-й день при плотности 1 х 105 ячеек на скважину в 24-луночной пластине (как описано выше на стадии 2.2.4.) в среде дифференцировки OPC (как описано в стадии 2.2.2.).
- Настройка совместной культуры iN-iOPC
- На 15-й день отделите индуцированные нейроны человека на этапе 2-го дня после отбора пуромицина (как описано в шаге 1.2.6.) раствором для отслоения клеток (см. Таблицу материалов).
- Добавляют нейроны на культивируемые OPC, покрывая при плотности посева 2 х 105 клеток на лунку в 24-луночной пластине с растущими OPC (из шага 3.1.1). Используйте кокультурную среду, содержащую среду Neurobasal-A, 2% добавку B27 и 100 нг/мл T3 трийодтиронина. Меняйте среду на следующий день, а затем через день. Если ОПК размножаются слишком быстро и достигают слияния менее чем за 3 дня, добавляют Ара-С в концентрации 2–5 мкМ. Репрезентативное изображение iNs и iOPC, выращенных в совместной культуре через 7 дней нейронов, показано на рисунке 3A.
- Используйте замороженные нейроны, приготовленные, как описано выше на этапе 1.2.7, для совместного культивирования с OPC. Пластинчатые замораживающие и размораживающие нейроны с более высокой плотностью 3 х 105 клеток на лунку.
- После 14-16 дня в кокультурах образование синапсов в iNs можно наблюдать путем окрашивания IHC пре- и постсинаптических маркеров, а к 21-му дню синаптическая пункта должна быть обильной (рисунок 3C), и нейронная активность может быть надежно зарегистрирована.
- Начиная с 21-го дня, протестируйте ячейки на наличие специфических маркеров OL (например, MBP и PLP1). К 28-му дню мы обычно наблюдаем явление оболивания аксонов iN процессами iOL, помеченное окрашиванием IHC для конкретных маркеров (рисунок 3B; нейрофиламент NF для аксонов iN и MBP для процессов iOPC).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Прямая генерация индуцированных человеком нейронов из человеческих плюрипотентных стволовых клеток
Очень важно, чтобы исходные плюрипотентные стволовые клетки человека проявляли высокую степень плюрипотентности для успешной генерации iNs или iOPCs / iOL. Поэтому клетки должны быть окрашены для конкретных маркеров, таких как Oct4 и SOX2, перед началом любого из протоколов индукции, описанных в настоящей рукописи (рисунок 1A). Человеческие клетки H1 использовались для получения индуцированных возбуждающих нейронов переднего мозга в соответствии с ранее опубликованным протоколом Zhang et al. с некоторыми модификациями (Рисунок 1C)12,16,17,18. Здесь мы представляем протокол, в котором iNs на 2-й день повторно покрываются в чистую культуру на матричном растворе (см. Таблицу материалов), при отсутствии какого-либо питающего слоя: глии или фибробластов. В дополнение к ранее опубликованным протоколам мы наблюдаем, что замораживание iNs на 2-й день существенно не влияет на жизнеспособность клеток (~ 15-20% гибели клеток после оттаивания). Чистые нейроны в культуре начнут экспрессировать синапсин1 на 14–16 день (рисунок 1D). Создание чистой нейронной культуры очень важно, потому что определенные факторы, например, ведущий фактор риска AD ApoE, могут быть экспрессированы клетками в фидерном слое, и это может значительно запутать результаты.
