

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
As interações neurônio-glial na neurodegeneração não são bem compreendidas devido a ferramentas e métodos inadequados. Aqui, descrevemos protocolos otimizados para obter neurônios induzidos, células precursoras de oligodendrócitos e oligodendrócitos de células-tronco pluripotentes humanas e fornecemos exemplos dos valores desses métodos na compreensão das contribuições específicas do tipo celular na doença de Alzheimer.
Abstract
Na doença de Alzheimer (DA) e outros distúrbios neurodegenerativos, a falha oligodendroglial é uma característica patológica precoce comum, mas como ela contribui para o desenvolvimento e progressão da doença, particularmente na massa cinzenta do cérebro, permanece amplamente desconhecida. A disfunção das células da linhagem de oligodendrócitos é marcada por deficiências na mielinização e auto-renovação prejudicada das células precursoras de oligodendrócitos (OPCs). Esses dois defeitos são causados, pelo menos em parte, pela interrupção das interações entre neurônios e oligodendrócitos ao longo do acúmulo de patologia. Os OPCs dão origem a oligodendrócitos mielinizantes durante o desenvolvimento do SNC. No córtex cerebral maduro, as OPCs são as principais células proliferativas (compreendendo ~ 5% do total de células cerebrais) e controlam a formação de nova mielina de maneira dependente da atividade neural. Tais comunicações neurônio-oligodendrócitos são significativamente pouco estudadas, especialmente no contexto de condições neurodegenerativas, como a DA, devido à falta de ferramentas apropriadas. Nos últimos anos, nosso grupo e outros fizeram progressos significativos para melhorar os protocolos atualmente disponíveis para gerar neurônios funcionais e oligodendrócitos individualmente a partir de células-tronco pluripotentes humanas. Neste manuscrito, descrevemos nossos procedimentos otimizados, incluindo o estabelecimento de um sistema de co-cultura para modelar as conexões neurônio-oligodendrócitos. Nossos resultados ilustrativos sugerem uma contribuição inesperada dos OPCs/oligodendrócitos para a amiloidose cerebral e a integridade das sinapses e destacam a utilidade dessa metodologia para a pesquisa da DA. Essa abordagem reducionista é uma ferramenta poderosa para dissecar as interações heterocelulares específicas da complexidade inerente dentro do cérebro. Espera-se que os protocolos que descrevemos aqui facilitem futuros estudos sobre defeitos oligodendrogliais na patogênese da neurodegeneração.
Introduction
As células da linhagem de oligodendrócitos – incluindo células precursoras de oligodendrócitos (OPCs), oligodendrócitos mielinizantes e tipos de transição entre eles – constituem um grupo importante de células cerebrais humanas1 que participam ativamente de muitas funções críticas para o bom funcionamento e manutenção do nosso sistema nervoso central ao longo do desenvolvimento neural e envelhecimento 2,3,4 . Enquanto os oligodendrócitos são bem conhecidos por produzir mielina para facilitar a transmissão da atividade neuronal e apoiar a saúde axonal na substância branca, as OPCs são abundantes (~5%) na massa cinzenta, onde a mielinização é escassa e desempenham funções de sinalização dependentes da atividade para governar o comportamento de aprendizagem e a formação da memória 5,6,7,8 . Como as células oligodendrogliais funcionam e a disfunção na patogênese da doença de Alzheimer (DA) e outras condições neurodegenerativas associadas à idade têm sido pouco estudadas9. As inadequações de um sistema modelo adequado e as deficiências nos conhecimentos gerais para orientar um caminho experimental a seguir são as principais razões para essa lacuna.
À luz dos mais recentes avanços na derivação de células cerebrais humanas a partir de células-tronco pluripotentes, incluindo células-tronco embrionárias (ES) e células-tronco pluripotentes induzidas (iPS), tais modelos celulares em conjunto com ferramentas modernas de edição de genes surgiram como ferramentas robustas para lidar com o intrincado nexo de interações celulares no cérebro e são capazes de demonstrar manifestações de doenças específicas de humanos10, 11. Considerando que os tipos individuais de células cerebrais podem exibir efeitos distintos e até conflitantes diante das mesmas condições promotoras de DA12,13, essa metodologia de células-tronco oferece exclusivamente informações específicas do tipo de célula que anteriormente foram perdidas usando modelos estabelecidos in vivo ou in vitro que fornecem apenas leituras agregadas de coleções de tipos de células cerebrais. Na última década, um bom número de protocolos confiáveis tem sido desenvolvido para gerar neurônios humanos a partir da transdiferenciação de células ES/iPS ou conversão direta de outros tipos celulares terminalmente diferenciados (por exemplo, fibroblastos)14,15. Em particular, a aplicação de fatores-chave de transcrição neurogênica (por exemplo, neurogenina 2, Ngn2)16 a células-tronco pluripotentes humanas pode gerar uma população homogênea de tipos de células neuronais bem caracterizadas para culturas puras sem a necessidade de cocultura com células gliais12,17,18. Para oligodendrócitos humanos induzidos, existem alguns protocolos publicados que podem gerar células funcionais altamente semelhantes às suas contrapartes primárias, com uma ampla gama de eficiência e demanda em tempo e recursos 19,20,21,22,23,24,25,26,27,28 . Até o momento, nenhum desses protocolos foi aplicado para investigar como as células oligodendrogliais respondem e afetam a patogênese da DA.
