

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Le interazioni neurone-glia nella neurodegenerazione non sono ben comprese a causa di strumenti e metodi inadeguati. Qui, descriviamo protocolli ottimizzati per ottenere neuroni indotti, cellule precursori degli oligodendrociti e oligodendrociti da cellule staminali pluripotenti umane e forniamo esempi dei valori di questi metodi nella comprensione dei contributi specifici del tipo di cellula nella malattia di Alzheimer.
Abstract
Nella malattia di Alzheimer (AD) e in altri disturbi neurodegenerativi, l'insufficienza oligodendrogliale è una caratteristica patologica precoce comune, ma il modo in cui contribuisce allo sviluppo e alla progressione della malattia, in particolare nella materia grigia del cervello, rimane in gran parte sconosciuto. La disfunzione delle cellule della linea oligodendrocitaria è caratterizzata da carenze nella mielinizzazione e compromissione dell'auto-rinnovamento delle cellule precursori degli oligodendrociti (OPC). Questi due difetti sono causati almeno in parte dall'interruzione delle interazioni tra neurone e oligodendrociti lungo l'accumulo di patologia. Gli OPC danno origine a oligodendrociti mielinizzanti durante lo sviluppo del SNC. Nella corteccia cerebrale matura, le OPC sono le principali cellule proliferative (che comprendono ~ 5% delle cellule cerebrali totali) e controllano la formazione di nuova mielina in modo dipendente dall'attività neurale. Tali comunicazioni neurone-oligodendrocita sono significativamente poco studiate, specialmente nel contesto di condizioni neurodegenerative come l'AD, a causa della mancanza di strumenti appropriati. Negli ultimi anni, il nostro gruppo e altri hanno compiuto progressi significativi per migliorare i protocolli attualmente disponibili per generare neuroni funzionali e oligodendrociti individualmente da cellule staminali pluripotenti umane. In questo manoscritto, descriviamo le nostre procedure ottimizzate, compresa la creazione di un sistema di co-coltura per modellare le connessioni neurone-oligodendrocitario. I nostri risultati illustrativi suggeriscono un contributo inaspettato da OPC / oligodendrociti all'amiloidosi cerebrale e all'integrità delle sinapsi ed evidenziano l'utilità di questa metodologia per la ricerca sull'AD. Questo approccio riduzionista è un potente strumento per sezionare le specifiche interazioni eterocellulari dalla complessità intrinseca all'interno del cervello. I protocolli che descriviamo qui dovrebbero facilitare studi futuri sui difetti oligodendrogliali nella patogenesi della neurodegenerazione.
Introduction
Le cellule della linea oligodendrocitaria, comprese le cellule precursori degli oligodendrociti (OPC), gli oligodendrociti mielinizzanti e i tipi di transizione intermedi, costituiscono un gruppo importante di cellule cerebrali umane1 che partecipano attivamente a molte funzioni critiche per il corretto funzionamento e mantenimento del nostro sistema nervoso centrale durante lo sviluppo neurale e l'invecchiamento 2,3,4 . Mentre gli oligodendrociti sono ben noti per la produzione di mielina per facilitare la trasmissione dell'attività neuronale e supportare la salute assonale nella sostanza bianca, le OPC sono abbondanti (~ 5%) nella materia grigia dove la mielinizzazione è scarsa e svolgono funzioni di segnalazione dipendenti dall'attività per governare il comportamento di apprendimento e la formazione della memoria 5,6,7,8 . Il modo in cui le cellule oligodendrogliali funzionano e disagiscono nella patogenesi della malattia di Alzheimer (AD) e di altre condizioni neurodegenerative associate all'età è stato poco studiato9. Le inadeguatezze di un sistema modello appropriato e le carenze nella conoscenza generale per guidare un percorso sperimentale in avanti sono le ragioni principali di questa lacuna.
Alla luce delle ultime scoperte nel derivare cellule cerebrali umane da cellule staminali pluripotenti, tra cui le cellule staminali embrionali (ES) e le cellule staminali pluripotenti indotte (iPS), tali modelli cellulari in combinazione con moderni strumenti di modifica genetica sono emersi come strumenti robusti per gestire l'intricato nesso delle interazioni cellulari nel cervello e sono in grado di dimostrare manifestazioni di malattia specifiche per l'uomo10, 11. Considerando che i singoli tipi di cellule cerebrali possono mostrare effetti distinti e persino contrastanti di fronte alle stesse condizioni che promuovono l'AD12,13, questa metodologia di cellule staminali offre in modo univoco informazioni specifiche sul tipo di cellula che in precedenza sono state perse utilizzando modelli stabiliti in vivo o in vitro che forniscono solo letture aggregate da collezioni di tipi di cellule cerebrali. Nell'ultimo decennio, un buon numero di protocolli affidabili sono stati sviluppati per generare neuroni umani dalla trans-differenziazione di cellule ES/iPS o dalla conversione diretta da altri tipi cellulari terminalmente differenziati (ad esempio, fibroblasti)14,15. In particolare, l'applicazione di fattori chiave di trascrizione neurogenica (ad esempio, neurogenina 2, Ngn2)16 a cellule staminali pluripotenti umane può generare una popolazione omogenea di tipi di cellule neuronali ben caratterizzati per colture pure senza necessità di cocoltura con cellule gliali12,17,18. Per gli oligodendrociti umani indotti, ci sono alcuni protocolli pubblicati che possono generare cellule funzionali molto simili alle loro controparti primarie, con una vasta gamma di efficienza e richiesta di tempo e risorse 19,20,21,22,23,24,25,26,27,28 . Ad oggi, nessuno di questi protocolli è stato applicato per studiare come le cellule oligodendrogliali rispondono e influenzano la patogenesi dell'AD.