Генерация iOPC и созревание iOL улучшается лечением ДМСО
Здесь мы представляем быстрый и эффективный протокол, который позволяет генерировать iOPC за 2 недели и зрелые iOL за 4–5 недель (рисунок 2A). Мы использовали метод транзиторного лечения ДМСО, который мы ранее разработали, чтобы повысить эффективность дифференцировки для ES и iPS клеток 29,30,31. Лечение ДМСО обогащает количество клеток в ранней фазе G1 для лучшей интеграции сигналов, способствуя дифференцировке. Мы выполнили первое лечение перед индуцированием человеческих ES-клеток для генерации NPC, а второе лечение перед дифференцировкой NPC в iOPC. Мы можем обнаружить специфические маркеры OPC (Olig2, CSPG4, NKX2.2 и PDGFRA) уже через 2 недели после покрытия клеток ES H1 (рисунок 2B, E). Популяция iOPC на этом этапе довольно однородна, при этом >95% клеток положительны на окрашивание O4 и высокий уровень иммунореактивности для других маркеров (рисунок 2B). После начала созревания OL на 15-й день мы обычно можем обнаружить специфические маркеры OL (MBP, O1, CLDN11 и PLP1), начиная с 28-го дня (рисунок 2C, E). Экспрессия этих специфических для стадии маркеров коррелирует с течением развития олигодендроглиальных клеток и предполагает ускоренный темп, при этом маркеры NPC постепенно снижаются, маркеры OPC достигают пика примерно на второй неделе, а маркеры OL повышаются к третьей неделе (рисунок 2D, E)37. Обратите внимание, что этот процесс созревания диверсифицирует клеточные популяции. Субпопуляции в континууме, включающие несколько промежуточных стадий между OPC и зрелыми миелинизирующими олигодендроцитами, могут присутствовать и составлять различный процент от общего числа клеток, причем более зрелые клетки доминируют в более позднее время.
Для сравнения, мы приобрели iOPC с высокими ссылками и созрели в iOL в соответствии с протоколом производителя. Мы проверили экспрессию маркеров, упомянутых выше, как в наших препаратах iOPC и iOL, так и в клетках, которые мы приобрели. Мы определили, что клетки, сгенерированные в соответствии с нашим протоколом, имели более высокую экспрессию всех протестированных генов (рисунок 2E). Интересно, что когда мы проверили секретируемые уровни двух основных изоформ амилоид-β (Aβ40 и Aβ42) в iNs по сравнению с iOPC, мы заметили, что iOPC секретируют больше обоих фрагментов, но соотношение осталось прежним (рисунок 2F).
Совместное культивирование iNs и iOPC
Этот протокол оптимизирован специально для совместного культивирования iNs и iOPC и позволяет в режиме реального времени контролировать межклеточные коммуникации между этими двумя типами клеток в ходе развития нейронов. Идеальная плотность покрытия для обоих типов клеток должна быть определена с помощью серии титрования номеров клеток для достижения правильной дифференцировки (рисунок 3A). Ожидается, что через 4 недели в кокультурах iOPC будут адекватно дифференцированы на OL, которые являются положительными для конкретных маркеров, таких как MBP, и расширяют процессы до аксонов ensheath (рисунок 3B). Система кокультурирования может надежно увеличить количество синапсов, что указывает на то, что iOPC обеспечивают нейронную поддержку через физические контакты или высвобождение трофических факторов (рисунок 3C). Мы можем поддерживать кокультуры в приемлемом состоянии здоровья в течение 6 недель и наблюдать, что число синапсов и другие нейронные атрибуты плато около пятой недели. Следует отметить, что астроциты и микроглия отсутствуют в наших препаратах, и их отсутствие можно задокументировать, проверив экспрессию специфических маркеров (рисунок 3D). IOPCs экспрессируют большое количество хорошо охарактеризованных генов, которые потенциально могут реагировать и опосредуть зависимые от активности сигналы от соседних нейронов, в паракрине (например, нейротрофины и метаболиты) и / или синаптическом способе (рисунок 3E и 3F).

Рисунок 1: Прямая генерация индуцированных человеком нейронов (iNs) из hPSCs. (A) Блок-схема генерации iN. (B) Репрезентативные яркие полевые и иммунофлуоресцентные изображения исходной культуры плюрипотентных стволовых клеток человека (H1) для подтверждения плюрипотентности. Oct4 показан красным цветом, а Sox2 — зеленым. (C) Репрезентативные яркие полевые изображения iNs на 4-й и 6-й день. (D) Характерная морфология для дендритной арборизации и синапсиновой пункты в iNs, выращенных в чистой культуре в течение 24 дней и окрашенных иммунофлуоресцентным окрашиванием для дендритного маркера Map2 и пресинаптического маркера Synapsin 1 (Syn1). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.