Aqui, descrevemos nossos protocolos aprimorados para culturas únicas e mistas de neurônios induzidos por humanos (iNs) e OPCs/oligodendrócitos (iOPCs/iOLs). O protocolo iN descrito aqui é baseado na abordagem Ngn2 amplamente utilizada16 e tem a característica adicional de ser livre de glia. Os iNs resultantes são homogêneos e se assemelham muito aos neurônios excitatórios da camada cortical 2/3, com morfologia piramidal, padrão de expressão gênica e características eletrofisiológicascaracterísticas 17,18 (Figura 1). Para superar algumas das barreiras fundamentais na diferenciação dirigida de células-tronco pluripotentes, desenvolvemos um método simples e eficaz de pré-tratamento com baixa dose de dimetilsulfóxido (DMSO)29,30 e relatamos uma maior propensão das células ES/iPS humanas a se transdiferenciarem em iOPCs e iOLs31, com base em um protocolo amplamente adaptado de Douvaras e Fossati 32 . Simplificamos ainda mais o protocolo e incorporamos um composto promotor de diferenciação robusto, a clemastina 7,33,34, para acelerar o processo de maturação oligodendroglial. Como resultado (Figura 2), os iOPCs podem ser gerados em 2 semanas (~95% positivos para o marcador O4) e os iOLs em quatro semanas (expressando marcadores maduros MBP e PLP1). Curiosamente, descobrimos que as iOPCs sozinhas secretam uma quantidade notável de β amiloide (Aβ), consistente com os dados transcriptômicos independentes que mostram a expressão abundante da proteína precursora amiloide (APP) e da protease de processamento β-secretase (BACE1) em células de linhagem de oligodendrócitos35,36. Além disso, nosso sistema de cocultura iN-iOPC promove o revestimento de axônios por processos iOL positivos para MBP e fornece suporte significativo para a formação de sinapses (Figura 3). Assim, os protocolos que detalhamos abaixo têm vantagens técnicas e biológicas sobre os métodos de co-cultura neurônio-oligodendroglia previamente catalogados e são promissores na melhor modelagem da neurodegeneração na DA.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Indução de neurônios humanos a partir de células-tronco pluripotentes humanas
- Preparação de Lentivirus (~5 dias, protocolo detalhado conforme descrito anteriormente16)
- Placa ~1 milhão de células HEK293T cada frasco T75, para tê-las ~40% confluentes ao realizar a transfecção. Transfecte-os com plasmídeos que expressam Ngn2 induzível por tetraciclina e gene resistente à puromicina (PuroR; sob o mesmo controle promotor de TetO), rtTA e os três plasmídeos auxiliares pRSV-REV, pMDLg/pRRE e VSV-G (12 μg de DNA vetorial lentiviral e 6 μg de cada um dos DNA plasmídeos auxiliares). Preparar pelo menos três frascos por preparação de lentivírus. Use PEI para transfecção seguindo as instruções do fabricante. Troque a mídia após 16 h e descarte o problema.
- Colha partículas virais liberadas coletando meios de cultura todos os dias e substitua por meios frescos por 3 dias. Agrupe o meio coletado contendo partículas virais para purificação. Filtrar o vírus através de um filtro de 0,22 μm e centrifugar a 49.000 x g durante 90 min. Ressuscite o pellet no volume apropriado de PBS-glicose (~150 μL).
- Indução de neurônios (~ 5 dias)
NOTA: Este protocolo de indução (Figura 1A; diagrama de fluxo) é altamente eficaz para células iPS e ES de pluripotência validada (que pode ser ensaiada por coloração imuno-histoquímica de marcadores de pluripotência bem caracterizados; Figura 1B).- Utilizar células ES humanas H1 comercialmente disponíveis à passagem de 52 (ver Tabela de Materiais). Cultivar as células em placas de 6 poços revestidas de solução de matriz extracelular (~0,5 mg de solução matricial por placa de 6 poços; ver Tabela de Materiais) utilizando meio de manutenção celular ES (ver Tabela de Materiais) e incubar as placas a 37 °C com 5% de CO2.
- No Dia -2, desprenda as células ES (80% confluentes) com 1 mL de solução de descolamento celular (ver Tabela de Materiais) e incube à temperatura ambiente por 10 min. Transfira as células para um tubo; lavar o poço com 2 mL de meio e combinar no mesmo tubo. Centrifugar a 300 x g durante 5 min, ressuspender o pellet em meio e chapear as células em placas de 6 poços revestidas de matriz à densidade de semeadura de 1 x 105 células por poço.
- No Dia -1, adicione lentivírus expressando Ngn2 mais PuroR e rtTA juntamente com polibreno (8 μg/ml) às células ES em meio de manutenção celular ES fresco (ver Tabela de Materiais). A quantidade exata de vírus deve ser determinada por títulos reais ou pela titulação. Normalmente, adicionamos 5 μL de cada vírus por poço em uma placa de 6 poços.
- No Dia 0, adicione doxiciclina (2 μg/mL, para ativar a expressão de Ngn2) em meio DMEM-F12 com suplemento de N2 sem morfogênios.
- No Dia 1, adicionar Puromicina em meio fresco de DMEM-F12 mais N2 e doxiciclina, à concentração final de 1 μg/mL de meio. Selecione as células transduzidas em Puromicina por pelo menos 24 h. Maior concentração de puromicina (até 5 μg/mL) e período de seleção mais longo (até 48 h) podem ser necessários para remover adequadamente as células sub-transduzidas se o título do vírus for baixo.
- No Dia 2, desprenda os neurônios diferenciadores com solução de descolamento celular (ver Tabela de Materiais) e recoloque-os em placas de 24 poços (entre 80.000 e 200.000 células/poço) revestidas com solução de matriz (ver Tabela de Materiais) e mantenha-as em meio NBA/B27 sem doxiciclina. A densidade de semeadura é crítica.
- Nesta fase, os neurônios destacados podem ser congelados em meio de congelamento comercial especializado (ver Tabela de Materiais) e armazenados em nitrogênio líquido por até 3 meses. Os neurônios puros podem ser banhados pela típica morte celular de ~15% a 20% após o descongelamento, cultivados sozinhos ou co-cultivados com outros tipos de células cerebrais (ver etapa 3.2.3. para co-cultura com OPCs).
- Cultura de iNs puro nas placas revestidas com soluções extracelulares à base de matriz, conforme instruído pelo fabricante (ver Tabela de Materiais). A morfologia piramidal característica deve ser aparente no Dia 4 (e Dia 6; Figura 1C). A formação de sinapses pode ser detectada já no dia 14 a 16 e é proeminente no dia 24 por coloração imuno-histoquímica com marcadores pré e pós-sinápticos padrão. (Figura 1D; rotulado com o marcador pré-sináptico Sinapsina 1 e o marcador dendrítico Map2).
2. Indução de células precursoras de oligodendrócitos humanos (OPCs) a partir de células-tronco pluripotentes e maturação de oligodendrócitos
- Geração de Células Progenitoras Neurais (NPC): protocolo monocamada (~7 dias). Consulte a Figura 2A para obter o diagrama de fluxo.
