

Summary
介绍并讨论了 体外 肌动球蛋白束的形成和使用光学镊子测量肌球蛋白集合力的产生。
Abstract
肌球蛋白是水解ATP以沿着肌动蛋白丝(AF)轨道行走的运动蛋白,在运动和肌肉收缩等细胞过程中至关重要。为了了解它们的力产生机制,肌球蛋白II已经在单分子(SM)水平和使用生物物理方法(如光学捕获)在 体外作为 电机团队进行了研究。
这些研究表明,当从三珠排列中的单分子水平移动到滑动排列中的刚性珠或盖玻片表面上一起工作的电机组时,肌球蛋白力的产生行为可能会有很大差异。然而,这些测定结构不允许像在细胞内那样评估粘弹性结构层次结构中肌球蛋白的组动力学。我们开发了一种使用光学镊子的方法,以研究肌球蛋白系综与多个肌动蛋白丝相互作用的力产生机制。
这些肌动肌肽束有助于在分层和顺应的环境中进行研究,从而捕获运动通信和集合力输出。该测定的可定制性质允许改变实验条件,以了解对肌球蛋白集合、肌动蛋白丝束或周围环境的修饰如何导致不同的力输出。
Introduction
运动蛋白对生命至关重要,将化学能转化为机械功1,2,3。肌球蛋白马达通过沿着类似于轨道的细丝采取步骤与肌动蛋白丝相互作用,肌动蛋白-肌球蛋白网络的动力学执行肌肉收缩、细胞运动、细胞分裂过程中的收缩环以及细胞内货物的运动,以及其他基本任务3,4,5,6,7,8.由于肌球蛋白具有如此多的重要作用,肌球蛋白-肌动蛋白网络功能衰竭可导致疾病发展,例如肌球蛋白重链突变导致肥厚型心肌病(HCM)的心脏过度收缩9,10,11,12,13,14.在肌肉收缩中,单个肌球蛋白马达通过作为一个整体相互合作,以提供所需的机械能,执行AFs4,15,16,17,18的相对滑动。肌球蛋白马达在AF之间形成交叉桥,并利用由于其机械化学循环引起的构象变化共同向对齐的细丝17,18,19,20,21的倒刺端移动。
使用光学捕获等技术在SM水平上开发定量体外运动测定有助于收集有关单个肌球蛋白马达如何工作的前所未有的细节,包括测量SM力的产生和步长 22,23,24,25,26,27,28,29,30.Finer等人开发了“三珠”或“哑铃”光学捕获测定法,以探测单个肌球蛋白II电机的力产生机制23,31。由于肌肉肌球蛋白II在团队中工作以收缩AF,但在SM水平上是非过程性的,因此必须从经典的电机结合珠方法32重新排列光学捕获测定方向。为了形成哑铃测定,使用两个光学陷阱将AF固定在与盖玻片连接的珠子上的肌球蛋白马达上,并通过陷阱23内的AF运动测量单个电机输出的力。
然而,SM力和使用单电机/单丝测定方向并不能给出系统级力产生的完整图像,因为许多运动蛋白(包括肌球蛋白II)不能单独工作,并且通常不能作为其各部分的总和起作用15,16,17,32,33,34,35,36.更复杂的结构,包括一个以上的电机与不止一根细丝相互作用,对于更好地了解肌球蛋白和肌动蛋白丝网络的协同作用是必要的15,32。哑铃测定方向已被利用来研究小集合力的产生,方法是将多个肌球蛋白附着在珠子上或使用肌球蛋白厚的细丝连接到表面并允许电机与悬浮的AF4,23,34,37,38,39,40相互作用。
其他小型集合测定包括体外细丝滑动测定,其中肌球蛋白马达被涂覆在盖玻片表面上,并且使用与AF结合的珠子来探测电机组产生的力4,35,36,38,39,40,41,42,43.在这两种情况下,肌球蛋白都与刚性表面(珠子或盖玻片)结合并利用一个AF。在这些情况下,电机不能自由移动或相互通信,肌球蛋白刚性结合也不能反映电机在肌节中一起工作的顺从的分层环境32。先前的研究表明,肌球蛋白II可以通过改变力产生和占空比41,44,45等特性来感知其环境并相应地适应不断变化的粘弹性或运动浓度条件。因此,需要开发一种光学捕获测定法,以促进和捕获运动通信和系统顺应性,以更真实地描绘肌球蛋白II集合力生成的机制基础。