Рисунок 2: Генерация iOPC и созревание iOL. (A) Блок-схема генерации iOPC и iOL. (B) Репрезентативные яркие полевые и иммунофлуоресцентные изображения iOPC на 15-й день. Olig2 (маркер пан-олигодендроглии) показан зеленым цветом, O4 (маркер OPC) красным, а DAPI синим. Визуализация показала, что >95% iOPC положительны для O4 и 25% для Olig2. (C) Репрезентативные изображения ярких полей и иммунофлуоресцентных изображений iOL на 28-й день. MBP отображается зеленым цветом, O1 — красным, а DAPI — синим. (D) Экспрессия NPC-маркера PAX6 резко снижается в iOPC на 14-й день и далее снижается до фона в OLs на 28-й день, что указывает на надежную трансдифференцировку NPC и высокий уровень однородности в популяции iOPC. (E) Профиль экспрессии во временном ходе общих маркерных генов OPC и OL в культурах, генерируемых описанным протоколом, без (-DMSO) или с (+DMSO) стадией инкубации DMSO (этапы 2.1.3 и 2.2.2), анализируемой в разные моменты времени. Для сравнения, коммерческие iOPC (см. Таблицу материалов) созревали в соответствии с инструкциями производителя, и оба iOPC (iOPC-Tempo) или iOL (iOL-Tempo) были протестированы на одни и те же маркеры. Как и ожидалось, MBP (зрелый маркер олигодендроцитов) не был обнаружен (N.D.) на ранних стадиях дифференцировки во всех тестируемых iOPC. DMSO значительно повысил эффективность дифференциации OPC и созревания OL. (F) Производство и секреция Aβ40 и Aβ42 в чистых культурах iNs и iOPCs, измеренные коммерческими наборами ИФА (см. Таблицу материалов) на супернатанте, полученном из чистых культур iNs и iOPCs как на 15-й день, так и нормализованные по номерам клеток (при плотности 200 000 клеток на скважину в 24-луночной пластине). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Данные в гистограммах строятся как среднее ± SEM (n ≥ 3). Статистическая значимость оценивалась t-тестом Стьюдента (*, p < 0,05; ***, p < 0,001); в (D), по сравнению с NPC; в (E), по сравнению с контрольным iOPC-Tempo; в (F), по сравнению с iN.

Рисунок 3: Совместная культура iNs и iOPC. (A) Репрезентативное яркое полевое изображение совместно культивируемых iNs и iOPC на 7-й день, показывающее надлежащую плотность для дальнейшего созревания. (B) Репрезентативное иммунофлуоресцентное изображение iNs и iOPCs, совместно культивируемых в течение 28 дней. Аксональный маркер нейрофиламента НФ показан зеленым цветом, а олигодендроцитарный маркер МБП — красным. Справа, сегмент аксона iN, заключенный процессом iOL (MBP+). (C) Образование синапсов, анализируемое в 4-недельных сокультурах. Клетки окрашивали для Synapsin 1 (Syn1, зеленый) и MAP2 (красный), а synaptic puncta количественно определяли конфокальным анализом плотности вдоль дендритных сегментов, как описано17,18. (D) В наших кокультурах iNs и iOPC (7 дней совместного культивирования) экспрессия маркеров астроцитов, ALDHL1 и GFAP, минимальна (сверху), а экспрессия маркеров микроглии, TMEM119, TREM2 и CD33, не обнаруживается (N.D.) qPCR. Таким образом, загрязнение этими двумя типами глиальных клеток исключается. (E) Кокультурирование iOPC с iN приводит к образованию нейрон-OPC синапсов. Флуоресцентный постсинаптический маркер PSD95-mCherry выражается только в OPC и отображает диффузный рисунок в отдельных культурах (слева), но агрегируется с образованием пункты в кокультурах (справа, обозначен стрелками; Tuj1, нейрональный маркер). (F) Экспрессия хорошо олигодендроглиальных генов, которые могут ощущать и реагировать на нейронную активность в чистых культурах iOPC на 14-й день. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Данные в гистограммах отображаются как среднее ± SEM (n ≥ 3). Статистическая значимость оценивалась по t-тесту Стьюдента (**, p < 0,005; ***, p < 0,001); в (C), по сравнению с условием отсутствия OPC; в (D), по сравнению с первичными астроцитами в верхней панели.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