- Cultive células ES humanas H1 conforme descrito anteriormente (ver etapa 1.2.1.) e transdiferencie-as em células progenitoras neurais (NPCs) por uma abordagem estabelecida chamada SMADi duplo, com inibidores de moléculas pequenas para múltiplas vias de sinalização. Aqui usamos um kit comercial amplamente aceito e seguimos o protocolo monocamada fornecido pelo fabricante (consulte Tabela de Materiais).
- No Dia -1, placa 0,5–1 x 10 6 células por poço em uma placa de 6 poços revestida por uma solução de matriz reduzida com fator de crescimento (ver Tabela de Materiais; ~0,5 mg de solução matricial por placa de6 poços) com meio de manutenção de células ES (ver Tabela de Materiais). Esta solução de matriz reduzida com fator de crescimento é usada para revestir todas as placas que serão usadas nas etapas seguintes.
- No Dia 0, trate as células durante 24 horas com meio de manutenção celular ES (ver Tabela de Materiais) suplementado por 2% de DMSO.
- No Dia 1 a 6, troque o meio completo com o meio de indução neural quente (37 °C) contendo os inibidores de SMAD do kit comercial (ver Tabela de Materiais). Se as células se dividirem e atingirem a confluência antes do Dia 7, passe-as para a densidade de semeadura de 0,5–1 x 106, conforme descrito anteriormente na etapa 2.1.2.
- No Dia 7, os NPCs de passagem usando solução de descolamento celular (ver Tabela de Materiais) e placa a uma densidade de semeadura de 1–2 x 105 células/poço de uma placa de 24 poços.
- Avalie a eficiência de diferenciação por coloração imuno-histoquímica (IHC) para ausência de marcador de pluripotência, OCT4, por exemplo, e presença de marcadores NPC como PAX6, Nestin e Sox1.
- Nesta fase, os NPCs destacados podem ser congelados no meio de congelamento de NPC comercial especializado (consulte Tabela de Materiais) e armazenados em nitrogênio líquido por até 3 meses. Após o congelamento e descongelamento por uma vez, os NPCs ainda retêm a multipotência para dar origem a neurônios, astrócitos e OPCs com protocolos confiáveis.
- Geração de células precursoras de oligodendrócitos (OPC) (~7 dias). Consulte a Figura 2A para o diagrama de fluxo.
- No Dia 7, os NPCs de passagem usando solução de descolamento celular (ver Tabela de Materiais) e chapeá-los a uma densidade de semeadura de 1-2 x 105 células por poço em uma placa de 24 poços em meio de indução neural quente (37 °C) mais inibidores de SMAD do kit comercial (ver Tabela de Materiais).
- No dia 8, prepare uma solução de DMSO a 1% no meio de diferenciação OPC e trate os NPCs banhados por 24 h. O meio de diferenciação OPC é composto por: meio DMEM/F12, suplemento de N2 a 1%, suplemento de B27 a 1%, bFGF a 20 ng/mL, SAG a 1 μM, PDGF-AA a 10 ng/mL (ver Tabela de Materiais).
- No dia 9, substitua a mídia por um novo meio de diferenciação OPC sem DMSO. Alimente as células a cada dois dias até o dia 15. Se as células atingirem a confluência antes do Dia 15, passe-as para a densidade de semeadura de 1 a 2 x 105 células por poço, conforme descrito na etapa 2.2.1.
- No Dia 14, as OPCs de placa em meio de diferenciação de OPC a uma densidade de 1–2 x 105 células/poço em uma placa de 24 poços.
- Nesta fase (Dia 15), células de teste para a presença de marcadores específicos de OPC por coloração IHC ou qPCR (por exemplo, O4, Olig1/2, CSPG4/Ng2, NKX2.2, PDGFRa; Figura 2B) e para a ausência de marcadores NPC (Pax6 ou Nestin; Figura 2D). Normalmente, detectamos a imunorreatividade do O4 em mais de 95% das células no Dia 15. De particular relevância para a doença de Alzheimer, a expressão de APP (proteína precursora amiloide), BACE1 (protease de processamento β-secreatase 1) e peptídeo amilóide-β (Aβ) é abundante em OPCs (Figura 2F).
- Maturação de oligodendrócitos (OL) (~7–20 dias)
- No Dia 15, substitua a mídia pelo meio de maturação OL: meio neurobasal-A, suplemento de B27 a 2%, 1 μM de cAMP, triiodotironina T3 de 200 ng/mL e clemastina de 1 μM (ver Tabela de Materiais). Mude o meio a cada dois dias ou todos os dias, se necessário.
- Quando as células atingirem 90% de confluência, dividam-se em uma proporção de 1:3 até 2 passagens ou até que a divisão celular diminua substancialmente. Se os OPCs se dividirem muito rápido e atingirem a confluência em menos de 3 dias, adicione Ara-C (ver Tabela de Materiais) a uma concentração de 2–5 μM por 1–3 dias. A proliferação ativa indica menor eficiência de maturação.
- Examine a eficiência da maturação oligodendroglial avaliando a expressão de marcadores OL, por exemplo, CLDN11, PLP1, MBP por qPCR, coloração IHC ou immunoblotting. A morfologia característica de estruturas de alta complexidade (Figura 2C) e a expressão de marcadores OL (Figura 2E) devem ser prontamente detectadas até o Dia 28.
3. Co-cultura de neurônios induzidos pelo homem (iNs) e células precursoras de oligodendrócitos (iOPCs)
- Revestimento iOPC (~3 dias)
- IOPCs de placa no dia 14 a uma densidade de 1 x 105 células por poço em uma placa de 24 poços (conforme descrito acima na etapa 2.2.4.) em meio de diferenciação OPC (conforme descrito na etapa 2.2.2.).
- Configuração da co-cultura iN-iOPC
- No Dia 15, desprenda os neurónios humanos induzidos no passo do Dia 2 após a seleção da Puromicina (conforme descrito no passo 1.2.6.) com a solução de descolamento celular (ver Tabela de Materiais).
- Adicionar neurônios aos OPCs cultivados, chapeando na densidade de semeadura de 2 x 105 células por poço na placa de 24 poços com OPCs em crescimento (a partir da etapa 3.1.1). Use o meio de co-cultura contendo meio Neurobasal-A, suplemento de B27 a 2% e triiodotironina T3 a 100 ng/mL. Mude o meio no dia seguinte e, em seguida, a cada dois dias depois. Se as OPCs proliferarem muito rápido e atingirem a confluência em menos de 3 dias, adicione Ara-C a uma concentração de 2–5 μM. Uma imagem representativa dos iNs e iOPCs cultivados em co-cultura após 7 dias de neurônios é mostrada na Figura 3A.