在这里,我们开发了一种通过形成由两个肌动蛋白丝之间相互作用的多个肌球蛋白马达组成的肌球蛋白束或三明治来将体外分层结构与光学捕获偶联的方法。这种模块化检测几何形状能够直接探测分子和环境因素如何影响集合肌球蛋白力的产生。此外,通过这些肌动蛋白 - 肌球蛋白集合研究力产生机制有可能帮助建模和理解大规模细胞任务(如肌肉收缩)如何从分子水平9,10,13向上传播。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 蚀刻盖玻片
- 将 100 g KOH 溶解在 1,000 mL 烧杯中的 300 mL 100% 乙醇中。用搅拌棒搅拌,直到大部分 KOH 溶解。
注意:浓缩的KOH溶液会导致灼伤和衣服损坏。戴上手套、护目镜和实验室外套。 - 将盖玻片单独放入盖玻片清洁架中。
注意:机架设计有狭缝,可将单个盖玻片隔开,以便在盖玻片的每个面、底部的排水孔上进行蚀刻和冲洗,并由可以承受恶劣蚀刻条件的材料制成。它们可以定制或商业购买。 - 准备并标记三个 1,000 mL 烧杯:一个装有 300 mL 乙醇,两个装有 300 mL 反渗透 (RO) 水的烧杯。
注意:在这里,RO水来自实验室净水器,但如果当地没有净化器,也可以商业购买。 - 将四个烧杯中的每一个放入浴超声仪中脱气5分钟。
- 将一排盖玻片浸入KOH和乙醇的烧杯中,超声处理5分钟。
- 将盖玻片架从KOH/乙醇烧杯转移到仅乙醇烧杯中。在烧杯中上下浸入架子,直到没有串珠。
注意:注意不要打扰盖玻片或用力将架子放入烧杯中。这将导致盖玻片从机架中脱落或引起化学品飞溅。 - 小心地将盖玻片架从乙醇烧杯转移到水杯中,上下浸入直到没有珠子。
- 将盖玻片的架子浸入尚未使用的烧杯中,然后再次超声处理5分钟。
- 用瓶子用水喷洒盖玻片架,直到它顺利地从盖玻片上流下来。用乙醇重复。
- 将架子放入90°C的烤箱中干燥20分钟。将蚀刻盖玻片的架子在室温下存放在密闭容器中,以防止在使用前受到污染。
2. 肌动蛋白丝聚合
- 制作解决方案 T
- 在 50 mL 锥形管中,加入 3.94 g Tris-HCl 和 0.147 g CaCl2。加入RO水,使总体积为50 mL并充分混合。
注意:溶液T的最终浓度分别为500mM Tris-HCl和20mM CaCl2 。 - 标记管溶液T并将其储存在4°C。
- 在 50 mL 锥形管中,加入 3.94 g Tris-HCl 和 0.147 g CaCl2。加入RO水,使总体积为50 mL并充分混合。
- 制作 TC 缓冲区
- 在 50 mL 锥形管中混合 40 mL RO 水和 1.5 mL 溶液 T。通过添加少量浓缩KOH将pH值更改为8.0。加水制成 50 mL 溶液,并验证 pH 值。如果需要,调整pH值。
注意:最终的TC缓冲液含有pH 8时的5 mM Tris-HCl和0.2 mM CaCl2 。 - 标记管TC并将其储存在4°C。
- 在 50 mL 锥形管中混合 40 mL RO 水和 1.5 mL 溶液 T。通过添加少量浓缩KOH将pH值更改为8.0。加水制成 50 mL 溶液,并验证 pH 值。如果需要,调整pH值。
- 制作 FC 缓冲区
- 将 85 mL RO 水、10 mL 溶液 T、3.73 g KCl 和 0.041 g MgCl2 加入 100 mL 缓冲瓶中。通过添加少量浓缩KOH将pH值修改至7.5。加水使最终体积为 100 mL 并验证 pH 值。
