

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Las interacciones neurona-glial en la neurodegeneración no se comprenden bien debido a herramientas y métodos inadecuados. Aquí, describimos protocolos optimizados para obtener neuronas inducidas, células precursoras de oligodendrocitos y oligodendrocitos a partir de células madre pluripotentes humanas y proporcionamos ejemplos de los valores de estos métodos para comprender las contribuciones específicas del tipo de célula en la enfermedad de Alzheimer.
Abstract
En la enfermedad de Alzheimer (EA) y otros trastornos neurodegenerativos, la insuficiencia oligodendroglial es una característica patológica temprana común, pero la forma en que contribuye al desarrollo y progresión de la enfermedad, particularmente en la materia gris del cerebro, sigue siendo en gran parte desconocida. La disfunción de las células del linaje de oligodendrocitos se caracteriza por deficiencias en la mielinización y alteración de la auto-renovación de las células precursoras de oligodendrocitos (OPC). Estos dos defectos son causados, al menos en parte, por la interrupción de las interacciones entre la neurona y los oligodendrocitos a lo largo de la acumulación de patología. Las OPC dan lugar a oligodendrocitos mielinizantes durante el desarrollo del SNC. En la corteza cerebral madura, las OPC son las principales células proliferativas (que comprenden ~ 5% del total de células cerebrales) y controlan la formación de nueva mielina de una manera dependiente de la actividad neuronal. Tales comunicaciones neuronómicas a oligodendrocitos están significativamente poco estudiadas, especialmente en el contexto de afecciones neurodegenerativas como la EA, debido a la falta de herramientas adecuadas. En los últimos años, nuestro grupo y otros han hecho progresos significativos para mejorar los protocolos actualmente disponibles para generar neuronas funcionales y oligodendrocitos individualmente a partir de células madre pluripotentes humanas. En este manuscrito, describimos nuestros procedimientos optimizados, incluido el establecimiento de un sistema de cocultivo para modelar las conexiones neurona-oligodendrocitos. Nuestros resultados ilustrativos sugieren una contribución inesperada de OPC / oligodendrocitos a la amiloidosis cerebral y la integridad de la sinapsis y destacan la utilidad de esta metodología para la investigación de la EA. Este enfoque reduccionista es una herramienta poderosa para diseccionar las interacciones heterocelulares específicas de la complejidad inherente dentro del cerebro. Se espera que los protocolos que describimos aquí faciliten futuros estudios sobre defectos oligodendrogliales en la patogénesis de la neurodegeneración.
Introduction
Las células del linaje de oligodendrocitos, incluidas las células precursoras de oligodendrocitos (OPC), los oligodendrocitos mielinizantes y los tipos de transición intermedios, constituyen un grupo importante de células cerebrales humanas1 que participan activamente en muchas funciones críticas para el correcto funcionamiento y mantenimiento de nuestro sistema nervioso central a lo largo del desarrollo neuronal y el envejecimiento 2,3,4 . Mientras que los oligodendrocitos son bien conocidos por producir mielina para facilitar la transmisión de la actividad neuronal y apoyar la salud axonal en la sustancia blanca, los OPC son abundantes (~ 5%) en la materia gris donde la mielinización es escasa y realizan funciones de señalización dependientes de la actividad para gobernar el comportamiento de aprendizaje y la formación de la memoria 5,6,7,8 . La forma en que las células oligodendrogliales funcionan y la disfunción en la patogénesis de la enfermedad de Alzheimer (EA) y otras afecciones neurodegenerativas asociadas a la edad han sido poco estudiadas9. Las insuficiencias de un sistema modelo apropiado y las deficiencias en el conocimiento general para guiar un camino experimental hacia adelante son las principales razones de esta brecha.
A la luz de los últimos avances en la obtención de células cerebrales humanas a partir de células madre pluripotentes, incluidas las células madre embrionarias (ES) y las células madre pluripotentes inducidas (iPS), tales modelos celulares junto con las herramientas modernas de edición de genes han surgido como herramientas robustas para manejar el intrincado nexo de las interacciones celulares en el cerebro, y son capaces de demostrar manifestaciones de enfermedades específicas del ser humano10, 11. Teniendo en cuenta que los tipos individuales de células cerebrales pueden exhibir efectos distintos e incluso conflictivos frente a las mismas condiciones promotoras de la EA12,13, esta metodología de células madre ofrece de manera única información específica del tipo de célula que anteriormente se había pasado por alto utilizando modelos establecidos in vivo o in vitro que solo proporcionan lecturas agregadas de colecciones de tipos de células cerebrales. En la última década, se han desarrollado un buen número de protocolos confiables para generar neuronas humanas a partir de la transdiferenciación de células ES/iPS o la conversión directa de otros tipos de células diferenciadas terminalmente (por ejemplo, fibroblastos)14,15. En particular, la aplicación de factores de transcripción neurogénicos clave (por ejemplo, neurogenina 2, Ngn2)16 a células madre pluripotentes humanas puede generar una población homogénea de tipos de células neuronales bien caracterizadas para cultivos puros sin necesidad de cocultivo con células gliales12,17,18. Para los oligodendrocitos humanos inducidos, hay algunos protocolos publicados que pueden generar células funcionales muy parecidas a sus contrapartes primarias, con una amplia gama de eficiencia y demanda en tiempo y recursos 19,20,21,22,23,24,25,26,27,28 . Hasta la fecha, ninguno de estos protocolos se ha aplicado para investigar cómo las células oligodendrogliales responden y afectan la patogénesis de la EA.