Qui, descriviamo i nostri protocolli migliorati per colture singole e miste di neuroni indotti umani (iN) e OPC / oligodendrociti (iOPC / iOL). Il protocollo iN qui descritto si basa sull'approccio Ngn216 ampiamente utilizzato e ha la caratteristica aggiuntiva di essere privo di glia. Gli iN risultanti sono omogenei e molto simili ai neuroni eccitatori dello strato corticale 2/3, con morfologia piramidale caratteristica, pattern di espressione genica e caratteristiche elettrofisiologiche17,18 (Figura 1). Per superare alcune delle barriere fondamentali nella differenziazione diretta delle cellule staminali pluripotenti, abbiamo sviluppato un metodo semplice ed efficace di pre-trattamento a basse dosi di dimetilsolfossido (DMSO)29,30 e riportato una maggiore propensione delle cellule ES / iPS umane a transdifferenziarsi in iOPC e iOLs31, basato su un protocollo ampiamente adattato da Douvaras e Fossati 32 . Abbiamo ulteriormente semplificato il protocollo e incorporato un robusto composto che promuove la differenziazione, clemastina 7,33,34, per accelerare il processo di maturazione oligodendrogliale. Di conseguenza (Figura 2), le iOPC possono essere generate in 2 settimane (~95% positivo per il marcatore O4) e iOL in quattro settimane (esprimendo marcatori maturi MBP e PLP1). È interessante notare che abbiamo scoperto che le iOPC da sole secernono una notevole quantità di amiloide-β (Aβ), coerente con i dati trascrittomici indipendenti che mostrano l'abbondante espressione della proteina precursore dell'amiloide (APP) e della proteasi di trasformazione β-secretasi (BACE1) nelle cellule del lignaggio oligodendrocitario35,36. Inoltre, il nostro sistema di co-coltura iN-iOPC promuove l'inguaina degli assoni mediante processi iOL MBP-positivi e fornisce un supporto significativo per la formazione delle sinapsi (Figura 3). Pertanto, i protocolli che abbiamo dettagliato di seguito hanno vantaggi tecnici e biologici rispetto ai metodi di co-coltura neurone-oligodendroglia precedentemente catalogati e promettono di modellare meglio la neurodegenerazione nell'AD.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Induzione di neuroni umani da cellule staminali pluripotenti umane
- Preparazione del lentivirus (~5 giorni, protocollo dettagliato come descritto in precedenza16)
- Piastra ~ 1 milione di cellule HEK293T ogni pallone T75, per averle ~ 40% confluenti durante l'esecuzione della trasfezione. Trasfettarli con plasmidi che esprimono Ngn2 inducibile da tetraciclina e gene resistente alla puromicina (PuroR; sotto lo stesso controllo del promotore TetO), rtTA e i tre plasmidi helper pRSV-REV, pMDLg/pRRE e VSV-G (12 μg di DNA vettore lentivirale e 6 μg di ciascuno dei DNA plasmidi helper). Preparare almeno tre palloni per preparazione di lentivirus. Utilizzare PEI per la trasfezione seguendo le istruzioni del produttore. Cambiare il supporto dopo 16 ore e scartare.
- Harvest ha rilasciato particelle virali raccogliendo terreni di coltura ogni giorno e sostituirli con terreni freschi per 3 giorni. Raggruppare i mezzi raccolti contenenti particelle virali per la purificazione. Filtrare il virus attraverso un filtro da 0,22 μm e centrifugare a 49.000 x g per 90 minuti. Risospendere il pellet nel volume appropriato di glucosio PBS (~150 μL).
- Induzione neuronale (~5 giorni)
NOTA: Questo protocollo di induzione (Figura 1A; diagramma di flusso) è altamente efficace sia per le cellule iPS che ES di pluripotenza convalidata (che può essere dosata mediante colorazione immunoistochimica di marcatori di pluripotenza ben caratterizzati; Figura 1B).- Utilizzare cellule ES umane H1 disponibili in commercio al passaggio di 52 (vedi Tabella dei materiali). Coltura delle cellule su piastre a 6 pozzetti rivestite in soluzione di matrice extracellulare (~0,5 mg di soluzione di matrice per piastra a 6 pozzetti; vedere Tabella dei materiali) utilizzando il mezzo di manutenzione delle cellule ES (vedi Tabella dei materiali) e incubare le piastre a 37 °C con il 5% di CO2.
- Il giorno -2, staccare le cellule ES (80% confluenti) con 1 mL di soluzione di distacco cellulare (vedere Tabella dei materiali) e incubare a temperatura ambiente per 10 minuti. Trasferire le cellule in un tubo; lavare il pozzetto con 2 ml di fluido e combinare nello stesso tubo. Centrifugare a 300 x g per 5 minuti, risospendere il pellet in mezzi e placcare le celle su piastre a 6 pozzetti rivestite con matrice alla densità di semina di 1 x 105 celle per pozzetto.
- Il giorno -1, aggiungere lentivirus che esprimono Ngn2 più PuroR e rtTA insieme a polibrene (8 μg/ml) alle cellule ES in un mezzo di mantenimento delle cellule ES fresco (vedere Tabella dei materiali). La quantità esatta di virus deve essere determinata dai titoli effettivi o dalla titolazione. In genere aggiungiamo 5 μL per ogni virus per pozzetto in una piastra a 6 pozzetti.
- Il giorno 0, aggiungere Doxiciclina (2 μg/ml, per attivare l'espressione di Ngn2) nel mezzo DMEM-F12 con supplemento di N2 senza morfogeni.
- Il giorno 1, aggiungere Puromicina in mezzo fresco di DMEM-F12 più N2 e doxiciclina, alla concentrazione finale di 1 μg/mL di mezzo. Selezionare le cellule trasdotte in Puromicina per almeno 24 ore. Una maggiore concentrazione di puromicina (fino a 5 μg/ml) e un periodo di selezione più lungo (fino a 48 ore) possono essere necessari per rimuovere adeguatamente le cellule sotto-trasdotte se il titolo virale è basso.
- Il giorno 2, staccare i neuroni differenzianti con la soluzione di distacco cellulare (vedi Tabella dei materiali) e riplaccarli su piastre da 24 pozzetti (tra 80.000-200.000 cellule / pozzetto) rivestiti con soluzione di matrice (vedi Tabella dei materiali) e mantenerli in mezzo NBA / B27 senza doxiciclina. La densità di semina è fondamentale.
- In questa fase, i neuroni distaccati possono essere congelati in un mezzo di congelamento commerciale specializzato (vedi Tabella dei materiali) e conservati in azoto liquido per un massimo di 3 mesi. I neuroni puri possono essere placcati tenendo conto della tipica morte cellulare ~ 15% -20% post-disgelo, coltivati da soli o co-coltivati con altri tipi di cellule cerebrali (vedi passo 3.2.3. per la co-coltura con OPC).
- Coltura iNs pura sulle piastre rivestite con soluzioni a base di matrice extracellulare secondo le istruzioni del produttore (vedi Tabella dei materiali). La caratteristica morfologia piramidale dovrebbe essere evidente entro il Giorno 4 (e il Giorno 6; Figura 1C). La formazione di sinapsi può essere rilevata già dal giorno 14 al giorno 16 ed è prominente al giorno 24 mediante colorazione immunoistochimica con marcatori standard pre e post-sinaptici. (Figura 1D; etichettato con il marcatore pre-sinaptico Synapsin 1 e il marcatore dendritico Map2).
2. Induzione di cellule precursori degli oligodendrociti umani (OPC) da cellule staminali pluripotenti e maturazione degli oligodendrociti
- Generazione di cellule progenitrici neurali (NPC): protocollo monostrato (~7 giorni). Vedere la Figura 2A per il diagramma di flusso.