注意:最终的FC缓冲液含有pH 7.5的500 mM Tris-HCl,500 mM KCl,2 mM MgCl2和2 mM CaCl2 。 - 标记管FC并将其储存在4°C。
- 将 85 mL RO 水、10 mL 溶液 T、3.73 g KCl 和 0.041 g MgCl2 加入 100 mL 缓冲瓶中。通过添加少量浓缩KOH将pH值修改至7.5。加水使最终体积为 100 mL 并验证 pH 值。
- 准备一般肌动蛋白缓冲液(GAB)。
- 在微量离心管中混合 485 μL TC 缓冲液、10 μL 10 mM ATP 和 5 μL 50 mM DTT。
注意:最终缓冲条件为 5 mM Tris-HCl、0.2 mM CaCl 2、0.5 mM DTT 和0.2 mM ATP。 - 将其标记为GAB并将其储存在4°C。
- 在微量离心管中混合 485 μL TC 缓冲液、10 μL 10 mM ATP 和 5 μL 50 mM DTT。
- 制备肌动蛋白聚合缓冲液(APB)。
- 在微量离心管中混合 455 μL FC 缓冲液、25 μL 100 mM ATP 和 20 μL 50 mM DTT。
注意:最终缓冲条件为 50 mM Tris-HCl、500 mM KCl、2 mM MgCl 2、2 mM CaCl 22 mM DTT 和 5 mM ATP。 - 将管标记为APB并将其储存在4°C。
- 在微量离心管中混合 455 μL FC 缓冲液、25 μL 100 mM ATP 和 20 μL 50 mM DTT。
- 重组肌动蛋白
- 通过将 100 μL 去离子水添加到 1 mg 冻干肌动蛋白小瓶中来重建兔骨骼肌肌动蛋白。通过轻轻上下移液充分混合。分装到 5 μL 样品中,快速冷冻,并将 10 mg/mL 肌动蛋白等分试样储存在 -80 °C。
- 通过加入 20 μL RO 水重建生物素化的兔骨骼肌肌动蛋白。分装到 5 μL 样品中,快速冷冻,并将 1 mg/mL 生物素化的肌动蛋白等分试样储存在 -80 °C。
- 非标记肌动蛋白聚合与罗丹明鬼笔环肽稳定
- 解冻一瓶10mg / mL肌动蛋白并将其放在冰上。
- 准备新鲜的 GAB 缓冲液,向肌动蛋白等分试样中加入 100 μL GAB,然后轻轻上下移液混合。将溶液在冰上孵育1小时。
- 在孵育期间准备新鲜的APB。孵育后,通过将 11 μL APB 添加到肌动蛋白溶液中将肌动蛋白聚合成细丝。通过轻轻上下移液充分混合。放在冰上20分钟。
- 将 5 μL 罗丹明标记的鬼笔环肽添加到新鲜聚合的肌动蛋白丝溶液中。在黑暗中在冰上放置1小时。
- 将包裹在铝箔中的罗丹明肌动蛋白小瓶在4°C的黑暗中储存。
注意:建议使用这些灯丝最多 1 周。通过对仅包含自动对焦的流通池进行快速成像并每天查看一致的细丝,可以每天确认自动对焦质量。
- 生物素化肌动蛋白聚合与Alexa Fluor 488鬼笔环肽稳定
- 解冻一瓶 10 mg/mL 肌动蛋白和 1 瓶 1 mg/mL 生物素化肌动蛋白,并将它们放在冰上。
- 制作新鲜的GAB缓冲液。
- 将两个小瓶(步骤2.8.1)以10:1的肌动蛋白:生物素化肌动蛋白比例合并。向肌动蛋白混合物中加入 100 μL GAB,并通过轻轻上下移液充分混合。在冰上孵育1小时。
- 在孵育过程中制作新鲜的APB。
- 孵育步骤后,通过将 11 μL APB 添加到肌动蛋白溶液中来聚合肌动蛋白。轻轻上下移液,充分混合。在冰上孵育20分钟。