Aquí, describimos nuestros protocolos mejorados para cultivos únicos y mixtos de neuronas inducidas humanas (iNs) y OPC / oligodendrocitos (iOPC / iOL). El protocolo iN descrito aquí se basa en el enfoqueNgn2 16 ampliamente utilizado, y tiene la característica adicional de estar libre de glía. Los iNs resultantes son homogéneos y se asemejan mucho a las neuronas excitadoras de la capa cortical 2/3, con morfología piramidal característica, patrón de expresión génica y características electrofisiológicas17,18 (Figura 1). Para superar algunas de las barreras fundamentales en la diferenciación dirigida de células madre pluripotentes, hemos desarrollado un método simple y eficaz de pretratamiento con dosis bajas de dimetilsulfóxido (DMSO)29,30, y hemos reportado una mayor propensión de las células ES/iPS humanas a transdiferenciarse en iOPCs e iOLs31, basado en un protocolo ampliamente adaptado por Douvaras y Fossati 32 . Hemos simplificado aún más el protocolo e incorporado un compuesto robusto que promueve la diferenciación, clemastine 7,33,34, para acelerar el proceso de maduración oligodendroglial. Como resultado (Figura 2), las iOPC se pueden generar en 2 semanas (~95% positivo para el marcador O4) y las iOLs en cuatro semanas (expresando marcadores maduros MBP y PLP1). Curiosamente, encontramos que las iOPC solas secretan una cantidad notable de β amiloide (Aβ), consistente con los datos transcriptómicos independientes que muestran la abundante expresión de la proteína precursora amiloide (APP) y la proteasa de procesamiento β-secretasa (BACE1) en células de linaje oligodendrocitos35,36. Además, nuestro sistema de cocultivo iN-iOPC promueve el envoltorio de axones mediante procesos iOL positivos para MBP y proporciona un apoyo significativo para la formación de sinapsis (Figura 3). Por lo tanto, los protocolos que hemos detallado a continuación tienen ventajas técnicas y biológicas sobre los métodos de cocultivo neurona-oligodendroglia previamente catalogados, y prometen modelar mejor la neurodegeneración en la EA.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Inducción de neuronas humanas a partir de células madre pluripotentes humanas
- Preparación de lentivirus (~ 5 días, protocolo detallado como se describió anteriormente16)
- Placa ~1 millón de células HEK293T cada matraz T75, para tenerlas ~40% confluentes al realizar la transfección. Transfectarlos con plásmidos que expresan Ngn2 inducible por tetraciclina y gen resistente a la puromicina (PuroR; bajo el mismo control del promotor TetO), rtTA y los tres plásmidos auxiliares pRSV-REV, pMDLg/pRRE y VSV-G (12 μg de ADN vectorial lentiviral y 6 μg de cada uno de los ADN plásmidos auxiliares). Preparar al menos tres frascos por preparación de lentivirus. Use PEI para la transfección siguiendo las instrucciones del fabricante. Cambie el soporte después de 16 h y deséchelo.
- La cosecha liberó partículas virales recolectando medios de cultivo todos los días y reemplazándolos con medios nuevos durante 3 días. Agrupe los medios recolectados que contienen partículas virales para su purificación. Filtrar el virus a través de un filtro de 0,22 μm y centrifugar a 49.000 x g durante 90 min. Resuspender el pellet en el volumen apropiado de PBS-glucosa (~150 μL).
- Inducción neuronal (~5 días)
NOTA: Este protocolo de inducción (Figura 1A; diagrama de flujo) es altamente efectivo para células iPS y ES de pluripotencia validada (que puede ensayarse mediante tinción inmunohistoquímica de marcadores de pluripotencia bien caracterizados; Figura 1B).- Utilice células madre embrionarias humanas H1 disponibles comercialmente al paso de 52 (ver Tabla de materiales). Cultivar las células en placas de 6 pocillos recubiertas con solución de matriz extracelular (~0,5 mg de solución de matriz por placa de 6 pocillos; ver Tabla de materiales) utilizando medio de mantenimiento de células madre embrionarias (ver Tabla de materiales) e incubar las placas a 37 °C con 5% de CO2.
- En el día -2, separe las células madre embrionarias (80% confluentes) con 1 ml de solución de desprendimiento celular (consulte la Tabla de materiales) e incube a temperatura ambiente durante 10 minutos. Transfiera las células a un tubo; lavar el pozo con 2 mL de medio y combinar en el mismo tubo. Centrifugar a 300 x g durante 5 min, resuspender el pellet en medios y colocar las células en placas de 6 pocillos recubiertas con matriz a la densidad de siembra de 1 x 105 células por pocillo.
- El día -1, agregue lentivirus que expresen Ngn2 más PuroR y rtTA junto con polebreno (8 μg/ml) a las células madre embrionarias en un medio de mantenimiento de células madre embrionarias frescas (ver Tabla de materiales). La cantidad exacta de virus debe determinarse por títulos reales o la titulación. Por lo general, agregamos 5 μL cada virus por pocillo en una placa de 6 pocillos.
- El día 0, añadir doxiciclina (2 μg/ml, para activar la expresión de Ngn2) en medio DMEM-F12 con suplemento N2 sin morfógenos.
- El día 1, agregue puromicina en medio fresco de DMEM-F12 más N2 y doxiciclina, a la concentración final de 1 μg / ml de medio. Seleccione las células transducidas en Puromicina durante al menos 24 h. Puede ser necesaria una mayor concentración de puromicina (hasta 5 μg/ml) y un período de selección más largo (hasta 48 h) para eliminar adecuadamente las células subtransducidas si el título del virus es bajo.
- El día 2, separar las neuronas diferenciadoras con una solución de desprendimiento celular (ver Tabla de materiales) y volver a colocarlas en placas de 24 pocillos (entre 80,000-200,000 células / pocillo) recubiertas con solución de matriz (ver Tabla de materiales), y mantenerlas en medio NBA / B27 sin doxiciclina. La densidad de siembra es crítica.
- En esta etapa, las neuronas separadas pueden congelarse en un medio de congelación comercial especializado (consulte la Tabla de materiales) y almacenarse en nitrógeno líquido hasta por 3 meses. Las neuronas puras pueden ser plateadas que representan la típica muerte celular de ~15%-20% después del deshielo, cultivadas solas o cocultivadas con otros tipos de células cerebrales (ver paso 3.2.3. para el cocultivo con OPC).
- Cultive iNs puros en las placas recubiertas con soluciones basadas en matriz extracelular según las instrucciones del fabricante (consulte la Tabla de materiales). La morfología piramidal característica debe ser evidente en el día 4 (y el día 6; Figura 1C). La formación de sinapsis se puede detectar ya en el día 14 al 16 y es prominente en el día 24 mediante tinción inmunohistoquímica con marcadores pre y postsinápticos estándar. (Figura 1D; etiquetado con el marcador presináptico Synapsin 1 y el marcador dendrítico Map2).
2. Inducción de células precursoras de oligodendrocitos humanos (OPC) a partir de células madre pluripotentes y maduración de oligodendrocitos
- Generación de células progenitoras neurales (NPC): protocolo monocapa (~7 días). Consulte la Figura 2A para el diagrama de flujo.