- Coltura di cellule ES umane H1 come descritto in precedenza (vedi passo 1.2.1.) e trans-differenziarle in cellule progenitrici neurali (NPC) con un approccio consolidato chiamato doppia SMADi, con inibitori di piccole molecole per molteplici vie di segnalazione. Qui utilizziamo un kit commerciale ampiamente accettato e seguiamo il protocollo monostrato fornito dal produttore (vedi Tabella dei materiali).
- Il giorno -1, piastra 0,5-1 x 10 6 celle per pozzetto in una piastra a 6 pozzetti rivestita da una soluzione a matrice ridotta con fattore di crescita (vedi Tabella dei materiali; ~ 0,5 mg di soluzione di matrice per piastra a6 pozzetti) con mezzo di mantenimento delle cellule ES (vedi Tabella dei materiali). Questa soluzione a matrice ridotta del fattore di crescita viene utilizzata per rivestire tutte le piastre che verranno utilizzate nelle fasi successive.
- Il giorno 0, trattare le cellule per 24 ore con terreno di mantenimento delle cellule ES (vedi Tabella dei materiali) integrato da DMSO al 2%.
- Nei giorni 1-6, sostituire il mezzo di induzione neurale completo con un mezzo di induzione neurale caldo (37 °C) contenente gli inibitori SMAD dal kit commerciale (vedere Tabella dei materiali). Se le cellule si dividono e raggiungono la confluenza prima del giorno 7, passarle alla densità di semina di 0,5-1 x 106, come descritto in precedenza nel passaggio 2.1.2.
- Il giorno 7, passaggio NPC utilizzando la soluzione di distacco cellulare (vedi Tabella dei materiali) e piastra ad una densità di semina di 1-2 x 105 celle / pozzetto di una piastra a 24 pozzetti.
- Analizzare l'efficienza di differenziazione mediante colorazione immunoistochimica (IHC) per l'assenza di marcatori di pluripotenza, ad esempio OCT4, e la presenza di marcatori NPC come PAX6, Nestin e Sox1.
- In questa fase, gli NPC distaccati possono essere congelati nei mezzi di congelamento NPC commerciali specializzati (vedi Tabella dei materiali) e conservati in azoto liquido per un massimo di 3 mesi. Dopo il congelamento e lo scongelamento per una volta, gli NPC mantengono ancora la multipotenza per dare origine a neuroni, astrociti e OPC con protocolli affidabili.
- Generazione di cellule precursori degli oligodendrociti (OPC) (~7 giorni). Vedere la Figura 2A per il diagramma di flusso.
- Il giorno 7, far passare gli NPC usando la soluzione di distacco cellulare (vedi Tabella dei materiali) e placcarli a una densità di semina di 1-2 x 105 cellule per pozzetto in una piastra a 24 pozzetti in mezzo di induzione neurale caldo (37 ° C) più inibitori SMAD dal kit commerciale (vedi Tabella dei materiali).
- Il giorno 8, preparare una soluzione di DMSO all'1% nel mezzo di differenziazione OPC e trattare gli NPC placcati per 24 ore. Il mezzo di differenziazione OPC è composto da: DMEM/F12 medio, 1% supplemento N2, 1% B27 supplemento, bFGF a 20 ng/mL, SAG a 1 μM, PDGF-AA a 10 ng/mL (vedi Tabella dei materiali).
- Il giorno 9, sostituire i supporti con un nuovo mezzo di differenziazione OPC senza DMSO. Nutrire le cellule a giorni alterni fino al giorno 15. Se le cellule raggiungono la confluenza prima del giorno 15, passarle alla densità di semina di 1-2 x 105 cellule per pozzetto come descritto al punto 2.2.1.
- Il giorno 14, OPC di piastre in mezzo di differenziazione OPC ad una densità di 1-2 x 105 celle / pozzetto in una piastra a 24 pozzetti.
- In questa fase (Giorno 15), celle di prova per la presenza di marcatori OPC-specifici mediante colorazione IHC o qPCR (ad esempio, O4, Olig1/2, CSPG4/Ng2, NKX2.2, PDGFRa; Figura 2B) e per l'assenza di marcatori NPC (Pax6 o Nestin; Figura 2D). In genere rileviamo l'immunoreattività O4 in oltre il 95% delle cellule al giorno 15. Di particolare rilevanza per la malattia di Alzheimer, l'espressione di APP (proteina precursore dell'amiloide), BACE1 (la proteasi di trasformazione β-secreatasi 1) e peptide amiloide-β (Aβ) è abbondante nelle OPC (Figura 2F).
- Maturazione degli oligodendrociti (OL) (~7–20 giorni)
- Il giorno 15, sostituire i terreni con il terreno di maturazione OL: terreno neurobasale-A, supplemento di B27 al 2%, cAMP da 1 μM, triiodotironina T3 da 200 ng/ml e Clemastina da 1 μM (vedere Tabella dei materiali). Cambia il mezzo a giorni alterni o ogni giorno, se necessario.
- Quando le cellule raggiungono il 90% di confluenza, dividersi in un rapporto 1: 3 fino a 2 passaggi o fino a quando la divisione cellulare rallenta sostanzialmente. Se gli OPC si dividono troppo velocemente e raggiungono la confluenza in meno di 3 giorni, aggiungere Ara-C (vedi Tabella dei materiali) ad una concentrazione di 2-5 μM per 1-3 giorni. La proliferazione attiva indica una ridotta efficienza di maturazione.
- Esaminare l'efficienza della maturazione oligodendrogliale valutando l'espressione di marcatori OL, ad esempio CLDN11, PLP1, MBP mediante qPCR, colorazione IHC o immunoblotting. La morfologia caratteristica di strutture altamente complesse (Figura 2C) e l'espressione dei marcatori OL (Figura 2E) dovrebbero essere prontamente rilevate entro il Giorno 28.
3. Co-coltura di neuroni umani indotti (iNs) e cellule precursori degli oligodendrociti (iOPCs)
- placcatura iOPC (~3 giorni)
- IOPC a piastra al giorno 14 ad una densità di 1 x 105 celle per pozzetto in una piastra a 24 pozzetti (come descritto sopra al punto 2.2.4.) nel mezzo di differenziazione OPC (come descritto nella fase 2.2.2.).
- Creazione della co-cultura iN-iOPC
- Il giorno 15, staccare i neuroni umani indotti nella fase del giorno 2 dopo la selezione della puromicina (come descritto nella fase 1.2.6.) con la soluzione di distacco cellulare (vedere Tabella dei materiali).