В дополнение к физической и метаболической поддержке для стабилизации синапсных структур и облегчения проведения соляного сигнала путем миелинизации, клетки линии олигодендроцитов могут формировать паттерн активности нейронов посредством быстрых и динамических перекрестных разговоров с нейронами 5,6,7. В то время как при патологии БА олигодендроглиальные реакции первоначально рассматривались как просто вторичные по отношению к воспалению и окислительным стрессам, в настоящее время существуют многообещающие доказательства, утверждающие, что нарушение целостности миелина является ранним патогенным событием до появления агрегации Aβ и гиперфосфорилирования тау9. Кроме того, восстановление миелинизации путем самообновления OPC особенно уязвимо вAD 38, процесс, который в значительной степени зависит от активности нейронов. Таким образом, понимание механизма поддержки здоровой передачи сигналов нейронов-олигодендроцитов представляет собой прекрасную возможность для выявления новых терапевтических мишеней.
Протокол Ngn2 с одним транскрипционным фактором является одним из наиболее часто используемых методов генерации нейронов человека, полученных из стволовых клеток, и процедуры, описанные здесь, являются дальнейшими уточнениями для получения чистых нейронных культур. Наш протокол iOPC/iOL имеет период индукции короче, чем в ранее опубликованных исследованиях (от 4 до 24 недель), с надежным выходом и чистотой, сопоставимыми с другими широко используемыми протоколами 19,20,22,23,24,25,26,27,28 . Наш протокол вводит пошаговую дифференцировку ES-клеток к NPC, OPC и, наконец, олигодендроцитам путем характеризующихся паттернов и генерирует функциональные клетки, которые могут быть использованы для изучения регуляции гомеостаза миелинизации и восстановления in vitro или in vivo (например, путем приживления в модель мыши-дрожания), как описано в предыдущей работе. Улучшению нашего протокола в значительной степени способствует инкубация DMSO, которая активирует белок ретинобластомы и продлевает фазу G1 клеточного цикла, чтобы лучше интегрировать стимулы направленной дифференцировки, а также усиливает терминальную дифференцировку в функциональныепроизводные 29,30. Наконец, применение клемастина, мускаринового и антигистаминного соединения, выявленного с помощью скринингов лекарств для ремиелинизационнойтерапии 33, дополнительно сокращает созревание олигодендроцитов, как это наблюдалось у препарата iPS-клеток и живых животных 7,28.
Ограничения метода в основном заключаются во внутреннем несоответствии между упрощенными настройками in vitro и средой in vivo в мозге; это несоответствие приводит к дисконтированию полного потенциала развития на поздних стадиях для отдельных типов клеток мозга. Для iNs недавние исследования смогли поддерживать культуры в хорошем синаптическом здоровье в течение значительно длительного периода времени, но все же выявили некоторую относительную незрелость, проявляющуюся в виде сокращения спиноподобных структур и нарушения спонтанной синаптической передачи в «старых» культурах iN (даже в 25-месячных)39. В то время как iOPC были воспроизведены для миелинатных аксонов in vivo после трансплантации в трансгенный мозг мыши, анализы миелинизации in vitro с электронно-микроскопической оценкой по-прежнему представляют собой техническую проблему с неудовлетворительной эффективностью почти для всех опубликованных протоколов19,28, включая этот. Таким образом, наша система кокультуры нейрон-OPC не должна точно имитировать процесс старения мозга, а также позднюю стадию патологии БА. Скорее, он уникально способен распутать сложные межклеточные взаимодействия между нейронами и OPC или олигодендроцитами на ранней стадии, которые не зависят от миелинизации и в то же время имеют основополагающее значение для правильного развития нервной системы и патогенеза заболеваний.