- Cultivar células madre embrionarias humanas H1 como se describió anteriormente (ver paso 1.2.1.) y transdiferenciarlas en células progenitoras neurales (NPC) mediante un enfoque establecido llamado SMADi dual, con inhibidores de moléculas pequeñas para múltiples vías de señalización. Aquí utilizamos un kit comercial ampliamente aceptado y seguimos el protocolo monocapa proporcionado por el fabricante (ver Tabla de materiales).
- En el día -1, placa 0.5–1 x 10 6 células por pocillo en una placa de 6 pocillos recubierta por una solución de matriz reducida con factor de crecimiento (ver Tabla de materiales; ~0.5 mg de solución de matriz por placa de6 pocillos) con medio de mantenimiento de células madre embrionarias (ver Tabla de materiales). Esta solución de matriz reducida de factor de crecimiento se utiliza para recubrir todas las placas que se utilizarán en los siguientes pasos.
- El día 0, tratar las células durante 24 h con medio de mantenimiento de células madre embrionarias (ver Tabla de materiales) complementado con DMSO al 2%.
- En el día 1-6, cambie el medio completo con un medio de inducción neural tibio (37 °C) que contenga los inhibidores de SMAD del kit comercial (consulte la Tabla de materiales). Si las células se dividen y alcanzan la confluencia antes del día 7, pasarlas a la densidad de siembra de 0,5–1 x 106, como se describió anteriormente en el paso 2.1.2.
- El día 7, los NPC de paso usan una solución de desprendimiento celular (ver Tabla de materiales) y una placa a una densidad de siembra de 1–2 x 105 células/pocillo de una placa de 24 pocillos.
- Evaluar la eficiencia de diferenciación mediante tinción inmunohistoquímica (IHC) para detectar la ausencia de marcador de pluripotencia, OCT4, por ejemplo, y la presencia de marcadores NPC como PAX6, Nestin y Sox1.
- En esta etapa, los NPC separados pueden congelarse en los medios de congelación NPC comerciales especializados (consulte la Tabla de materiales) y almacenarse en nitrógeno líquido hasta por 3 meses. Después de congelar y descongelar por una vez, los NPC aún conservan la multipotencia para dar lugar a neuronas, astrocitos y OPC con protocolos confiables.
- Generación de células precursoras de oligodendrocitos (OPC) (~7 días). Consulte la Figura 2A para ver el diagrama de flujo.
- El día 7, pase NPC usando solución de desprendimiento celular (ver Tabla de materiales) y plaquelos a una densidad de siembra de 1–2 x 105 células por pocillo en una placa de 24 pocillos en medio de inducción neural caliente (37 °C) más inhibidores de SMAD del kit comercial (ver Tabla de materiales).
- El día 8, preparar una solución de DMSO al 1% en el medio de diferenciación OPC y tratar los NPC plateados durante 24 h. El medio de diferenciación OPC está compuesto por: medio DMEM/F12, suplemento de N2 al 1%, suplemento de B27 al 1%, bFGF a 20 ng/ml, SAG a 1 μM, PDGF-AA a 10 ng/ml (ver Tabla de materiales).
- El día 9, reemplace los medios con un nuevo medio de diferenciación OPC sin DMSO. Alimente las células cada dos días hasta el día 15. Si las células alcanzan la confluencia antes del día 15, pasarlas a la densidad de siembra de 1–2 x 105 células por pocillo, como se describe en el paso 2.2.1.
- El día 14, OPC en placa en medio de diferenciación OPC a una densidad de 1–2 x 105 células/pocillo en una placa de 24 pocillos.
- En esta etapa (Día 15), las células de prueba para detectar la presencia de marcadores específicos de OPC mediante tinción IHC o qPCR (p. ej., O4, Olig1/2, CSPG4/Ng2, NKX2.2, PDGFRa; Figura 2B) y por la ausencia de marcadores NPC (Pax6 o Nestin; Figura 2D). Por lo general, detectamos la inmunorreactividad O4 en más del 95% de las células en el día 15. De particular relevancia para la enfermedad de Alzheimer, la expresión de APP (proteína precursora amiloide), BACE1 (la proteasa de procesamiento β-secreatasa 1) y péptido amiloide-β (Aβ) es abundante en OPC (Figura 2F).
- Maduración de oligodendrocitos (OL) (~7–20 días)
- El día 15, reemplace los medios con medio de maduración OL: medio neurobasal-A, suplemento de B27 al 2%, cAMP de 1 μM, triyodotironina T3 de 200 ng / ml y clemastina de 1 μM (consulte la Tabla de materiales). Cambie el medio cada dos días o todos los días, si es necesario.
- Cuando las células alcanzan el 90% de confluencia, dividir en una proporción de 1:3 hasta 2 pasajes o hasta que la división celular disminuya sustancialmente. Si los OPC se dividen demasiado rápido y alcanzan la confluencia en menos de 3 días, agregue Ara-C (consulte la Tabla de materiales) a una concentración de 2-5 μM durante 1-3 días. La proliferación activa indica una menor eficiencia de maduración.
- Examinar la eficiencia de la maduración oligodendroglial evaluando la expresión de marcadores OL, por ejemplo, CLDN11, PLP1, MBP por qPCR, tinción IHC o immunoblotting. La morfología característica de estructuras altamente complejas (Figura 2C) y la expresión de marcadores OL (Figura 2E) deben detectarse fácilmente antes del día 28.
3. Cocultivo de neuronas humanas inducidas (iNs) y células precursoras de oligodendrocitos (iOPC)
- Recubrimiento iOPC (~3 días)
- IOPC en placa en el día 14 a una densidad de 1 x 105 células por pocillo en una placa de 24 pocillos (como se describe anteriormente en el paso 2.2.4.) en medio de diferenciación OPC (como se describe en el paso 2.2.2.).
- Creación de la cocultura iN-iOPC
- El día 15, separar las neuronas humanas inducidas en el paso del día 2 después de la selección de puromicina (como se describe en el paso 1.2.6.) con solución de desprendimiento celular (ver Tabla de materiales).
- Añadir neuronas a las OPC cultivadas, colocando en placa la densidad de siembra de 2 x 105 células por pocillo en la placa de 24 pocillos con OPC en crecimiento (a partir del paso 3.1.1). Use el medio de cocultivo que contiene medio neurobasal-A, suplemento de B27 al 2% y triyodotironina T3 de 100 ng / ml. Cambie el medio al día siguiente y luego cada dos días después. Si los OPC proliferan demasiado rápido y alcanzan la confluencia en menos de 3 días, agregue Ara-C a una concentración de 2-5 μM. Una imagen representativa de las iNs e iOPC cultivadas en cocultivo después de 7 días de neuronas se muestra en la Figura 3A.