- Aggiungere neuroni agli OPC in coltura, placcando alla densità di semina di 2 x 105 cellule per pozzetto nella piastra a 24 pozzetti con OPC in crescita (dal punto 3.1.1). Utilizzare il terreno di coltura contenente terreno Neurobasal-A, supplemento di B27 al 2% e triiodotironina T3 da 100 ng / ml. Cambia il mezzo il giorno successivo e poi ogni altro giorno dopo. Se le OPC proliferano troppo velocemente e raggiungono la confluenza in meno di 3 giorni, aggiungere Ara-C ad una concentrazione di 2-5 μM. Un'immagine rappresentativa degli iNs e iOPC cresciuti in co-coltura dopo 7 giorni di neuroni è mostrata nella Figura 3A.
- Utilizzare neuroni congelati preparati come descritto sopra nel passaggio 1.2.7 per la co-coltura con OPC. Congelare e scongelare i neuroni delle piastre a una densità maggiore di 3 x 105 cellule per pozzetto.
- Dopo il giorno 14-16 nelle co-colture, la formazione di sinapsi nelle iNs può essere osservata dalla colorazione IHC dei marcatori pre- e post-sinaptici, ed entro il giorno 21 il puncta sinaptico dovrebbe essere abbondante (Figura 3C) e le attività neuronali possono essere registrate in modo affidabile.
- A partire dal giorno 21, celle di prova per marcatori specifici OL (ad esempio, MBP e PLP1). Dal giorno 28, osserviamo normalmente il fenomeno dell'inguaina degli assoni iN da parte dei processi iOL, etichettato mediante colorazione IHC per marcatori specifici (Figura 3B; neurofilamento NF per assoni iN e MBP per processi iOPC).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Generazione diretta di neuroni umani indotti da cellule staminali pluripotenti umane
È molto importante che le cellule staminali pluripotenti umane di partenza mostrino un alto grado di pluripotenza per la generazione di successo di iNs o iOPC / iOL. Pertanto, le cellule dovrebbero essere colorate per marcatori specifici, come Oct4 e SOX2, prima di iniziare uno dei protocolli di induzione descritti nel presente manoscritto (Figura 1A). Le cellule H1 umane sono state utilizzate per ottenere neuroni eccitatori del proencefalo indotti seguendo il protocollo precedentemente pubblicato da Zhang et al. con alcune modifiche (Figura 1C)12,16,17,18. Qui, presentiamo un protocollo in cui gli iN al Day 2 vengono riplaccati in coltura pura su soluzione di matrice (vedi Tabella dei materiali), in assenza di qualsiasi strato di alimentazione: glia o fibroblasti. Oltre ai protocolli pubblicati in precedenza, osserviamo che il congelamento di iNs al giorno 2 non influisce significativamente sulla vitalità cellulare (~ 15% -20% di morte cellulare dopo lo scongelamento). I neuroni puri in coltura inizieranno a esprimere sinapsina1 al giorno 14-16 (Figura 1D). Stabilire una coltura neuronale pura è molto importante perché alcuni fattori, ad esempio il principale fattore di rischio di AD ApoE, possono essere espressi dalle cellule nello strato di alimentazione e questo può confondere significativamente i risultati.
La generazione di iOPC e la maturazione di iOL sono migliorate dal trattamento DMSO
Qui presentiamo un protocollo veloce ed efficiente che consente la generazione di iOPC in 2 settimane e iOL maturi in 4-5 settimane (Figura 2A). Abbiamo sfruttato il metodo di trattamento con DMSO transitorio che abbiamo precedentemente sviluppato per aumentare l'efficienza di differenziazione per le cellule ES e iPS 29,30,31. Il trattamento DMSO arricchisce il numero di cellule nella fase iniziale G1 per una migliore integrazione della segnalazione, favorendo la differenziazione. Abbiamo eseguito il primo trattamento prima di indurre le cellule ES umane a generare NPC e il secondo trattamento prima di differenziare gli NPC in iOPC. Siamo in grado di rilevare specifici marcatori OPC (Olig2, CSPG4, NKX2.2 e PDGFRA) già 2 settimane dopo la placcatura delle cellule ES H1 (Figura 2B,E). La popolazione iOPC in questa fase è abbastanza omogenea, con il >95% delle cellule positive per la colorazione di O4 e un alto livello di immunoreattività per altri marcatori (Figura 2B). Dopo l'inizio della maturazione OL al Giorno 15, possiamo tipicamente rilevare specifici marcatori OL (MBP, O1, CLDN11 e PLP1) a partire dal Giorno 28 (Figura 2C,E). L'espressione di questi marcatori stadio-specifici è correlata con il decorso dello sviluppo delle cellule oligodendrogliali e suggerisce un ritmo accelerato, con i marcatori NPC che diminuiscono progressivamente, i marcatori OPC che raggiungono il picco intorno alla seconda settimana e i marcatori OL che aumentano entro la terza settimana (Figura 2D,E)37. Si noti che questo processo di maturazione diversifica le popolazioni cellulari. Le sottopopolazioni in continuo, che comprendono più stadi intermedi tra OPC e oligodendrociti mielinizzanti maturi, possono essere presenti e rappresentare una percentuale variabile delle cellule totali, con cellule più mature che dominano in un secondo momento.
A titolo di confronto, abbiamo acquistato gli iOPC altamente referenziati e li abbiamo maturati in iOL seguendo il protocollo del produttore. Abbiamo testato l'espressione dei marcatori sopra menzionati sia nei nostri preparati iOPC e iOL che nelle cellule che abbiamo acquistato. Abbiamo determinato che le cellule generate seguendo il nostro protocollo avevano una maggiore espressione di tutti i geni testati (Figura 2E). È interessante notare che, quando abbiamo testato i livelli secreti di due principali isoforme di β amiloide (Aβ40 e Aβ42) in iNs rispetto a iOPC, abbiamo notato che le iOPC secernevano più di entrambi i frammenti, ma il rapporto rimaneva lo stesso (Figura 2F).