Каждая из трех процедур, описанных здесь, имеет свои c ритические шаги в рамках протокола и может потребовать модификаций и устранения неполадок. Для протокола iN существует два критических этапа: выбор пуромицина (шаг 1.2.5) и плотность покрытия (шаг 1.2.6). Неполное удаление недостаточно трансдуцированных клеток приводит к загрязнению плохо дифференцированных клеток и ставит под угрозу выживание и функции нейронов. Необходимо будет рассмотреть модификации для более сильного отбора пуромицина с более высокой концентрацией и более длительной инкубацией, как описано на этапе 1.2.5. Подходящая плотность покрытия должна определяться титрованием для каждой плюрипотентной клеточной линии, так как низкая плотность приводит к коллапсу культур, а высокая плотность стимулирует агрегацию клеток и препятствует росту нейронов. Для протокола iOPC/iOL двумя важнейшими этапами являются контроль пролиферации клеток при дифференцировке OPC (этап 2.2.3) и плотность покрытия для созревания OL (этапы 2.2.4 и 2.3.2). Чрезмерный рост дифференцирующих NPC сигнализирует о плохой реакции на стимулы дифференцировки OPC и должен быть ослаблен соответствующей дозировкой лечения Ara-C (в пределах указанного диапазона). При покрытии OPC для созревания здесь предпочтительнее более низкий диапазон плотности клеток, поскольку разреженное распределение может облегчить индукцию физиологической морфологии сложных структур (как показано на рисунке 2C). Для протокола совместного культивирования iN-iOPC мы хотели бы обратить внимание на критический этап покрытия с соответствующей плотностью для обоих типов клеток (шаги 3.1.1 и 3.2.2). В частности, iNs могут плохо прикрепляться к поверхности между растущими OPC и иметь тенденцию отсоединяться первыми, когда культура достигает слияния. Оптимальное соотношение должно быть определено путем титрования номеров ячеек.
В целом, этот редукционистский подход, находящийся в наших протоколах, является мощным инструментом для препарирования специфических гетероклеточных взаимодействий от присущей человеческому мозгу сложности и служит для раскрытия олигодендроглиальной биологии в области здравоохранения и при БА. Таким образом, значение по отношению к существующим методам, на наш взгляд, довольно очевидно. Дополнительной полезностью методов, разработанных здесь для будущих применений, является клеточная терапия для демиелинизирующих состояний, таких как постлучеваятерапия 40 и травма спинного мозга41,42. Кроме того, высокая пропускная способность этой системы на основе стволовых клеток также может быть использована в более широком масштабе для скрининга библиотек малых молекул на наличие соединений, которые могут защищать или восстанавливать физиологический статус нейронов, OPC, олигодендроцитов и их взаимодействий. Таким образом, мы считаем, что протоколы, описанные здесь, облегчат будущую работу по разработке лучших инструментов моделирования и эффективных методов лечения БА и других нейродегенеративных расстройств.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Авторам нечего раскрывать.
Acknowledgments
Эта работа была поддержана грантами Национальных институтов здравоохранения (R00 AG054616 для Y.A.H. и T32 GM136566 для K.C.), Медицинской школы Стэнфордского университета и стипендии Siebel (присуждается S.C.). Y.A.H. является профессором трансляции GFL из Центра трансляционной неврологии в Институте трансляционных наук Брауна.