- Utilice neuronas congeladas preparadas como se describe anteriormente en el paso 1.2.7 para el cocultivo con OPC. Las neuronas de congelación y descongelación de placas a una densidad mayor de 3 x 105 células por pocillo.
- Después del día 14-16 en cocultivos, la formación de sinapsis en iNs se puede observar mediante tinción IHQ de marcadores pre y postsinápticos, y para el día 21 los puntos sinápticos deben ser abundantes (Figura 3C) y las actividades neuronales se pueden registrar de manera confiable.
- A partir del día 21, pruebe las células para marcadores específicos de OL (por ejemplo, MBP y PLP1). Para el día 28, normalmente observamos el fenómeno de envainamiento de los axones iN mediante procesos iOL, marcados por tinción IHC para marcadores específicos (Figura 3B; neurofilamento NF para axones iN y MBP para procesos iOPC).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Generación directa de neuronas humanas inducidas a partir de células madre pluripotentes humanas
Es muy importante que las células madre pluripotentes humanas iniciales exhiban un alto grado de pluripotencia para la generación exitosa de iNs o iOPCs/iOLs. Por lo tanto, las células deben teñirse para marcadores específicos, como Oct4 y SOX2, antes de iniciar cualquiera de los protocolos de inducción descritos en el presente manuscrito (Figura 1A). Se utilizaron células H1 humanas para obtener neuronas excitatorias inducidas del prosencéfalo siguiendo el protocolo previamente publicado por Zhang et al. con algunas modificaciones (Figura 1C)12,16,17,18. Aquí, presentamos un protocolo en el que los iNs en el Día 2 se vuelven a colocar en cultivo puro en solución de matriz (ver Tabla de materiales), en ausencia de cualquier capa de alimentación: glía o fibroblastos. Además de los protocolos publicados anteriormente, observamos que la congelación de iNs en el día 2 no afecta significativamente la viabilidad celular (~ 15% -20% de muerte celular después de la descongelación). Las neuronas puras en cultivo comenzarán a expresar sinapsina1 en el día 14-16 (Figura 1D). Establecer un cultivo neuronal puro es muy importante porque ciertos factores, por ejemplo, el principal factor de riesgo de EA ApoE, pueden ser expresados por las células en la capa de alimentación y esto puede confundir significativamente los resultados.
La generación de iOPC y la maduración de iOL mejoran con el tratamiento con DMSO
Aquí, presentamos un protocolo rápido y eficiente que permite la generación de iOPC en 2 semanas y iOL maduros en 4-5 semanas (Figura 2A). Aprovechamos el método de tratamiento transitorio con DMSO que desarrollamos previamente para aumentar la eficiencia de diferenciación para células madre embrionarias e iPS 29,30,31. El tratamiento con DMSO enriquece el número de células en la fase temprana G1 para una mejor integración de la señalización, favoreciendo la diferenciación. Realizamos el primer tratamiento antes de inducir células madre embrionarias humanas para generar NPC, y el segundo tratamiento antes de diferenciar NPC en iOPC. Podemos detectar marcadores OPC específicos (Olig2, CSPG4, NKX2.2 y PDGFRA) tan pronto como 2 semanas después del recubrimiento de las células ES H1 (Figura 2B, E). La población de iOPC en esta etapa es bastante homogénea, con >95% de células positivas para tinción de O4 y un alto nivel de inmunorreactividad para otros marcadores (Figura 2B). Después del inicio de la maduración del OL en el día 15, normalmente podemos detectar marcadores OL específicos (MBP, O1, CLDN11 y PLP1) a partir del día 28 (Figura 2C, E). La expresión de estos marcadores específicos de la etapa se correlaciona con el curso del desarrollo de las células oligodendrogliales y sugiere un ritmo acelerado, con los marcadores NPC disminuyendo progresivamente, los marcadores OPC alcanzando su punto máximo alrededor de la segunda semana y los marcadores OL elevándose en la tercera semana (Figura 2D, E)37. Tenga en cuenta que este proceso de maduración diversifica las poblaciones celulares. Las subpoblaciones en el continuo, que comprenden múltiples etapas intermedias entre OPC y oligodendrocitos mielinizantes maduros, pueden estar presentes y representan un porcentaje variable de células totales, con células más maduras dominando en un momento posterior.
A modo de comparación, compramos los iOPC altamente referenciados y los maduramos en iOL siguiendo el protocolo del fabricante. Probamos la expresión de los marcadores mencionados anteriormente en nuestras preparaciones iOPC e iOL, y en las células que compramos. Determinamos que las células generadas siguiendo nuestro protocolo tenían mayor expresión de todos los genes probados (Figura 2E). Curiosamente, cuando probamos los niveles secretados de dos isoformas principales de β amiloide (Aβ40 y Aβ42) en iNs versus iOPC, notamos que las iOPC secretaban más de ambos fragmentos, pero la proporción seguía siendo la misma (Figura 2F).
Cocultivo de iNs e iOPCs
Este protocolo está optimizado específicamente para el cocultivo de iNs e iOPC y permite nuestro monitoreo en tiempo real de las comunicaciones intercelulares entre estos dos tipos de células a lo largo del desarrollo neuronal. Las densidades de recubrimiento ideales para ambos tipos de células deben decidirse con una serie de titulación del número de células para lograr una diferenciación adecuada (Figura 3A). Después de 4 semanas en cocultivos, se espera que las iOPC se diferencien adecuadamente en LO que son positivas para marcadores específicos como MBP y extienden los procesos a los axones de la vaina (Figura 3B). El sistema de cocultivo puede aumentar robustamente el número de sinapsis, lo que indica que las iOPC proporcionan un soporte neuronal a través de contactos físicos o liberación de factores tróficos (Figura 3C). Podemos mantener los cocultivos en condiciones de salud aceptables hasta por 6 semanas y observar que el número de sinapsis y otros atributos neuronales se estabilizan alrededor de la quinta semana. Cabe destacar que los astrocitos y la microglía no están presentes en nuestras preparaciones y su ausencia puede documentarse comprobando la expresión de marcadores específicos (Figura 3D). Las iOPC expresan un buen número de genes bien caracterizados que potencialmente pueden responder y mediar las señales dependientes de la actividad de las neuronas vecinas, de manera paracrina (por ejemplo, neurotrofinas y metabolitos) y / o sináptica (Figura 3E y 3F).