Co-coltura di iNs e iOPC
Questo protocollo è ottimizzato specificamente per la co-coltura di iNs e iOPC e consente il nostro monitoraggio in tempo reale delle comunicazioni intercellulari tra questi due tipi di cellule lungo il corso dello sviluppo neurale. Le densità di placcatura ideali per entrambi i tipi di cellule devono essere decise con una serie di titolazioni del numero di cellule per ottenere una corretta differenziazione (Figura 3A). Dopo 4 settimane in co-colture, ci si aspetta che le iOPC siano adeguatamente differenziate in OL positivi per marcatori specifici come MBP ed estendano i processi agli assoni di enheath (Figura 3B). Il sistema di co-coltura può aumentare notevolmente il numero di sinapsi, indicando che le iOPC forniscono un supporto neuronale attraverso contatti fisici o rilascio di fattori trofici (Figura 3C). Possiamo mantenere le co-colture in condizioni di salute accettabili fino a 6 settimane e osservare che il numero di sinapsi e altri attributi neuronali si stabilizzano intorno alla quinta settimana. Da notare che astrociti e microglia non sono presenti nei nostri preparati e la loro assenza può essere documentata controllando l'espressione di marcatori specifici (Figura 3D). Le iOPC esprimono un buon numero di geni ben caratterizzati che possono potenzialmente rispondere e mediare i segnali dipendenti dall'attività dei neuroni vicini, in modo paracrino (ad esempio neurotrofine e metaboliti) e/o sinaptico (Figure 3E e 3F).

Figura 1: Generazione diretta di neuroni umani indotti (iN) da hPSC. (A) Diagramma di flusso della generazione di iN. (B) Immagini rappresentative in campo chiaro e immunofluorescenza della coltura iniziale di cellule staminali pluripotenti umane (H1) per confermare la pluripotenza. Oct4 è mostrato in rosso e Sox2 in verde. (C) Immagini rappresentative in campo chiaro di iNs al giorno 4 e al giorno 6. (D) La morfologia caratteristica per l'arborizzazione dendritica e la sinapsi puncta in iNs coltivate in coltura pura per 24 giorni e colorate mediante colorazione immunofluorescenza per marcatore dendritico Map2 e marcatore pre-sinaptico Synapsin 1 (Syn1). Fare clic qui per visualizzare una versione ingrandita di questa figura.

Figura 2: generazione di iOPC e maturazione di iOL. (A) Diagramma di flusso della generazione di iOPC e iOL. (B) Immagini rappresentative del campo luminoso e dell'immunofluorescenza delle iOPC al giorno 15. Olig2 (marcatore pan-oligodendroglia) è mostrato in verde, O4 (marcatore OPC) in rosso e DAPI in blu. L'imaging ha rivelato che >95% delle iOPC sono positive per O4 e il 25% per Olig2. (C) Immagini rappresentative del campo luminoso e dell'immunofluorescenza delle iOL al giorno 28. MBP è mostrato in verde, O1 in rosso e DAPI in blu. (D) L'espressione del marcatore NPC PAX6 diminuisce drasticamente nelle iOPC al giorno 14 e si abbassa ulteriormente allo sfondo negli OL al giorno 28, indicando una robusta trans-differenziazione NPC e un alto livello di omogeneità nella popolazione iOPC. (E) Il profilo di espressione del corso temporale di geni marcatori OPC e OL comuni in colture generate dal protocollo descritto, senza (-DMSO) o con (+DMSO) la fase di incubazione del DMSO (fasi 2.1.3 e 2.2.2), dosata in diversi punti temporali. A titolo di confronto, le iOPC commerciali (vedi Tabella dei materiali) sono state maturate secondo le istruzioni del produttore e sia le iOPC (iOPC-Tempo) che le iOL (iOL-Tempo) sono state testate per gli stessi marcatori. Come previsto, MBP (un marcatore di oligodendrociti maturi) non è stato rilevato (N.D.) nelle prime fasi di differenziazione in tutte le iOPC testate. Il DMSO ha migliorato significativamente l'efficienza della differenziazione OPC e della maturazione OL. (F) La produzione e la secrezione di Aβ40 e Aβ42 in colture di iNs e iOPC pure, misurate mediante kit ELISA commerciali (vedi Tabella dei materiali) su surnatante ottenuto da colture di iNs e iOPC pure sia al Giorno 15 che normalizzate dal numero di cellule (entrambe alla densità di 200.000 cellule per pozzetto in una piastra a 24 pozzetti). Fare clic qui per visualizzare una versione ingrandita di questa figura.
I dati nei grafici a barre sono rappresentati come media ± SEM (n ≥ 3). La significatività statistica è stata valutata mediante il t-test di Student (*, p < 0,05; ***, p < 0,001); in (D), rispetto all'NPC; in (E), rispetto al controllo iOPC-Tempo; in (F), rispetto a iN.

Figura 3: Co-coltura di iNs e iOPC. (A) Immagine rappresentativa in campo chiaro di iNs e iOPC co-coltivati al giorno 7, che mostra una densità adeguata per un'ulteriore maturazione. (B) Immagine rappresentativa di immunofluorescenza di iNs e iOPC co-coltivate per 28 giorni. Il neurofilamento marcatore assonale NF è mostrato in verde e il marcatore oligodendrocitico MBP in rosso. A destra, un segmento dell'assone iN inguainato dal processo iOL (MBP+). (C) Formazione di sinapsi analizzata in co-colture di 4 settimane. Le cellule sono state colorate per sinapsina 1 (Syn1, verde) e MAP2 (rosso), e la puncta sinaptica è stata quantificata mediante analisi confocale della densità lungo i segmenti dendritici come descritto17,18. (D) Nelle nostre co-colture di iNs e iOPC (7 giorni di co-coltura), l'espressione dei marcatori astrocitari, ALDHL1 e GFAP, è minima (in alto), e l'espressione dei marcatori della microglia, TMEM119, TREM2 e CD33, non viene rilevata (N.D.) dalla qPCR. La contaminazione da questi due tipi di cellule gliali è quindi esclusa. (E) La cocoltura di iOPC con iN porta alla formazione di sinapsi neurone-OPC. Il marcatore post-sinaptico PSD95-mCherry marcato con fluorescenza è espresso solo nelle OPC e mostra un modello diffuso in singole colture (a sinistra) ma aggregato per formare puncta nelle cocolture (a destra, indicato da frecce; Tuj1, marcatore neuronale). (F) L'espressione di geni oligodendrogliali ben caratterizzati che possono percepire e rispondere alle attività neuronali nelle colture pure di iOPC al Giorno 14. Fare clic qui per visualizzare una versione ingrandita di questa figura.
I dati nei grafici a barre sono tracciati come media ± SEM (n ≥ 3). La significatività statistica è stata valutata mediante il t-test di Student (**, p < 0,005; ***, p < 0,001); in (C), rispetto alla condizione senza OPC; in (D), rispetto agli astrociti primari nel pannello superiore.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Oltre al supporto fisico e metabolico per stabilizzare le strutture sinaptiche e facilitare la conduzione del segnale saltatorio da parte della mielinizzazione, le cellule del lignaggio oligodendrocitario possono modellare il modello di attività neuronale attraverso cross-talk rapidi e dinamici con i neuroni 5,6,7. Mentre nella patologia di AD le risposte oligodendrogliali erano inizialmente considerate semplicemente secondarie all'infiammazione e agli stress ossidativi, ora ci sono prove promettenti che sostengono che l'integrità della mielina compromessa è un evento patogeno precoce prima della comparsa dell'aggregazione Aβ e dell'iperfosforilazione tau9. Inoltre, la riparazione della mielinizzazione attraverso l'auto-rinnovamento delle OPC è particolarmente vulnerabile nell'AD38, un processo che dipende fortemente dalle attività neuronali. Comprendere il meccanismo per supportare la segnalazione di neuroni-oligodendrociti sani rappresenta quindi un'ottima opportunità per identificare nuovi bersagli terapeutici.