Materials
| Name | Company | Catalog Number | Comments |
| Accutase | STEMCELL Technologies | 7920 | |
| B27 supplement | ThermoFisher | 17504044 | |
| bFGF | ThermoFisher | PHG 0266 | |
| cAMP | MilliporeSigma | A9501 | |
| Clemastine | MilliporeSigma | SML0445 | |
| DMEM/F12 medium | STEMCELL Technologies | 36254 | |
| DMSO | ThermoFisher | D12345 | |
| Doxycycline | MilliporeSigma | D3072 | |
| Fetal Bovine Serum | ScienCell | 10 | |
| H1 human ES cells | WiCell | WA01 | |
| Matrigel | Corning | 354234 | |
| mTeSR plus | STEMCELL Technologies | 5825 | |
| N2 supplement | ThermoFisher | 17502001 | |
| Neurobasal A medium | ThermoFisher | 10888-022 | |
| Non Essential Amino Acids | ThermoFisher | 11140-050 | |
| PDGF-AA | R&D Systems | 221-AA-010 | |
| PEI | VWR | 71002-812 | |
| pMDLg/pRRE | Addgene | 12251 | |
| Polybrene | MilliporeSigma | TR-1003-G | |
| pRSV-REV | Addgene | 12253 | |
| Puromycin | ThermoFisher | A1113803 | |
| ROCK Inhibitor Y-27632 | STEMCELL Technologies | 72302 | |
| SAG | Tocris | 4366 | |
| STEMdiff Neural Progenitor Freezing Media | STEMCELL Technologies | 5838 | |
| STEMdiff SMADi Neural Induction Kit | STEMCELL Technologies | 8581 | |
| T3 triiodothyronine | MilliporeSigma | T6397 | |
| Tempo-iOlogo: Human iPSC-derived OPCs | Tempo BioScience | SKU102 | |
| TetO-Ng2-Puro | Addgene | 52047 | |
| VSV-G | Addgene | 12259 |
References
- Pelvig, D. P., Pakkenberg, H., Stark, A. K., Pakkenberg, B. Neocortical glial cell numbers in human brains. Neurobiology of Aging. 29 (11), 1754-1762 (2008).
- Barres, B. A. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 60 (3), 430-440 (2008).
- De Strooper, B., Karran, E. The cellular phase of Alzheimer's disease. Cell. 164 (4), 603-615 (2016).
- Monje, M. Myelin plasticity and nervous system function. Annual Review of Neuroscience. 41, 61-76 (2018).
- Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., Bergles, D. E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nature Neuroscience. 21 (5), 696-706 (2018).
- Gibson, E. M., et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science. 344 (6183), 1252304 (2014).
- Pan, S., Mayoral, S. R., Choi, H. S., Chan, J. R., Kheirbek, M. A. Preservation of a remote fear memory requires new myelin formation. Nature Neuroscience. 23 (4), 487-499 (2020).
- Thornton, M. A., Hughes, E. G. Neuron-oligodendroglia interactions: Activity-dependent regulation of cellular signaling. Neuroscience Letters. 727, 134916 (2020).
- Ettle, B., Schlachetzki, J. C. M., Winkler, J. Oligodendroglia and myelin in neurodegenerative diseases: more than just bystanders. Molecular Neurobiology. 53 (5), 3046-3062 (2016).
- Essayan-Perez, S., Zhou, B., Nabet, A. M., Wernig, M., Huang, Y. A. Modeling Alzheimer's disease with human iPS cells: advancements, lessons, and applications. Neurobiology of Disease. 130, 104503 (2019).
- Li, L., et al. GFAP mutations in astrocytes impair oligodendrocyte progenitor proliferation and myelination in an hiPSC model of Alexander disease. Cell Stem Cell. 23 (2), 239-251 (2018).
- Lin, Y. T., et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer's disease phenotypes in human iPSC-derived brain cell types. Neuron. 98 (6), 1294 (2018).
- TCW, J., et al. Cholesterol and matrisome pathways dysregulated in human APOE ε4 glia. bioRxiv. , (2019).
- Ang, C. E., Wernig, M.
- Penney, J., Ralvenius, W. T., Tsai, L. H. Modeling Alzheimer's disease with iPSC-derived brain cells. Molecular Psychiatry. 25 (1), 148-167 (2020).
- Zhang, Y., et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron. 78 (5), 785-798 (2013).
- Huang, Y. A., Zhou, B., Nabet, A. M., Wernig, M., Sudhof, T. C. Differential signaling mediated by ApoE2, ApoE3, and ApoE4 in human neurons parallels Alzheimer's Disease risk. Journal of Neuroscience. 39 (37), 7408-7427 (2019).
- Huang, Y. A., Zhou, B., Wernig, M., Sudhof, T. C. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Abeta Secretion. Cell. 168 (3), 427-441 (2017).