Figura 1: Generación directa de neuronas humanas inducidas (iNs) a partir de hPSCs. (A) Diagrama de flujo de la generación de iN. (B) Imágenes representativas de campo claro e inmunofluorescencia del cultivo inicial de células madre pluripotentes humanas (H1) para confirmar la pluripotencia. Oct4 se muestra en rojo y Sox2 en verde. (C) Imágenes representativas de campo brillante de iNs en el Día 4 y el Día 6. (D) La morfología característica para la arborización dendrítica y sinapsis puntual en iNs cultivados en cultivo puro durante 24 días y teñidos por tinción de inmunofluorescencia para el marcador dendrítico Map2 y el marcador presináptico Synapsin 1 (Syn1). Haga clic aquí para ver una versión más grande de esta figura.

Figura 2: generación de iOPC y maduración de iOL. (A) Diagrama de flujo de la generación de iOPC e iOL. (B) Imágenes representativas de campo brillante e inmunofluorescencia de iOPC en el día 15. Olig2 (marcador pan-oligodendroglia) se muestra en verde, O4 (marcador OPC) en rojo y DAPI en azul. Las imágenes revelaron que el >95% de las iOPC son positivas para O4 y el 25% para Olig2. (C) Imágenes representativas de campo brillante e inmunofluorescencia de iOLs en el día 28. MBP se muestra en verde, O1 en rojo y DAPI en azul. (D) La expresión del marcador NPC PAX6 disminuye drásticamente en las iOPC en el día 14 y disminuye aún más a fondo en las OL en el día 28, lo que indica una transdiferenciación robusta de NPC y un alto nivel de homogeneidad en la población de iOPC. (E) El perfil de expresión temporal de los genes marcadores OPC y OL comunes en cultivos generados por el protocolo descrito, sin (-DMSO) o con (+DMSO) la etapa de incubación de DMSO (pasos 2.1.3 y 2.2.2), ensayado en diferentes puntos temporales. A modo de comparación, las iOPC comerciales (ver Tabla de materiales) maduraron de acuerdo con las instrucciones del fabricante, y tanto las iOPC (iOPC-Tempo) como las iOL (iOL-Tempo) se probaron para los mismos marcadores. Como era de esperar, no se detectó MBP (un marcador de oligodendrocitos maduro) (ND) en las primeras etapas de diferenciación en todas las iOPC probadas. El DMSO mejoró significativamente la eficiencia de la diferenciación de OPC y la maduración de OL. (F) La producción y secreción de Aβ40 y Aβ42 en cultivos puros de iNs e iOPC, medida por kits comerciales de ELISA (ver Tabla de materiales) en sobrenadante obtenido de cultivos puros de iNs e iOPC tanto en el día 15 como normalizado por el número de células (ambos a la densidad de 200,000 células por pocillo en una placa de 24 pocillos). Haga clic aquí para ver una versión más grande de esta figura.
Los datos en los gráficos de barras se representan como media ± SEM (n ≥ 3). La significación estadística fue evaluada por la prueba t de Student (*, p < 0,05; ***, p < 0,001); en (D), en comparación con el NPC; en (E), en comparación con el control iOPC-Tempo; en (F), en comparación con iN.

Figura 3: Cocultivo de iNs e iOPCs. (A) Imagen representativa de campo brillante de iNs e iOPC cocultivados en el día 7, que muestra una densidad adecuada para una mayor maduración. (B) Imagen representativa de inmunofluorescencia de iNs e iOPCs cocultivados durante 28 días. El marcador axonal de neurofilamento NF se muestra en verde y el marcador oligodendrocítico MBP en rojo. Derecha, un segmento del axón iN enfundado por el proceso iOL (MBP+). (C) Formación de sinapsis ensayada en coculturas de 4 semanas de edad. Las células se tiñeron para Sinapsina 1 (Syn1, verde) y MAP2 (rojo), y los puntos sinápticos se cuantificaron mediante análisis confocal de la densidad a lo largo de los segmentos dendríticos como se describe17,18. (D) En nuestros cocultivos de iNs e iOPCs (7 días de co-cultivo), la expresión de los marcadores de astrocitos, ALDHL1 y GFAP, es mínima (arriba), y la expresión de los marcadores de microglía, TMEM119, TREM2 y CD33, no se detecta (N.D.) por qPCR. Por lo tanto, se excluye la contaminación de estos dos tipos de células gliales. (E) El cocultivo de iOPC con iN conduce a la formación de sinapsis neurona-OPC. El marcador postsináptico PSD95-mCherry marcado con fluorescencia se expresa solo en OPC y muestra un patrón difuso en cultivos individuales (izquierda) pero agregado para formar puntos en cocultivos (derecha, indicado por flechas; Tuj1, marcador neuronal). (F) La expresión de genes oligodendrogliales bien caracterizados que pueden detectar y responder a las actividades neuronales en los cultivos puros de iOPC en el día 14. Haga clic aquí para ver una versión más grande de esta figura.
Los datos en los gráficos de barras se representan como media ± SEM (n ≥ 3). La significación estadística fue evaluada por la prueba t de Student (**, p < 0,005; ***, p < 0,001); en (C), en comparación con la condición de ausencia de OPC; en (D), en comparación con los astrocitos primarios en el panel superior.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Además del apoyo físico y metabólico para estabilizar las estructuras sinapsis y facilitar la conducción de señales saltatorias por mielinización, las células del linaje de oligodendrocitos pueden dar forma al patrón de actividad neuronal a través de conversaciones cruzadas rápidas y dinámicas con las neuronas 5,6,7. Mientras que en la patología de la EA las respuestas oligodendrogliales se consideraron inicialmente como meramente secundarias a la inflamación y al estrés oxidativo, ahora hay evidencia prometedora que argumenta que la integridad comprometida de la mielina es un evento patogénico temprano antes de la aparición de la agregación de Aβ y la hiperfosforilación tau9. Además, la reparación de la mielinización a través de la autorrenovación de las OPC es particularmente vulnerable en elaño 38, un proceso que depende en gran medida de las actividades neuronales. Por lo tanto, comprender el mecanismo para apoyar la señalización neuronal-oligodendrocitos saludable representa una excelente oportunidad para identificar nuevos objetivos terapéuticos.