- Use neurônios congelados preparados conforme descrito acima na etapa 1.2.7 para co-cultura com OPCs. Placas congelam e descongelam neurônios a uma densidade mais alta de 3 x 105 células por poço.
- Após o Dia 14–16 em coculturas, a formação de sinapses em iNs pode ser observada pela coloração IHC de marcadores pré e pós-sinápticos, e no Dia 21 a punção sináptica deve ser abundante (Figura 3C) e as atividades neuronais podem ser registradas de forma confiável.
- A partir do dia 21, teste as células para marcadores específicos de OL (por exemplo, MBP e PLP1). No Dia 28, normalmente observamos o fenômeno de revestimento de axônios iN por processos iOL, marcados por coloração IHC para marcadores específicos (Figura 3B; neurofilamento NF para axônios iN e MBP para processos iOPC).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Geração direta de neurônios induzidos por humanos a partir de células-tronco pluripotentes humanas
É muito importante que as células-tronco pluripotentes humanas iniciantes exibam um alto grau de pluripotência para a geração bem-sucedida de iNs ou iOPCs/iOLs. Portanto, as células devem ser coradas para marcadores específicos, como Oct4 e SOX2, antes de iniciar qualquer um dos protocolos de indução descritos no presente manuscrito (Figura 1A). Células H1 humanas foram utilizadas para a obtenção de neurônios excitatórios induzidos do prosencéfalo seguindo o protocolo previamente publicado por Zhang et al., com algumas modificações (Figura 1C)12,16,17,18. Aqui, apresentamos um protocolo no qual os iNs no Dia 2 são re-chapeados em cultura pura em solução matricial (ver Tabela de Materiais), na ausência de qualquer camada de alimentação: glia ou fibroblastos. Além dos protocolos publicados anteriormente, observamos que o congelamento de iNs no Dia 2 não afeta significativamente a viabilidade celular (~15% a 20% de morte celular após o descongelamento). Neurônios puros em cultura começarão a expressar sinapsina1 no Dia 14–16 (Figura 1D). Estabelecer uma cultura neuronal pura é muito importante porque certos fatores, por exemplo, o principal fator de risco de DA ApoE, podem ser expressos por células na camada de alimentação e isso pode confundir significativamente os resultados.
A geração de iOPC e a maturação do iOL são melhoradas pelo tratamento com DMSO
Aqui, apresentamos um protocolo rápido e eficiente que permite a geração de iOPCs em 2 semanas e iOLs maduros em 4-5 semanas (Figura 2A). Aproveitamos o método de tratamento de DMSO transitório que desenvolvemos anteriormente para aumentar a eficiência de diferenciação para células ES e iPS 29,30,31. O tratamento com DMSO enriquece o número de células na fase G1 inicial para melhor integração da sinalização, favorecendo a diferenciação. Realizamos o primeiro tratamento antes de induzir células ES humanas a gerar NPCs e o segundo tratamento antes de diferenciar NPCs em iOPCs. Podemos detectar marcadores OPC específicos (Olig2, CSPG4, NKX2.2 e PDGFRA) já 2 semanas após o plaqueamento das células ES H1 (Figura 2B,E). A população de iOPC nesta fase é bastante homogênea, com >95% de células positivas para coloração de O4 e um alto nível de imunorreatividade para outros marcadores (Figura 2B). Após o início da maturação da OL no Dia 15, normalmente podemos detectar marcadores OL específicos (MBP, O1, CLDN11 e PLP1) a partir do Dia 28 (Figura 2C,E). A expressão desses marcadores específicos do estágio correlaciona-se com o curso de desenvolvimento das células oligodendrogliais e sugere um ritmo acelerado, com os marcadores NPC diminuindo progressivamente, os marcadores OPC atingindo o pico na segunda semana e os marcadores OL elevando na terceira semana (Figura 2D,E)37. Por favor, note que este processo de maturação diversifica as populações celulares. As subpopulações em contínuo, compreendendo múltiplos estágios intermediários entre OPCs e oligodendrócitos mielinizantes maduros, podem estar presentes e representar uma porcentagem variável de células totais, com células mais maduras dominando em um momento posterior.
Como comparação, compramos os iOPCs altamente referenciados e os amadurecemos no iOL seguindo o protocolo do fabricante. Testamos a expressão dos marcadores mencionados acima em nossas preparações iOPC e iOL e nas células que compramos. Determinamos que as células geradas seguindo nosso protocolo apresentaram maior expressão de todos os genes testados (Figura 2E). Curiosamente, quando testamos os níveis secretados de duas isoformas principais de β amiloide (Aβ40 e Aβ42) em iNs versus iOPCs, notamos que os iOPCs secretaram mais de ambos os fragmentos, mas a proporção permaneceu a mesma (Figura 2F).
Co-cultura de iNs e iOPCs
Este protocolo é otimizado especificamente para co-cultura de iNs e iOPCs e permite o nosso monitoramento em tempo real das comunicações intercelulares entre esses dois tipos de células ao longo do curso do desenvolvimento neural. As densidades ideais de chapeamento para ambos os tipos de células precisam ser decididas com uma série de titulação do número de células para alcançar a diferenciação adequada (Figura 3A). Após 4 semanas em coculturas, espera-se que os iOPCs sejam adequadamente diferenciados em OLs que sejam positivos para marcadores específicos, como MBP, e estendam os processos para enxugar axônios (Figura 3B). O sistema de cocultura pode aumentar robustamente o número de sinapses, indicando que as iOPCs fornecem um suporte neuronal através de contatos físicos ou liberação de fatores tróficos (Figura 3C). Podemos manter as coculturas em condições de saúde aceitáveis por até 6 semanas e observar que o número de sinapses e outros atributos neuronais se estabilizam por volta da quinta semana. Vale ressaltar que astrócitos e micróglias não estão presentes em nossas preparações e sua ausência pode ser documentada verificando a expressão de marcadores específicos (Figura 3D). Os iOPCs expressam um bom número de genes bem caracterizados que podem potencialmente responder e mediar os sinais dependentes da atividade dos neurônios vizinhos, de forma parácrina (por exemplo, neurotrofinas e metabólitos) e/ou sináptica (Figura 3E e 3F).