- Yang, N., et al. Generation of oligodendroglial cells by direct lineage conversion. Nature Biotechnology. 31 (5), 434-439 (2013).
- Douvaras, P., et al. Efficient generation of myelinating oligodendrocytes from primary progressive multiple sclerosis patients by induced pluripotent stem cells. Stem Cell Reports. 3 (2), 250-259 (2014).
- Lee, E. H., Park, C. H. Comparison of reprogramming methods for generation of induced-oligodendrocyte precursor cells. Biomolecules & Therapeutics (Seoul). 25 (4), 362-366 (2017).
- Ehrlich, M., et al. Rapid and efficient generation of oligodendrocytes from human induced pluripotent stem cells using transcription factors. Proceedings of the National Academy of Sciences of the United States of America. 114 (11), 2243-2252 (2017).
- Rodrigues, G. M. C., et al. Defined and scalable differentiation of human oligodendrocyte precursors from pluripotent stem cells in a 3D culture system. Stem Cell Reports. 8 (6), 1770-1783 (2017).
- Hu, B. Y., Du, Z. W., Li, X. J., Ayala, M., Zhang, S. C. Human oligodendrocytes from embryonic stem cells: conserved SHH signaling networks and divergent FGF effects. Development. 136 (9), 1443-1452 (2009).
- Izrael, M., et al. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Molecular and Cellular Neuroscience. 34 (3), 310-323 (2007).
- Yamashita, T., et al. Differentiation of oligodendrocyte progenitor cells from dissociated monolayer and feeder-free cultured pluripotent stem cells. PLoS One. 12 (2), 0171947 (2017).
- Wang, S., et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell. 12 (2), 252-264 (2013).
- Chanoumidou, K., Mozafari, S., Baron-Van Evercooren, A., Kuhlmann, T. Stem cell derived oligodendrocytes to study myelin diseases. Glia. 68 (4), 705-720 (2020).
- Chetty, S., et al. A simple tool to improve pluripotent stem cell differentiation. Nature Methods. 10 (6), 553-556 (2013).
- Li, J., et al. A transient DMSO treatment increases the differentiation potential of human pluripotent stem cells through the Rb family. PLoS One. 13 (12), 0208110 (2018).
- Sambo, D., Li, J., Brickler, T., Chetty, S. Transient treatment of human pluripotent stem cells with DMSO to promote differentiation. Journal of Visualized Experiments: JoVE. (149), (2019).
- Douvaras, P., Fossati, V. Generation and isolation of oligodendrocyte progenitor cells from human pluripotent stem cells. Nature Protocols. 10 (8), 1143-1154 (2015).
- Mei, F., et al. Micropillar arrays as a high-throughput screening platform for therapeutics in multiple sclerosis. Nature Medicine. 20 (8), 954-960 (2014).
- Madhavan, M., et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nature Methods. 15 (9), 700-706 (2018).
- Zhang, Y., et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 89 (1), 37-53 (2016).
- Grubman, A., et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer's disease reveals cell-type-specific gene expression regulation. Nature Neuroscience. 22 (12), 2087-2097 (2019).
- Goldman, S. A., Kuypers, N. J. How to make an oligodendrocyte. Development. 142 (23), 3983-3995 (2015).
- Behrendt, G., et al. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia. 61 (2), 273-286 (2013).
- Patzke, C., et al. Neuromodulator signaling bidirectionally controls vesicle numbers in human synapses. Cell. 179 (2), 498-513 (2019).
- Piao, J., et al. Human embryonic stem cell-derived oligodendrocyte progenitors remyelinate the brain and rescue behavioral deficits following radiation. Cell Stem Cell. 16 (2), 198-210 (2015).
- Keirstead, H. S., et al. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. Journal of Neuroscience. 25 (19), 4694-4705 (2005).
- Kim, D. S., et al. Rapid generation of OPC-like cells from human pluripotent stem cells for treating spinal cord injury. Experimental & Molecular Medicine. 49 (7), 361 (2017).