- 加入 5 μL Alexa Fluor 488 标记的鬼笔环肽,并在冰上避光孵育 1 小时。
- 将包裹在铝箔中的生物素化肌动蛋白小瓶在4°C的黑暗中储存。
注意:这些灯丝最多可以使用 1 周。
3. 肌球蛋白和珠子制备
- 重建肌球蛋白 II
- 使用标准微型离心机短暂旋转(~5秒)冻干骨骼肌球蛋白II,将其收集在管底部。
- 通过加入 100 μL 在 RO 水中制备的 1 mM DTT,将肌球蛋白复溶至 10 mg/mL。
- 通过将 10 μL 10 mg/mL 肌球蛋白添加到 90 μL 1 mM DTT 的 RO 水中,将原样肌球蛋白溶液稀释 10 倍。制作小体积(1-5μL)等分试样,快速冷冻,并储存在-80°C。
注意:肌球蛋白活性可以通过执行之前发表的标准滑动丝测定来确认46,47。有关简要说明,请参阅讨论。
- 清洁链霉亲和素包被的磁珠
- 将 20 μL 1 μm 链霉亲和素珠稀释到 80 μL RO 水中。以 9,600 × g 离心并重新配入 100 μL RO 水中洗涤四次。
- 以40%振幅超声处理2分钟,并将洗涤的珠子储存在4°C的旋转器上。
4. 流通池制备
- 通过将 30 mL 100% 乙醇加入 50 mL 管中,并在水中加入 200 μL 0.1% w/v 聚 l-赖氨酸并充分混合来制备聚 l-赖氨酸溶液 (PLL)。
- 将蚀刻的盖玻片添加到PLL溶液中,使其浸泡15分钟。用镊子取下盖玻片,注意只接触盖玻片的边缘,因为它从管中拉起(见图1A-C)。用戴手套的手抓住盖玻片的边缘。
- 用过滤的空气器擦干盖玻片,直到盖玻片上没有乙醇残留物。
- 将两片双面胶带贴在显微镜载玻片的中间,彼此相距3-4毫米。撕下或切断悬挂在载玻片边缘的多余胶带。
- 将PLL涂层的盖玻片添加到垂直于显微镜载玻片长轴(形成T形)的胶带顶部以形成通道。
- 使用一根小管将盖玻片压缩到胶带上,显微镜载玻片彻底,直到胶带透明(图1A)。确保胶带中没有气泡,因为这会导致流道泄漏。
注意:流通池可容纳 10-15 μL 的体积。
5.肌动球蛋白束制备
- 在单独的管中,通过将 0.5 μL 相对标记的肌动蛋白与 300 μL APB 混合,将每种类型的肌动蛋白丝(罗丹明和生物素化的 488 标记)稀释 600 倍。向每个试管中额外添加 5 μL 相应标记的鬼笔环肽,并在黑暗中在冰上孵育 15 分钟。
- 向生物素化的肌动蛋白溶液中加入 500 mg/mL 的 1 μL β-D-葡萄糖、25 mg/mL 的 1 μL 葡萄糖氧化酶和 500 单位/mL 的 1 μL 过氧化氢酶的除氧系统。加入 1 μL 100 mM ATP 和 1 μL 100x 稀释、清洁的链霉亲和素珠。用移液器吸头轻轻搅拌。将悬浮液放在4°C的旋转器上,同时组装肌动球蛋白束的其余部分。
- 向PLL流通池中加入15 μL稀释的罗丹明肌动蛋白(图1D)。将多余的溶液吸干通过流通池,但不要让流道变干。在湿度室中孵育10分钟。
注意:湿度室可以由空的移液器吸头盒制成,底部加水,盖子上覆盖铝箔以阻挡光线。 - 在 APB 中制备 1 mg/mL 酪蛋白溶液。
- 加入 15 μL 1 mg/mL 酪蛋白,以防止后续组分的非特异性结合(图 1E)。在湿度室中孵育5分钟。
- 将所需浓度的肌球蛋白添加到步骤5.2中的生物素化肌动蛋白和珠悬浮液中。用移液器吸头轻轻搅拌,然后立即向流通池中加入 15 μL 步骤 5.2 悬浮液 + 所需的肌球蛋白浓度(图 1F,G)。孵育20分钟。用指甲油密封流通池的开口端,以防止成像和光学捕获实验期间蒸发。