Figura 1: Geração direta de neurônios induzidos por humanos (iNs) a partir de hPSCs. (A) Diagrama de fluxo da geração de iN. (B) Imagens representativas de campo brilhante e imunofluorescência da cultura inicial de células-tronco pluripotentes humanas (H1) para confirmar a pluripotência. Oct4 é mostrado em vermelho e Sox2 em verde. (C) Imagens representativas de campo brilhante de iNs no Dia 4 e Dia 6. (D) Morfologia característica para arborização dendrítica e puncta de sinapse em iNs cultivadas em cultura pura por 24 dias e coradas por coloração por imunofluorescência para marcador dendrítico Map2 e marcador pré-sináptico Sinapsina 1 (Syn1). Por favor, clique aqui para ver uma versão maior desta figura.

Figura 2: Geração de iOPC e maturação de iOL. (A) Diagrama de fluxo de geração de iOPC e iOL. (B) Imagens representativas de campo brilhante e imunofluorescência de iOPCs no Dia 15. Olig2 (marcador pan-oligodendroglia) é mostrado em verde, O4 (marcador OPC) em vermelho e DAPI em azul. O exame de imagem revelou que >95% dos iOPCs são positivos para O4 e 25% para Olig2. (C) Imagens representativas de campo brilhante e imunofluorescência de iOLs no Dia 28. O MBP é mostrado em verde, O1 em vermelho e DAPI em azul. (D) A expressão do marcador NPC PAX6 diminui drasticamente em iOPCs no Dia 14 e diminui ainda mais para o fundo em OLs no Dia 28, indicando uma transdiferenciação robusta de NPCs e um alto nível de homogeneidade na população de iOPC. (E) O perfil de expressão tempo-curso de genes marcadores comuns de OPC e OL em culturas geradas pelo protocolo descrito, sem (-DMSO) ou com (+DMSO) a etapa de incubação do DMSO (etapas 2.1.3 e 2.2.2), ensaiado em diferentes pontos de tempo. Como comparação, os iOPCs comerciais (ver Tabela de Materiais) foram amadurecidos de acordo com as instruções do fabricante, e ambos os iOPCs (iOPC-Tempo) ou iOLs (iOL-Tempo) foram testados para os mesmos marcadores. Como esperado, a PAM (um marcador de oligodendrócitos maduros) não foi detectada (N.D.) nos estágios iniciais de diferenciação em todas as iOPCs testadas. O DMSO aumentou significativamente a eficiência da diferenciação OPC e da maturação do OL. (F) A produção e secreção de Aβ40 e Aβ42 em culturas de iNs e iOPCs puras, medidas por kits comerciais de ELISA (ver Tabela de Materiais) em sobrenadantes obtidos de culturas puras de iNs e iOPCs tanto no Dia 15 quanto normalizadas por números celulares (ambos na densidade de 200.000 células por poço em uma placa de 24 poços). Por favor, clique aqui para ver uma versão maior desta figura.
Os dados em gráficos de barras são plotados como média ± MEV (n ≥ 3). A significância estatística foi avaliada pelo teste t de Student (*, p < 0,05; ***, p < 0,001); em (D), em comparação com o NPC; em (E), em comparação com o controle iOPC-Tempo; em (F), em comparação com iN.

Figura 3: Co-cultura de iNs e iOPCs. (A) Imagem representativa de campo brilhante de iNs e iOPCs co-cultivados no Dia 7, mostrando uma densidade adequada para maturação adicional. (B) Imagem de imunofluorescência representativa de iNs e iOPCs co-cultivados por 28 dias. O marcador axonal neurofilamento NF é mostrado em verde e o marcador oligodendrocítico MBP em vermelho. À direita, um segmento do axônio iN revestido pelo processo iOL (MBP+). (C) Formação de sinapse ensaiada em coculturas de 4 semanas de idade. As células foram coradas para Sinapsina 1 (Syn1, verde) e MAP2 (vermelho), e os punctos sinápticos foram quantificados por análise confocal da densidade ao longo dos segmentos dendríticos, conforme descrito17,18. (D) Em nossas coculturas de iNs e iOPCs (7 dias de co-cultura), a expressão de marcadores de astrócitos, ALDHL1 e GFAP, é mínima (acima), e a expressão de marcadores de micróglia, TMEM119, TREM2 e CD33, não é detectada (N.D.) por qPCR. A contaminação destes dois tipos de células gliais é, portanto, excluída. (E) A cocultura de iOPC com iN leva à formação de sinapses neurônio-OPC. O marcador pós-sináptico PSD95-mCherry marcado com fluorescência é expresso apenas em OPCs, e exibe um padrão difuso em culturas únicas (esquerda), mas agregado para formar puncta em coculturas (direita, indicado por setas; Tuj1, marcador neuronal). (F) A expressão de genes oligodendrogliais bem caracterizados que podem sentir e responder a atividades neuronais nas culturas puras de iOPCs no Dia 14. Por favor, clique aqui para ver uma versão maior desta figura.
Os dados em gráficos de barras são plotados como média ± MEV (n ≥ 3). A significância estatística foi avaliada pelo teste t de Student (**, p < 0,005; ***, p < 0,001); em (C), em comparação com a condição sem OPC; em (D), em comparação com os astrócitos primários no painel superior.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Além do suporte físico e metabólico para estabilizar as estruturas das sinapses e facilitar a condução do sinal salino por mielinização, as células da linhagem de oligodendrócitos podem moldar o padrão de atividade neuronal por meio de conversas cruzadas rápidas e dinâmicas com neurônios 5,6,7. Enquanto na patologia da DA as respostas oligodendrogliais foram inicialmente consideradas meramente secundárias à inflamação e estresses oxidativos, agora há evidências promissoras argumentando que a integridade comprometida da mielina é um evento patogênico precoce antes do aparecimento da agregação Aβ e da hiperfosforilação da tau9. Além disso, o reparo da mielinização através da auto-renovação de OPCs é particularmente vulnerável na AD38, um processo que depende fortemente das atividades neuronais. Compreender o mecanismo para apoiar a sinalização saudável de neurônios-oligodendrócitos representa, portanto, uma excelente oportunidade para a identificação de novos alvos terapêuticos.