El protocolo del factor de transcripción único Ngn2 es una de las técnicas más empleadas para la generación de neuronas humanas derivadas de células madre, y los procedimientos descritos aquí son refinamientos adicionales para obtener cultivos neuronales puros. Nuestro protocolo iOPC/iOL tiene un período de inducción más corto que los estudios publicados anteriormente (4 a 24 semanas), con un rendimiento robusto y pureza comparable a otros protocolos de uso común 19,20,22,23,24,25,26,27,28 . Nuestro protocolo introduce la diferenciación gradual de células madre embrionarias a NPC, OPC y, finalmente, oligodendrocitos mediante señales de patrones caracterizadas, y genera células funcionales que pueden usarse para estudiar la regulación de la homeostasis y reparación de la mielinización in vitro o in vivo (por ejemplo, por injerto en el modelo de ratón tembloroso) como se describe en el trabajo anterior. La mejora en nuestro protocolo es promovida en gran medida por la incubación DMSO, que activa la proteína retinoblastoma y prolonga la fase G1 del ciclo celular para integrar mejor los estímulos de diferenciación dirigida, y también mejora la diferenciación terminal en derivados funcionales29,30. Finalmente, el uso de clemastina, un compuesto muscarínico y antihistamínico identificado a través de exámenes farmacológicos para la terapéutica de remielinización33, además acorta la maduración de oligodendrocitos, como se observa en la preparación de células iPS y animales vivos 7,28.
Las limitaciones de la técnica radican principalmente en la discrepancia intrínseca entre los entornos in vitro simplificados y los entornos in vivo en el cerebro; Esta discrepancia conduce a un descuento en el potencial de desarrollo total en las etapas avanzadas para los tipos de células cerebrales individuales. Para iNs, estudios recientes fueron capaces de mantener los cultivos en buena salud sináptica durante un período de tiempo considerablemente largo, pero aún revelaron cierta inmadurez relativa manifestada como estructuras similares a la columna vertebral reducidas y alteración de la transmisión sináptica espontánea en cultivos iN "viejos" (incluso los de 25 meses de edad)39. Si bien se ha demostrado de manera reproducible que las iOPC mielinizan axones in vivo después de ser trasplantadas en cerebros de ratones transgénicos, los ensayos de mielinización in vitro con evaluación microscópica electrónica aún representan un desafío técnico con eficiencia insatisfactoria para casi todos los protocolos publicados19,28, incluido este. Por lo tanto, no se prevé que nuestro sistema de cocultivo neurona-OPC imite fielmente el proceso de envejecimiento cerebral, así como la etapa tardía de la patología de la EA. Más bien, está en una posición única para desentrañar las elaboradas interacciones intercelulares entre las neuronas y las OPC o oligodendrocitos en etapa temprana, que son independientes de la mielinización y, sin embargo, fundamentales para el desarrollo neuronal adecuado y la patogénesis de la enfermedad.
Cada uno de los tres procedimientos descritos aquí tiene sus pasos básicos dentro del protocolo y puede requerir modificaciones y solución de problemas. Para el protocolo iN, hay dos pasos críticos: selección de puromicina (paso 1.2.5) y densidad de recubrimiento (paso 1.2.6). La eliminación incompleta de células subtransducidas resulta en la contaminación de células mal diferenciadas y compromete la supervivencia y las funciones neuronales. Habría que considerar las modificaciones para una selección más fuerte de puromicina con mayor concentración e incubación más prolongada, como se describe en el paso 1.2.5. La densidad de recubrimiento adecuada debe determinarse mediante titulación para cada línea celular pluripotente, ya que la baja densidad conduce al colapso de los cultivos y la alta densidad estimula la agregación celular e impide el crecimiento neuronal. Para el protocolo iOPC/iOL, los dos pasos críticos son el control de la proliferación celular en la diferenciación OPC (paso 2.2.3) y la densidad de recubrimiento para la maduración de OL (pasos 2.2.4 y 2.3.2). El crecimiento excesivo de NPC diferenciadores indica una respuesta deficiente a los estímulos de diferenciación de OPC y debe amortiguarse con una dosis adecuada de tratamiento con Ara-C (dentro del rango indicado). Al colocar OPC para la maduración, se prefiere un rango más bajo de densidad celular aquí, ya que la distribución dispersa puede facilitar la inducción de una morfología fisiológica de estructuras complejas (como se muestra en la Figura 2C). Para el protocolo de cocultivo iN-iOPC, nos gustaría llamar la atención sobre el paso crítico de recubrimiento con una densidad adecuada para ambos tipos de células (pasos 3.1.1 y 3.2.2). Específicamente, los iNs pueden no adherirse bien a la superficie entre los OPC en crecimiento y tienden a desprenderse primero cuando el cultivo alcanza la confluencia. La proporción óptima tendría que decidirse titulando el número de celdas.
En general, este enfoque reduccionista que reside en nuestros protocolos es una herramienta poderosa para diseccionar las interacciones heterocelulares específicas de la complejidad inherente del cerebro humano, y sirve para descubrir la biología oligodendroglial en la salud y en la EA. La importancia con respecto a los métodos existentes es, por lo tanto, bastante evidente en nuestra opinión. Una utilidad adicional de los métodos desarrollados aquí para las aplicaciones futuras es la terapia basada en células para afecciones desmielinizantes, como la post-radioterapia40 y la lesión de la médula espinal41,42. Además, la capacidad de alto rendimiento de este sistema basado en células madre también se puede utilizar a mayor escala para examinar bibliotecas de moléculas pequeñas en busca de compuestos que puedan proteger o restaurar el estado fisiológico de las neuronas, OPC, oligodendrocitos y sus interacciones. Por lo tanto, creemos que los protocolos descritos aquí facilitarán el trabajo futuro en el desarrollo de mejores herramientas de modelado y tratamientos efectivos para la EA y otros trastornos neurodegenerativos.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Los autores no tienen nada que revelar.
Acknowledgments
Este trabajo fue apoyado por las subvenciones de los Institutos Nacionales de Salud (R00 AG054616 a Y.A.H. y T32 GM136566 a K.C.), la Escuela de Medicina de la Universidad de Stanford y una beca Siebel (otorgada a SC). Y.A.H. es profesor traslacional de GFL del Centro de Neurociencia Traslacional del Instituto Brown de Ciencias Traslacionales.