Il protocollo Ngn2 del singolo fattore di trascrizione è una delle tecniche più impiegate per la generazione di neuroni umani derivati da cellule staminali, e le procedure qui descritte sono ulteriori perfezionamenti per ottenere colture neuronali pure. Il nostro protocollo iOPC/iOL ha un periodo di induzione più breve rispetto agli studi pubblicati in precedenza (da 4 a 24 settimane), con una resa robusta e una purezza paragonabile ad altri protocolli comunemente usati 19,20,22,23,24,25,26,27,28 . Il nostro protocollo introduce la differenziazione graduale delle cellule ES in NPC, OPC e infine oligodendrociti mediante segnali di pattern caratterizzati e genera cellule funzionali che possono essere utilizzate per studiare la regolazione dell'omeostasi della mielinizzazione e la riparazione in vitro o in vivo (ad esempio, per attecchimento nel modello murino shiverer) come descritto nel lavoro precedente. Il miglioramento del nostro protocollo è fortemente promosso dall'incubazione del DMSO, che attiva la proteina retinoblastoma e prolunga la fase G1 del ciclo cellulare per integrare meglio gli stimoli di differenziamento diretto, e migliora anche il differenziamento terminale in derivati funzionali29,30. Infine, l'uso di clemastina, un composto muscarinico e antistaminico colpito identificato attraverso screening farmacologici per le terapie di rimielinizzazione33, riduce ulteriormente la maturazione degli oligodendrociti, come osservato nella preparazione di cellule iPS e animali vivi 7,28.
I limiti della tecnica risiedono principalmente nella discrepanza intrinseca tra le impostazioni semplificate in vitro e gli ambienti in vivo nel cervello; Questa discrepanza porta a uno sconto nel pieno potenziale di sviluppo nelle fasi avanzate per i singoli tipi di cellule cerebrali. Per gli iN, studi recenti sono stati in grado di mantenere le colture in buona salute sinaptica per un periodo di tempo considerevolmente lungo, ma hanno comunque rivelato una certa immaturità relativa manifestata come strutture simili alla colonna vertebrale ridotte e trasmissione sinaptica spontanea compromessa nelle "vecchie" colture di iN (anche quelle di 25 mesi)39. Mentre le iOPC sono state dimostrate in modo riproducibile agli assoni mielinati in vivo dopo essere state trapiantate in cervelli di topo transgenici, i saggi di mielinizzazione in vitro con valutazione al microscopio elettronico rappresentano ancora una sfida tecnica con efficienza insoddisfacente per quasi tutti i protocolli pubblicati19,28, incluso questo. Pertanto, non si prevede che il nostro sistema di co-coltura neurone-OPC imiti fedelmente il processo di invecchiamento cerebrale e la fase avanzata della patologia di AD. Piuttosto, è in grado di districare le elaborate interazioni intercellulari tra neuroni e OPC o oligodendrociti allo stadio iniziale, che sono indipendenti dalla mielinizzazione e tuttavia fondamentali per il corretto sviluppo neurale e la patogenesi della malattia.
Ciascuna delle tre procedure qui descritte ha le sue fasi critiche all'interno del protocollo e può richiedere modifiche e risoluzione dei problemi. Per il protocollo iN, ci sono due passaggi critici: la selezione della puromicina (fase 1.2.5) e la densità di placcatura (fase 1.2.6). La rimozione incompleta delle cellule sotto-trasdotte provoca la contaminazione di cellule scarsamente differenziate e compromette la sopravvivenza e le funzioni neuronali. Dovrebbero essere prese in considerazione le modifiche per una selezione più forte della puromicina con una maggiore concentrazione e un'incubazione più lunga, come descritto nella fase 1.2.5. La densità di placcatura adatta deve essere determinata mediante titolazione per ciascuna linea cellulare pluripotente, poiché la bassa densità porta al collasso delle colture e l'alta densità incoraggia l'aggregazione cellulare e impedisce la crescita neuronale. Per il protocollo iOPC/iOL, le due fasi critiche sono il controllo della proliferazione cellulare nel differenziamento OPC (fase 2.2.3) e la densità di placcatura per la maturazione OL (fasi 2.2.4 e 2.3.2). La crescita eccessiva di NPC differenzianti segnala una scarsa risposta agli stimoli di differenziazione OPC e deve essere smorzata da un dosaggio appropriato di trattamento con Ara-C (entro l'intervallo indicato). Durante la placcatura delle OPC per la maturazione, qui è preferibile un intervallo inferiore di densità cellulare poiché la distribuzione sparsa può facilitare l'induzione di una morfologia fisiologica di strutture complesse (come mostrato in Figura 2C). Per il protocollo di co-coltura iN-iOPC, vorremmo attirare l'attenzione sulla fase critica della placcatura con una densità appropriata per entrambi i tipi di cellule (fasi 3.1.1 e 3.2.2). In particolare, gli iNs potrebbero non attaccarsi bene alla superficie tra gli OPC in crescita e tendono a staccarsi per primi quando la coltura raggiunge la confluenza. Il rapporto ottimale dovrebbe essere deciso titolando i numeri delle celle.
Nel complesso, questo approccio riduzionista che risiede nei nostri protocolli è un potente strumento per sezionare le specifiche interazioni eterocellulari dalla complessità intrinseca del cervello umano e serve a scoprire la biologia oligodendrogliale nella salute e nell'AD. L'importanza rispetto ai metodi esistenti è quindi abbastanza evidente a nostro avviso. Un'ulteriore utilità dei metodi sviluppati qui per le applicazioni future è la terapia cellulare per condizioni demielinizzanti, come la post-radioterapia40 e la lesione del midollo spinale41,42. Inoltre, l'elevata capacità produttiva di questo sistema basato su cellule staminali può anche essere utilizzata su larga scala per selezionare librerie di piccole molecole alla ricerca di composti in grado di proteggere o ripristinare lo stato fisiologico di neuroni, OPC, oligodendrociti e le loro interazioni. Pertanto, riteniamo che i protocolli qui descritti faciliteranno il lavoro futuro nello sviluppo di migliori strumenti di modellazione e trattamenti efficaci per l'AD e altri disturbi neurodegenerativi.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Gli autori non hanno nulla da rivelare.