O protocolo Ngn2 de fator de transcrição único é uma das técnicas mais empregadas para a geração de neurônios humanos derivados de células-tronco, e os procedimentos descritos aqui são refinamentos adicionais para a obtenção de culturas neuronais puras. Nosso protocolo iOPC/iOL tem um período de indução menor do que os estudos publicados anteriormente (4 a 24 semanas), com um rendimento robusto e pureza comparável a outros protocolos comumente utilizados 19,20,22,23,24,25,26,27,28 . Nosso protocolo introduz a diferenciação gradual de células ES em NPCs, OPCs e, finalmente, oligodendrócitos por meio de pistas de padronização caracterizadas, e gera células funcionais que podem ser usadas para estudar a regulação da homeostase da mielinização e reparo in vitro ou in vivo (por exemplo, por enxerto no modelo de camundongo com tremores), conforme descrito no trabalho anterior. A melhoria em nosso protocolo é muito promovida pela incubação do DMSO, que ativa a proteína do retinoblastoma e prolonga a fase G1 do ciclo celular para melhor integrar os estímulos de diferenciação dirigida, além de potencializar a diferenciação terminal em derivados funcionais29,30. Por fim, o uso de clemastina, composto muscarínico e anti-histamínico atingido identificado por meio de telas medicamentosas para terapêutica de remielinização33, também encurta a maturação dos oligodendrócitos, como observado no preparo celular de iPS e animais vivos 7,28.
As limitações da técnica residem principalmente na discrepância intrínseca entre as configurações in vitro simplificadas e os meios in vivo no cérebro; essa discrepância leva a um desconto no pleno potencial de desenvolvimento nos estágios avançados para tipos individuais de células cerebrais. Para os iNs, estudos recentes foram capazes de manter as culturas em boa saúde sináptica por um período de tempo consideravelmente longo, mas ainda revelaram alguma imaturidade relativa manifestada como estruturas reduzidas semelhantes à coluna vertebral e transmissão sináptica espontânea prejudicada em culturas "velhas" de iN (mesmo as de 25 meses de idade)39. Embora as iOPCs tenham sido reprodutivelmente demonstradas para mielinizar axônios in vivo após serem transplantadas em cérebros de camundongos transgênicos, os ensaios de mielinização in vitro com avaliação microscópica eletrônica ainda representam um desafio técnico com eficiência insatisfatória para quase todos os protocolos publicados19,28, incluindo este. Portanto, nosso sistema de co-cultura neurônio-OPC não está previsto para imitar fielmente o processo de envelhecimento cerebral, bem como o estágio tardio da patologia da DA. Em vez disso, está singularmente posicionado para desembaraçar as elaboradas interações intercelulares entre neurônios e OPCs ou oligodendrócitos em estágio inicial, que são independentes da mielinização e ainda fundamentais para o desenvolvimento neural adequado e a patogênese da doença.
Cada um dos três procedimentos descritos aqui tem suas etapas c ríticas dentro do protocolo e pode exigir modificações e solução de problemas. Para o protocolo iN, existem duas etapas críticas: seleção de puromicina (etapa 1.2.5) e densidade de chapeamento (etapa 1.2.6). A remoção incompleta de células subtransduzidas resulta na contaminação de células pouco diferenciadas e compromete a sobrevivência e as funções neuronais. As modificações para uma seleção mais forte de puromicina com maior concentração e incubação mais longa, conforme descrito na etapa 1.2.5, teriam que ser consideradas. A densidade de revestimento adequada deve ser determinada por titulação para cada linhagem celular pluripotente, pois a baixa densidade leva ao colapso das culturas e a alta densidade estimula a agregação celular e impede o crescimento neuronal. Para o protocolo iOPC/iOL, as duas etapas críticas são o controle da proliferação celular na diferenciação OPC (etapa 2.2.3) e a densidade de chapeamento para maturação OL (etapas 2.2.4 e 2.3.2). O crescimento excessivo de NPCs diferenciadores sinaliza uma resposta fraca aos estímulos de diferenciação de OPC e precisa ser atenuado por uma dosagem apropriada de tratamento com Ara-C (dentro da faixa indicada). Ao plaquear OPCs para maturação, uma faixa mais baixa de densidade celular é preferida aqui, pois a distribuição esparsa pode facilitar a indução de uma morfologia fisiológica de estruturas complexas (como mostrado na Figura 2C). Para o protocolo de co-cultura iN-iOPC, gostaríamos de chamar a atenção para a etapa crítica do revestimento com uma densidade apropriada para ambos os tipos de células (etapas 3.1.1 e 3.2.2). Especificamente, os iNs podem não se ligar bem à superfície entre os OPCs em crescimento e tendem a se desprender primeiro quando a cultura atinge a confluência. A razão ideal teria que ser decidida pela titulação dos números de células.
No geral, essa abordagem reducionista que reside em nossos protocolos é uma ferramenta poderosa para dissecar as interações heterocelulares específicas da complexidade inerente do cérebro humano e servir para descobrir a biologia oligodendroglial na saúde e na DA. A importância em relação aos métodos existentes é, portanto, bastante aparente em nossa opinião. Uma utilidade adicional dos métodos aqui desenvolvidos para as futuras aplicações é a terapia baseada em células para condições desmielinizantes, como pós-radioterapia40 e lesão medular41,42. Além disso, a capacidade de alto rendimento deste sistema baseado em células-tronco também pode ser utilizada em maior escala para rastrear bibliotecas de pequenas moléculas em busca de compostos que possam proteger ou restaurar o estado fisiológico de neurônios, OPCs, oligodendrócitos e suas interações. Assim, acreditamos que os protocolos aqui descritos facilitarão o trabalho futuro no desenvolvimento de melhores ferramentas de modelagem e tratamentos eficazes para a DA e outros distúrbios neurodegenerativos.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Os autores não têm nada a revelar.
Acknowledgments
Este trabalho foi apoiado pelas subvenções dos Institutos Nacionais de Saúde (R00 AG054616 para Y.A.H. e T32 GM136566 para K.C.), Stanford University School of Medicine e uma Siebel Fellowship (concedida a S.C.). Y.A.H. é professor translacional da GFL do Centro de Neurociência Translacional do Brown Institute for Translational Sciences.