Materials
| Name | Company | Catalog Number | Comments |
| Accutase | STEMCELL Technologies | 7920 | |
| B27 supplement | ThermoFisher | 17504044 | |
| bFGF | ThermoFisher | PHG 0266 | |
| cAMP | MilliporeSigma | A9501 | |
| Clemastine | MilliporeSigma | SML0445 | |
| DMEM/F12 medium | STEMCELL Technologies | 36254 | |
| DMSO | ThermoFisher | D12345 | |
| Doxycycline | MilliporeSigma | D3072 | |
| Fetal Bovine Serum | ScienCell | 10 | |
| H1 human ES cells | WiCell | WA01 | |
| Matrigel | Corning | 354234 | |
| mTeSR plus | STEMCELL Technologies | 5825 | |
| N2 supplement | ThermoFisher | 17502001 | |
| Neurobasal A medium | ThermoFisher | 10888-022 | |
| Non Essential Amino Acids | ThermoFisher | 11140-050 | |
| PDGF-AA | R&D Systems | 221-AA-010 | |
| PEI | VWR | 71002-812 | |
| pMDLg/pRRE | Addgene | 12251 | |
| Polybrene | MilliporeSigma | TR-1003-G | |
| pRSV-REV | Addgene | 12253 | |
| Puromycin | ThermoFisher | A1113803 | |
| ROCK Inhibitor Y-27632 | STEMCELL Technologies | 72302 | |
| SAG | Tocris | 4366 | |
| STEMdiff Neural Progenitor Freezing Media | STEMCELL Technologies | 5838 | |
| STEMdiff SMADi Neural Induction Kit | STEMCELL Technologies | 8581 | |
| T3 triiodothyronine | MilliporeSigma | T6397 | |
| Tempo-iOlogo: Human iPSC-derived OPCs | Tempo BioScience | SKU102 | |
| TetO-Ng2-Puro | Addgene | 52047 | |
| VSV-G | Addgene | 12259 |
References
- Pelvig, D. P., Pakkenberg, H., Stark, A. K., Pakkenberg, B. Neocortical glial cell numbers in human brains. Neurobiology of Aging. 29 (11), 1754-1762 (2008).
- Barres, B. A. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 60 (3), 430-440 (2008).
- De Strooper, B., Karran, E. The cellular phase of Alzheimer's disease. Cell. 164 (4), 603-615 (2016).
- Monje, M. Myelin plasticity and nervous system function. Annual Review of Neuroscience. 41, 61-76 (2018).
- Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., Bergles, D. E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nature Neuroscience. 21 (5), 696-706 (2018).
- Gibson, E. M., et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science. 344 (6183), 1252304 (2014).
- Pan, S., Mayoral, S. R., Choi, H. S., Chan, J. R., Kheirbek, M. A. Preservation of a remote fear memory requires new myelin formation. Nature Neuroscience. 23 (4), 487-499 (2020).
- Thornton, M. A., Hughes, E. G. Neuron-oligodendroglia interactions: Activity-dependent regulation of cellular signaling. Neuroscience Letters. 727, 134916 (2020).
- Ettle, B., Schlachetzki, J. C. M., Winkler, J. Oligodendroglia and myelin in neurodegenerative diseases: more than just bystanders. Molecular Neurobiology. 53 (5), 3046-3062 (2016).
- Essayan-Perez, S., Zhou, B., Nabet, A. M., Wernig, M., Huang, Y. A. Modeling Alzheimer's disease with human iPS cells: advancements, lessons, and applications. Neurobiology of Disease. 130, 104503 (2019).
- Li, L., et al. GFAP mutations in astrocytes impair oligodendrocyte progenitor proliferation and myelination in an hiPSC model of Alexander disease. Cell Stem Cell. 23 (2), 239-251 (2018).
- Lin, Y. T., et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer's disease phenotypes in human iPSC-derived brain cell types. Neuron. 98 (6), 1294 (2018).
- TCW, J., et al. Cholesterol and matrisome pathways dysregulated in human APOE ε4 glia. bioRxiv. , (2019).
- Ang, C. E., Wernig, M.
- Penney, J., Ralvenius, W. T., Tsai, L. H. Modeling Alzheimer's disease with iPSC-derived brain cells. Molecular Psychiatry. 25 (1), 148-167 (2020).
- Zhang, Y., et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron. 78 (5), 785-798 (2013).
- Huang, Y. A., Zhou, B., Nabet, A. M., Wernig, M., Sudhof, T. C. Differential signaling mediated by ApoE2, ApoE3, and ApoE4 in human neurons parallels Alzheimer's Disease risk. Journal of Neuroscience. 39 (37), 7408-7427 (2019).
- Huang, Y. A., Zhou, B., Wernig, M., Sudhof, T. C. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Abeta Secretion. Cell. 168 (3), 427-441 (2017).
- Yang, N., et al. Generation of oligodendroglial cells by direct lineage conversion. Nature Biotechnology. 31 (5), 434-439 (2013).
- Douvaras, P., et al. Efficient generation of myelinating oligodendrocytes from primary progressive multiple sclerosis patients by induced pluripotent stem cells. Stem Cell Reports. 3 (2), 250-259 (2014).
- Lee, E. H., Park, C. H. Comparison of reprogramming methods for generation of induced-oligodendrocyte precursor cells. Biomolecules & Therapeutics (Seoul). 25 (4), 362-366 (2017).
- Ehrlich, M., et al. Rapid and efficient generation of oligodendrocytes from human induced pluripotent stem cells using transcription factors. Proceedings of the National Academy of Sciences of the United States of America. 114 (11), 2243-2252 (2017).
- Rodrigues, G. M. C., et al. Defined and scalable differentiation of human oligodendrocyte precursors from pluripotent stem cells in a 3D culture system. Stem Cell Reports. 8 (6), 1770-1783 (2017).
- Hu, B. Y., Du, Z. W., Li, X. J., Ayala, M., Zhang, S. C. Human oligodendrocytes from embryonic stem cells: conserved SHH signaling networks and divergent FGF effects. Development. 136 (9), 1443-1452 (2009).
- Izrael, M., et al. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Molecular and Cellular Neuroscience. 34 (3), 310-323 (2007).
- Yamashita, T., et al. Differentiation of oligodendrocyte progenitor cells from dissociated monolayer and feeder-free cultured pluripotent stem cells. PLoS One. 12 (2), 0171947 (2017).