Acknowledgments
Questo lavoro è stato sostenuto dalle sovvenzioni del National Institutes of Health (R00 AG054616 a Y.A.H. e T32 GM136566 a K.C.), dalla Stanford University School of Medicine e da una Siebel Fellowship (assegnata a S.C.). Y.A.H. è un professore traslazionale GFL presso il Center for Translational Neuroscience del Brown Institute for Translational Sciences.
Materials
| Name | Company | Catalog Number | Comments |
| Accutase | STEMCELL Technologies | 7920 | |
| B27 supplement | ThermoFisher | 17504044 | |
| bFGF | ThermoFisher | PHG 0266 | |
| cAMP | MilliporeSigma | A9501 | |
| Clemastine | MilliporeSigma | SML0445 | |
| DMEM/F12 medium | STEMCELL Technologies | 36254 | |
| DMSO | ThermoFisher | D12345 | |
| Doxycycline | MilliporeSigma | D3072 | |
| Fetal Bovine Serum | ScienCell | 10 | |
| H1 human ES cells | WiCell | WA01 | |
| Matrigel | Corning | 354234 | |
| mTeSR plus | STEMCELL Technologies | 5825 | |
| N2 supplement | ThermoFisher | 17502001 | |
| Neurobasal A medium | ThermoFisher | 10888-022 | |
| Non Essential Amino Acids | ThermoFisher | 11140-050 | |
| PDGF-AA | R&D Systems | 221-AA-010 | |
| PEI | VWR | 71002-812 | |
| pMDLg/pRRE | Addgene | 12251 | |
| Polybrene | MilliporeSigma | TR-1003-G | |
| pRSV-REV | Addgene | 12253 | |
| Puromycin | ThermoFisher | A1113803 | |
| ROCK Inhibitor Y-27632 | STEMCELL Technologies | 72302 | |
| SAG | Tocris | 4366 | |
| STEMdiff Neural Progenitor Freezing Media | STEMCELL Technologies | 5838 | |
| STEMdiff SMADi Neural Induction Kit | STEMCELL Technologies | 8581 | |
| T3 triiodothyronine | MilliporeSigma | T6397 | |
| Tempo-iOlogo: Human iPSC-derived OPCs | Tempo BioScience | SKU102 | |
| TetO-Ng2-Puro | Addgene | 52047 | |
| VSV-G | Addgene | 12259 |
References
- Pelvig, D. P., Pakkenberg, H., Stark, A. K., Pakkenberg, B. Neocortical glial cell numbers in human brains. Neurobiology of Aging. 29 (11), 1754-1762 (2008).
- Barres, B. A. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 60 (3), 430-440 (2008).
- De Strooper, B., Karran, E. The cellular phase of Alzheimer's disease. Cell. 164 (4), 603-615 (2016).
- Monje, M. Myelin plasticity and nervous system function. Annual Review of Neuroscience. 41, 61-76 (2018).
- Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., Bergles, D. E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nature Neuroscience. 21 (5), 696-706 (2018).
- Gibson, E. M., et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science. 344 (6183), 1252304 (2014).
- Pan, S., Mayoral, S. R., Choi, H. S., Chan, J. R., Kheirbek, M. A. Preservation of a remote fear memory requires new myelin formation. Nature Neuroscience. 23 (4), 487-499 (2020).
- Thornton, M. A., Hughes, E. G. Neuron-oligodendroglia interactions: Activity-dependent regulation of cellular signaling. Neuroscience Letters. 727, 134916 (2020).
- Ettle, B., Schlachetzki, J. C. M., Winkler, J. Oligodendroglia and myelin in neurodegenerative diseases: more than just bystanders. Molecular Neurobiology. 53 (5), 3046-3062 (2016).
- Essayan-Perez, S., Zhou, B., Nabet, A. M., Wernig, M., Huang, Y. A. Modeling Alzheimer's disease with human iPS cells: advancements, lessons, and applications. Neurobiology of Disease. 130, 104503 (2019).
- Li, L., et al. GFAP mutations in astrocytes impair oligodendrocyte progenitor proliferation and myelination in an hiPSC model of Alexander disease. Cell Stem Cell. 23 (2), 239-251 (2018).
- Lin, Y. T., et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer's disease phenotypes in human iPSC-derived brain cell types. Neuron. 98 (6), 1294 (2018).
- TCW, J., et al. Cholesterol and matrisome pathways dysregulated in human APOE ε4 glia. bioRxiv. , (2019).
- Ang, C. E., Wernig, M.
- Penney, J., Ralvenius, W. T., Tsai, L. H. Modeling Alzheimer's disease with iPSC-derived brain cells. Molecular Psychiatry. 25 (1), 148-167 (2020).
- Zhang, Y., et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron. 78 (5), 785-798 (2013).
- Huang, Y. A., Zhou, B., Nabet, A. M., Wernig, M., Sudhof, T. C. Differential signaling mediated by ApoE2, ApoE3, and ApoE4 in human neurons parallels Alzheimer's Disease risk. Journal of Neuroscience. 39 (37), 7408-7427 (2019).
- Huang, Y. A., Zhou, B., Wernig, M., Sudhof, T. C. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Abeta Secretion. Cell. 168 (3), 427-441 (2017).
- Yang, N., et al. Generation of oligodendroglial cells by direct lineage conversion. Nature Biotechnology. 31 (5), 434-439 (2013).
- Douvaras, P., et al. Efficient generation of myelinating oligodendrocytes from primary progressive multiple sclerosis patients by induced pluripotent stem cells. Stem Cell Reports. 3 (2), 250-259 (2014).
- Lee, E. H., Park, C. H. Comparison of reprogramming methods for generation of induced-oligodendrocyte precursor cells. Biomolecules & Therapeutics (Seoul). 25 (4), 362-366 (2017).
- Ehrlich, M., et al. Rapid and efficient generation of oligodendrocytes from human induced pluripotent stem cells using transcription factors. Proceedings of the National Academy of Sciences of the United States of America. 114 (11), 2243-2252 (2017).
- Rodrigues, G. M. C., et al. Defined and scalable differentiation of human oligodendrocyte precursors from pluripotent stem cells in a 3D culture system. Stem Cell Reports. 8 (6), 1770-1783 (2017).
- Hu, B. Y., Du, Z. W., Li, X. J., Ayala, M., Zhang, S. C. Human oligodendrocytes from embryonic stem cells: conserved SHH signaling networks and divergent FGF effects. Development. 136 (9), 1443-1452 (2009).