Tags
Неврология выпуск 165 индуцированные плюрипотентные стволовые клетки iPS-клетки эмбриональные стволовые клетки человека ES-клетки нейроны олигодендроциты клетки-предшественники олигодендроцитов OPCs болезнь Альцгеймера бета-амилоидные пептиды синапсыErratum
Formal Correction: Erratum: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions
Posted by JoVE Editors on 12/29/2020.
Citeable Link.
An erratum was issued for: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions. The Representative Results section has been updated.
Figure 3 was updated from:
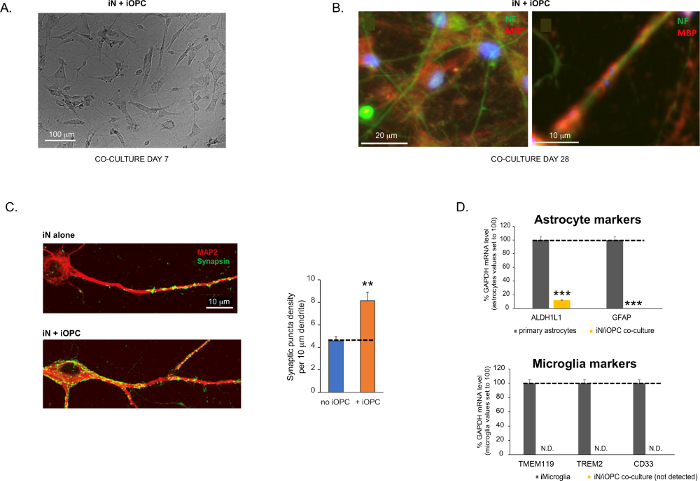
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. Please click here to view a larger version of this figure.
to:
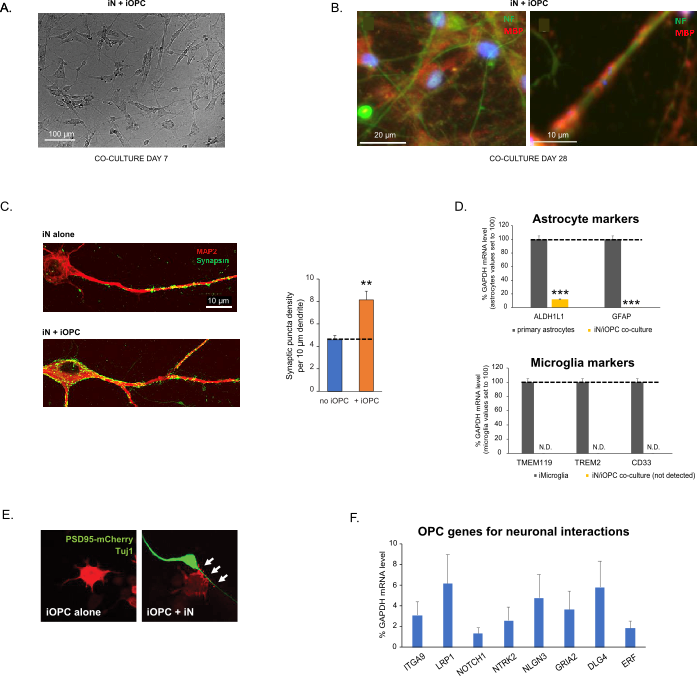
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. (E) Coculturing iOPC with iN leads to the formation of neuron-OPC synapses. The fluorescence-tagged post-synaptic marker PSD95-mCherry is expressed only in OPCs, and display a diffuse pattern in single cultures (left) but aggregate to form puncta in cocultures (right, indicated by arrows; Tuj1, neuronal marker). (F) The expression of well-characterized oligodendroglial genes that can sense and respond to neuronal activities in the pure cultures of iOPCs at Day 14. Please click here to view a larger version of this figure.
The fourth paragraph was updated from:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D).
to:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D). The iOPCs express a good number of well-characterized genes that can potentially respond to and mediate the activity-dependent signals from neighboring neurons, in a paracrine (e.g. neurotrophins and metabolites) and/or a synaptic manner (Figure 3E and 3F).