Materials
| Name | Company | Catalog Number | Comments |
| Accutase | STEMCELL Technologies | 7920 | |
| B27 supplement | ThermoFisher | 17504044 | |
| bFGF | ThermoFisher | PHG 0266 | |
| cAMP | MilliporeSigma | A9501 | |
| Clemastine | MilliporeSigma | SML0445 | |
| DMEM/F12 medium | STEMCELL Technologies | 36254 | |
| DMSO | ThermoFisher | D12345 | |
| Doxycycline | MilliporeSigma | D3072 | |
| Fetal Bovine Serum | ScienCell | 10 | |
| H1 human ES cells | WiCell | WA01 | |
| Matrigel | Corning | 354234 | |
| mTeSR plus | STEMCELL Technologies | 5825 | |
| N2 supplement | ThermoFisher | 17502001 | |
| Neurobasal A medium | ThermoFisher | 10888-022 | |
| Non Essential Amino Acids | ThermoFisher | 11140-050 | |
| PDGF-AA | R&D Systems | 221-AA-010 | |
| PEI | VWR | 71002-812 | |
| pMDLg/pRRE | Addgene | 12251 | |
| Polybrene | MilliporeSigma | TR-1003-G | |
| pRSV-REV | Addgene | 12253 | |
| Puromycin | ThermoFisher | A1113803 | |
| ROCK Inhibitor Y-27632 | STEMCELL Technologies | 72302 | |
| SAG | Tocris | 4366 | |
| STEMdiff Neural Progenitor Freezing Media | STEMCELL Technologies | 5838 | |
| STEMdiff SMADi Neural Induction Kit | STEMCELL Technologies | 8581 | |
| T3 triiodothyronine | MilliporeSigma | T6397 | |
| Tempo-iOlogo: Human iPSC-derived OPCs | Tempo BioScience | SKU102 | |
| TetO-Ng2-Puro | Addgene | 52047 | |
| VSV-G | Addgene | 12259 |
References
- Pelvig, D. P., Pakkenberg, H., Stark, A. K., Pakkenberg, B. Neocortical glial cell numbers in human brains. Neurobiology of Aging. 29 (11), 1754-1762 (2008).
- Barres, B. A. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 60 (3), 430-440 (2008).
- De Strooper, B., Karran, E. The cellular phase of Alzheimer's disease. Cell. 164 (4), 603-615 (2016).
- Monje, M. Myelin plasticity and nervous system function. Annual Review of Neuroscience. 41, 61-76 (2018).
- Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., Bergles, D. E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nature Neuroscience. 21 (5), 696-706 (2018).
- Gibson, E. M., et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science. 344 (6183), 1252304 (2014).
- Pan, S., Mayoral, S. R., Choi, H. S., Chan, J. R., Kheirbek, M. A. Preservation of a remote fear memory requires new myelin formation. Nature Neuroscience. 23 (4), 487-499 (2020).
- Thornton, M. A., Hughes, E. G. Neuron-oligodendroglia interactions: Activity-dependent regulation of cellular signaling. Neuroscience Letters. 727, 134916 (2020).
- Ettle, B., Schlachetzki, J. C. M., Winkler, J. Oligodendroglia and myelin in neurodegenerative diseases: more than just bystanders. Molecular Neurobiology. 53 (5), 3046-3062 (2016).
- Essayan-Perez, S., Zhou, B., Nabet, A. M., Wernig, M., Huang, Y. A. Modeling Alzheimer's disease with human iPS cells: advancements, lessons, and applications. Neurobiology of Disease. 130, 104503 (2019).
- Li, L., et al. GFAP mutations in astrocytes impair oligodendrocyte progenitor proliferation and myelination in an hiPSC model of Alexander disease. Cell Stem Cell. 23 (2), 239-251 (2018).
- Lin, Y. T., et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer's disease phenotypes in human iPSC-derived brain cell types. Neuron. 98 (6), 1294 (2018).
- TCW, J., et al. Cholesterol and matrisome pathways dysregulated in human APOE ε4 glia. bioRxiv. , (2019).
- Ang, C. E., Wernig, M.
- Penney, J., Ralvenius, W. T., Tsai, L. H. Modeling Alzheimer's disease with iPSC-derived brain cells. Molecular Psychiatry. 25 (1), 148-167 (2020).
- Zhang, Y., et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron. 78 (5), 785-798 (2013).
- Huang, Y. A., Zhou, B., Nabet, A. M., Wernig, M., Sudhof, T. C. Differential signaling mediated by ApoE2, ApoE3, and ApoE4 in human neurons parallels Alzheimer's Disease risk. Journal of Neuroscience. 39 (37), 7408-7427 (2019).
- Huang, Y. A., Zhou, B., Wernig, M., Sudhof, T. C. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Abeta Secretion. Cell. 168 (3), 427-441 (2017).
- Yang, N., et al. Generation of oligodendroglial cells by direct lineage conversion. Nature Biotechnology. 31 (5), 434-439 (2013).
- Douvaras, P., et al. Efficient generation of myelinating oligodendrocytes from primary progressive multiple sclerosis patients by induced pluripotent stem cells. Stem Cell Reports. 3 (2), 250-259 (2014).
- Lee, E. H., Park, C. H. Comparison of reprogramming methods for generation of induced-oligodendrocyte precursor cells. Biomolecules & Therapeutics (Seoul). 25 (4), 362-366 (2017).
- Ehrlich, M., et al. Rapid and efficient generation of oligodendrocytes from human induced pluripotent stem cells using transcription factors. Proceedings of the National Academy of Sciences of the United States of America. 114 (11), 2243-2252 (2017).
- Rodrigues, G. M. C., et al. Defined and scalable differentiation of human oligodendrocyte precursors from pluripotent stem cells in a 3D culture system. Stem Cell Reports. 8 (6), 1770-1783 (2017).
- Hu, B. Y., Du, Z. W., Li, X. J., Ayala, M., Zhang, S. C. Human oligodendrocytes from embryonic stem cells: conserved SHH signaling networks and divergent FGF effects. Development. 136 (9), 1443-1452 (2009).
- Izrael, M., et al. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Molecular and Cellular Neuroscience. 34 (3), 310-323 (2007).
- Yamashita, T., et al. Differentiation of oligodendrocyte progenitor cells from dissociated monolayer and feeder-free cultured pluripotent stem cells. PLoS One. 12 (2), 0171947 (2017).
- Wang, S., et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell. 12 (2), 252-264 (2013).