- Izrael, M., et al. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Molecular and Cellular Neuroscience. 34 (3), 310-323 (2007).
- Yamashita, T., et al. Differentiation of oligodendrocyte progenitor cells from dissociated monolayer and feeder-free cultured pluripotent stem cells. PLoS One. 12 (2), 0171947 (2017).
- Wang, S., et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell. 12 (2), 252-264 (2013).
- Chanoumidou, K., Mozafari, S., Baron-Van Evercooren, A., Kuhlmann, T. Stem cell derived oligodendrocytes to study myelin diseases. Glia. 68 (4), 705-720 (2020).
- Chetty, S., et al. A simple tool to improve pluripotent stem cell differentiation. Nature Methods. 10 (6), 553-556 (2013).
- Li, J., et al. A transient DMSO treatment increases the differentiation potential of human pluripotent stem cells through the Rb family. PLoS One. 13 (12), 0208110 (2018).
- Sambo, D., Li, J., Brickler, T., Chetty, S. Transient treatment of human pluripotent stem cells with DMSO to promote differentiation. Journal of Visualized Experiments: JoVE. (149), (2019).
- Douvaras, P., Fossati, V. Generation and isolation of oligodendrocyte progenitor cells from human pluripotent stem cells. Nature Protocols. 10 (8), 1143-1154 (2015).
- Mei, F., et al. Micropillar arrays as a high-throughput screening platform for therapeutics in multiple sclerosis. Nature Medicine. 20 (8), 954-960 (2014).
- Madhavan, M., et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nature Methods. 15 (9), 700-706 (2018).
- Zhang, Y., et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 89 (1), 37-53 (2016).
- Grubman, A., et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer's disease reveals cell-type-specific gene expression regulation. Nature Neuroscience. 22 (12), 2087-2097 (2019).
- Goldman, S. A., Kuypers, N. J. How to make an oligodendrocyte. Development. 142 (23), 3983-3995 (2015).
- Behrendt, G., et al. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia. 61 (2), 273-286 (2013).
- Patzke, C., et al. Neuromodulator signaling bidirectionally controls vesicle numbers in human synapses. Cell. 179 (2), 498-513 (2019).
- Piao, J., et al. Human embryonic stem cell-derived oligodendrocyte progenitors remyelinate the brain and rescue behavioral deficits following radiation. Cell Stem Cell. 16 (2), 198-210 (2015).
- Keirstead, H. S., et al. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. Journal of Neuroscience. 25 (19), 4694-4705 (2005).
- Kim, D. S., et al. Rapid generation of OPC-like cells from human pluripotent stem cells for treating spinal cord injury. Experimental & Molecular Medicine. 49 (7), 361 (2017).
Tags
Neuroscienze Numero 165 cellule staminali pluripotenti indotte cellule iPS cellule staminali embrionali umane cellule ES neuroni oligodendrociti cellule progenitrici degli oligodendrociti OPC morbo di Alzheimer peptidi amiloide-beta sinapsiErratum
Formal Correction: Erratum: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions
Posted by JoVE Editors on 12/29/2020.
Citeable Link.
An erratum was issued for: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions. The Representative Results section has been updated.
Figure 3 was updated from:
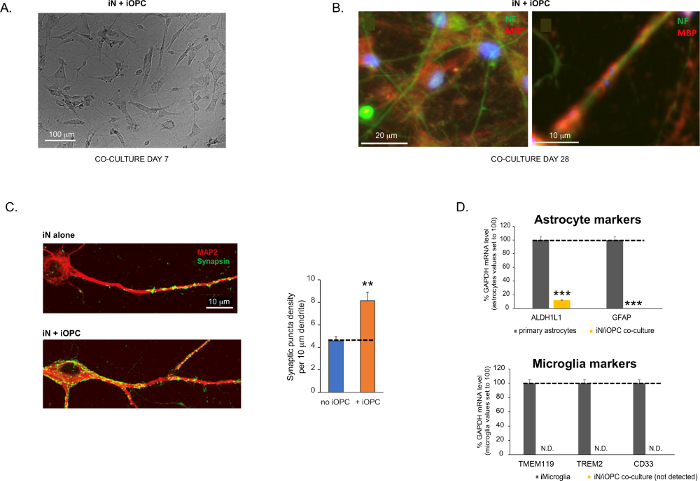
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. Please click here to view a larger version of this figure.
to:
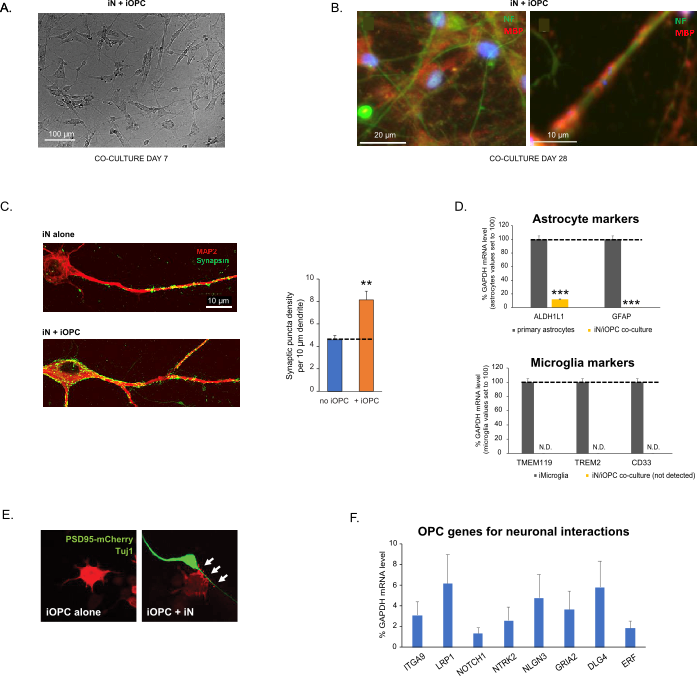
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. (E) Coculturing iOPC with iN leads to the formation of neuron-OPC synapses. The fluorescence-tagged post-synaptic marker PSD95-mCherry is expressed only in OPCs, and display a diffuse pattern in single cultures (left) but aggregate to form puncta in cocultures (right, indicated by arrows; Tuj1, neuronal marker). (F) The expression of well-characterized oligodendroglial genes that can sense and respond to neuronal activities in the pure cultures of iOPCs at Day 14. Please click here to view a larger version of this figure.
The fourth paragraph was updated from:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D).
to:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D). The iOPCs express a good number of well-characterized genes that can potentially respond to and mediate the activity-dependent signals from neighboring neurons, in a paracrine (e.g. neurotrophins and metabolites) and/or a synaptic manner (Figure 3E and 3F).