- Wang, S., et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell. 12 (2), 252-264 (2013).
- Chanoumidou, K., Mozafari, S., Baron-Van Evercooren, A., Kuhlmann, T. Stem cell derived oligodendrocytes to study myelin diseases. Glia. 68 (4), 705-720 (2020).
- Chetty, S., et al. A simple tool to improve pluripotent stem cell differentiation. Nature Methods. 10 (6), 553-556 (2013).
- Li, J., et al. A transient DMSO treatment increases the differentiation potential of human pluripotent stem cells through the Rb family. PLoS One. 13 (12), 0208110 (2018).
- Sambo, D., Li, J., Brickler, T., Chetty, S. Transient treatment of human pluripotent stem cells with DMSO to promote differentiation. Journal of Visualized Experiments: JoVE. (149), (2019).
- Douvaras, P., Fossati, V. Generation and isolation of oligodendrocyte progenitor cells from human pluripotent stem cells. Nature Protocols. 10 (8), 1143-1154 (2015).
- Mei, F., et al. Micropillar arrays as a high-throughput screening platform for therapeutics in multiple sclerosis. Nature Medicine. 20 (8), 954-960 (2014).
- Madhavan, M., et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nature Methods. 15 (9), 700-706 (2018).
- Zhang, Y., et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 89 (1), 37-53 (2016).
- Grubman, A., et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer's disease reveals cell-type-specific gene expression regulation. Nature Neuroscience. 22 (12), 2087-2097 (2019).
- Goldman, S. A., Kuypers, N. J. How to make an oligodendrocyte. Development. 142 (23), 3983-3995 (2015).
- Behrendt, G., et al. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia. 61 (2), 273-286 (2013).
- Patzke, C., et al. Neuromodulator signaling bidirectionally controls vesicle numbers in human synapses. Cell. 179 (2), 498-513 (2019).
- Piao, J., et al. Human embryonic stem cell-derived oligodendrocyte progenitors remyelinate the brain and rescue behavioral deficits following radiation. Cell Stem Cell. 16 (2), 198-210 (2015).
- Keirstead, H. S., et al. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. Journal of Neuroscience. 25 (19), 4694-4705 (2005).
- Kim, D. S., et al. Rapid generation of OPC-like cells from human pluripotent stem cells for treating spinal cord injury. Experimental & Molecular Medicine. 49 (7), 361 (2017).
Tags
Neurociencia Número 165 células madre pluripotentes inducidas células iPS células madre embrionarias humanas células ES neuronas oligodendrocitos células progenitoras de oligodendrocitos OPC enfermedad de Alzheimer péptidos beta-amiloide sinapsisErratum
Formal Correction: Erratum: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions
Posted by JoVE Editors on 12/29/2020.
Citeable Link.
An erratum was issued for: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions. The Representative Results section has been updated.
Figure 3 was updated from:
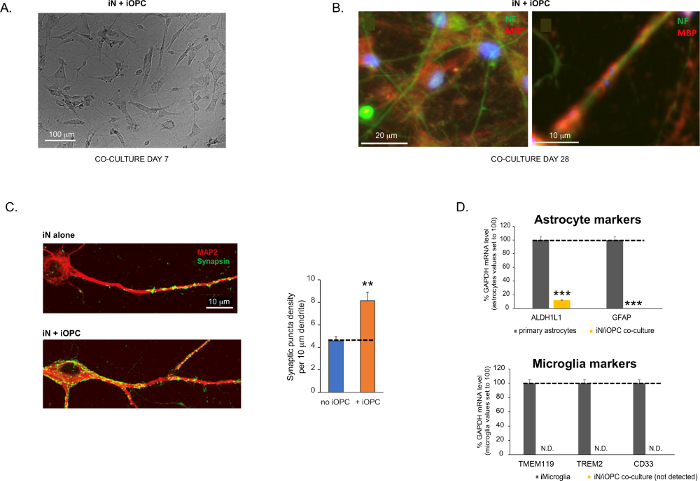
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. Please click here to view a larger version of this figure.
to:
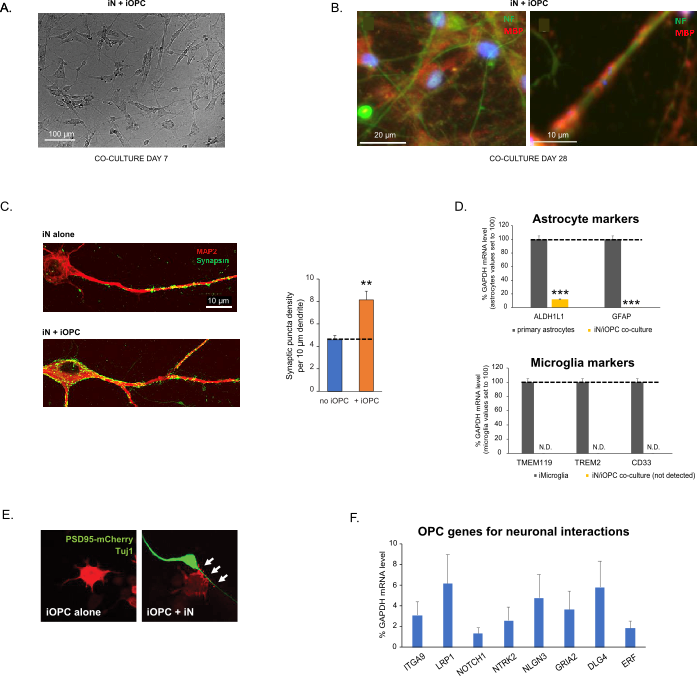
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. (E) Coculturing iOPC with iN leads to the formation of neuron-OPC synapses. The fluorescence-tagged post-synaptic marker PSD95-mCherry is expressed only in OPCs, and display a diffuse pattern in single cultures (left) but aggregate to form puncta in cocultures (right, indicated by arrows; Tuj1, neuronal marker). (F) The expression of well-characterized oligodendroglial genes that can sense and respond to neuronal activities in the pure cultures of iOPCs at Day 14. Please click here to view a larger version of this figure.
The fourth paragraph was updated from:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D).
to:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D). The iOPCs express a good number of well-characterized genes that can potentially respond to and mediate the activity-dependent signals from neighboring neurons, in a paracrine (e.g. neurotrophins and metabolites) and/or a synaptic manner (Figure 3E and 3F).