- Chanoumidou, K., Mozafari, S., Baron-Van Evercooren, A., Kuhlmann, T. Stem cell derived oligodendrocytes to study myelin diseases. Glia. 68 (4), 705-720 (2020).
- Chetty, S., et al. A simple tool to improve pluripotent stem cell differentiation. Nature Methods. 10 (6), 553-556 (2013).
- Li, J., et al. A transient DMSO treatment increases the differentiation potential of human pluripotent stem cells through the Rb family. PLoS One. 13 (12), 0208110 (2018).
- Sambo, D., Li, J., Brickler, T., Chetty, S. Transient treatment of human pluripotent stem cells with DMSO to promote differentiation. Journal of Visualized Experiments: JoVE. (149), (2019).
- Douvaras, P., Fossati, V. Generation and isolation of oligodendrocyte progenitor cells from human pluripotent stem cells. Nature Protocols. 10 (8), 1143-1154 (2015).
- Mei, F., et al. Micropillar arrays as a high-throughput screening platform for therapeutics in multiple sclerosis. Nature Medicine. 20 (8), 954-960 (2014).
- Madhavan, M., et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nature Methods. 15 (9), 700-706 (2018).
- Zhang, Y., et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 89 (1), 37-53 (2016).
- Grubman, A., et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer's disease reveals cell-type-specific gene expression regulation. Nature Neuroscience. 22 (12), 2087-2097 (2019).
- Goldman, S. A., Kuypers, N. J. How to make an oligodendrocyte. Development. 142 (23), 3983-3995 (2015).
- Behrendt, G., et al. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia. 61 (2), 273-286 (2013).
- Patzke, C., et al. Neuromodulator signaling bidirectionally controls vesicle numbers in human synapses. Cell. 179 (2), 498-513 (2019).
- Piao, J., et al. Human embryonic stem cell-derived oligodendrocyte progenitors remyelinate the brain and rescue behavioral deficits following radiation. Cell Stem Cell. 16 (2), 198-210 (2015).
- Keirstead, H. S., et al. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. Journal of Neuroscience. 25 (19), 4694-4705 (2005).
- Kim, D. S., et al. Rapid generation of OPC-like cells from human pluripotent stem cells for treating spinal cord injury. Experimental & Molecular Medicine. 49 (7), 361 (2017).
Tags
Neurociência Edição 165 células-tronco pluripotentes induzidas células iPS células-tronco embrionárias humanas células ES neurônios oligodendrócitos células progenitoras de oligodendrócitos OPCs doença de Alzheimer peptídeos beta-amilóides sinapsesErratum
Formal Correction: Erratum: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions
Posted by JoVE Editors on 12/29/2020.
Citeable Link.
An erratum was issued for: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions. The Representative Results section has been updated.
Figure 3 was updated from:
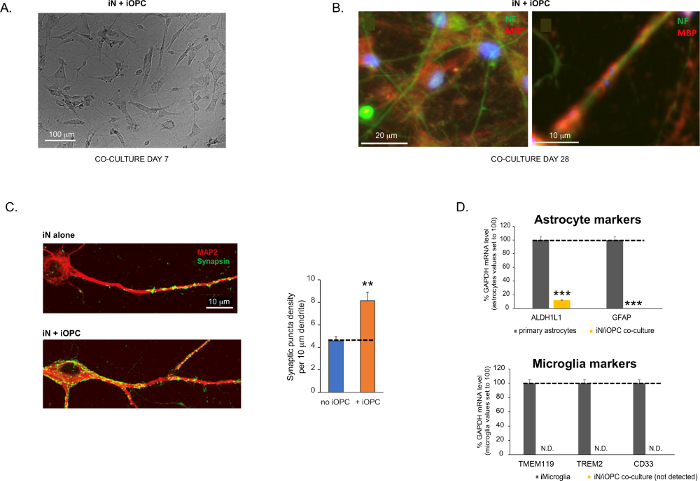
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. Please click here to view a larger version of this figure.
to:
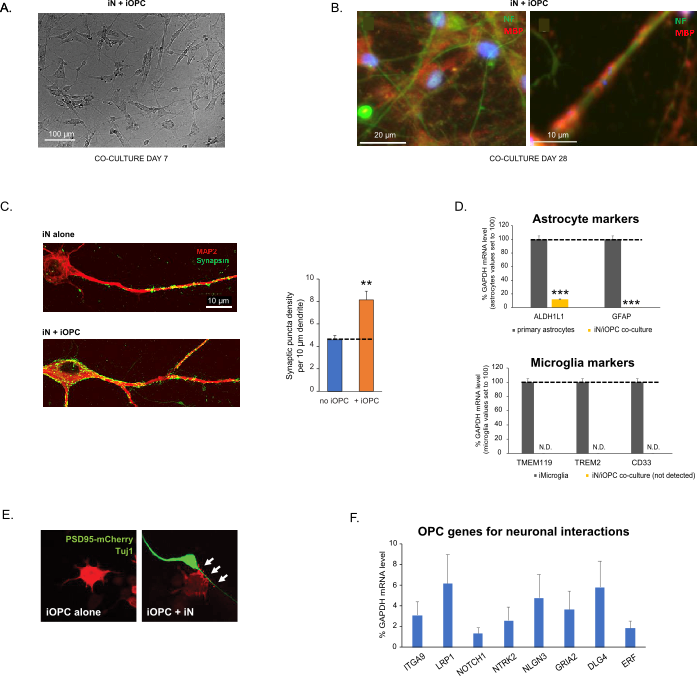
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. (E) Coculturing iOPC with iN leads to the formation of neuron-OPC synapses. The fluorescence-tagged post-synaptic marker PSD95-mCherry is expressed only in OPCs, and display a diffuse pattern in single cultures (left) but aggregate to form puncta in cocultures (right, indicated by arrows; Tuj1, neuronal marker). (F) The expression of well-characterized oligodendroglial genes that can sense and respond to neuronal activities in the pure cultures of iOPCs at Day 14. Please click here to view a larger version of this figure.
The fourth paragraph was updated from:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D).
to:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D). The iOPCs express a good number of well-characterized genes that can potentially respond to and mediate the activity-dependent signals from neighboring neurons, in a paracrine (e.g. neurotrophins and metabolites) and/or a synaptic manner (Figure 3E and 3F).