注意:肌球蛋白溶液浓度为1μM可产生稳健的捆绑,并可用作所需测定定制的起点(见 图2)。
6. 使用光学陷阱(NT2纳米跟踪仪)进行力测量2
注意:虽然以下方案专门用于NT2系统,但该测定可以与其他光学捕获系统一起使用,包括那些定制的,也具有荧光功能。一般工作流程与聚焦载玻片表面、执行磁珠校准以及通过查找荧光肌动蛋白束获取数据相同。对于NT2系统,补充图S1、补充图S2、补充图S3、补充图S4、补充图 S5、补充图S6和补充图S7 提供了光捕获系统和软件界面的详细信息。
- 打开控制盒和激光(补充图S1)。
- 通过单击桌面上的 JPK纳米跟踪器 图标启动光学陷阱计算机软件。
- 通过单击中间的 罗技 按钮唤醒遥控器(补充图 S2)。
- 通过切换开/关开关打开荧光模块(补充图S3)。
- 转动滤光片立方体转盘进行明场成像(补充图S4)。
- 系统准备就绪后,使用屏幕左下角的激光功率按钮将激光器打开至 50 mW,并使其稳定 30 分钟(补充图 S5)。
- 依次单击软件中的 照明、相机、物镜 和 载物台移动 按钮,以调出这些窗口,以便在实验过程中进行查看和操作。通过单击 开/关 按钮打开显微镜照明,并通过单击并一直向右拖动条将其设置为最大功率(补充图S5)。
- 打开样品区域并从显微镜载物台上取下样品架。添加流通池,用金属样品架固定,并确保带有盖玻片的载玻片位于底部。
- 在底部物镜的中心加入30 μLRO水。不要让移液器吸头接触镜头。重新插入样品台。
注意:由于NT2系统使用水浸物镜作为捕集物镜,因此浸没介质可能会有所不同,具体取决于用户设置中的捕集物镜。 - 使用遥控器上的屏幕控制箭头或L2升高下部物镜,直到水珠接触盖玻片(补充图S5)。
- 降低顶部物镜,直到使用遥控器上的屏幕箭头或R2达到到流通池距离的一半左右。将 170 μL RO 水添加到流通池顶部的顶部,直接位于顶部物镜下方。降低顶部物镜,直到它破坏水的表面张力并形成弯月面。
- 使用遥控器上的箭头垫移动显微镜载物台,直到到达靠近流道的胶带边缘。关闭样品门。
注意:关闭样品门时“咔嗒”声表示激光快门现已打开。这是一项安全功能,仅在门关闭时允许百叶窗打开。 - 使用屏幕中的物镜窗口,通过使用屏幕上的控件单击上方箭头,将名为“激光物镜”的底部物镜向上调,从而使胶带边缘清晰对焦。 通过单击底部箭头对顶部目标执行相同的操作(补充图S5)。
注意:双箭头可更快地移动物镜或舞台。胶带的边缘用于聚焦,因为它是一个靠近盖玻片表面的大型、易于找到的物体。胶带内的气泡是另一种选择。但是,如果用户有自动例程来查找表面焦点或首选的内部方法,则不需要这样做。 - 胶带对焦后,部分关闭光学陷阱顶部的光圈。向下放置顶部物镜,直到虹膜的多边形形状可见。聚焦这些边缘,重新打开光圈,然后通过单击 挂锁 图标来耦合物镜(补充图S5)。
- 找到一个漂浮的珠子并通过单击陷阱快门按钮来捕获它,这将打开 快门 并允许捕获激光击中样品。单击屏幕上的 陷 印光标并拖动它以移动陷印激光的位置。捕获后,校准磁珠以将电压测量值与力和位移相关联。
- 单击 校准 按钮。根据功率谱分析调整校准程序,并在软件中针对 X、Y 和 Z 方向拟合转角频率(补充图 S6)。
- 单击 设置。输入磁珠的直径(1,000 nm),并在软件窗口左下角输入载物台的温度。(见 补充图S6)。
- 单击 陷阱 1。点击 X 信号。单击 “运行 ”以执行拐角频率拟合。在窗口中单击并拖动以优化功能拟合。单击“ 使用它” 以获取灵敏度和刚度值。单击接受 值。对 Y 和 Z 信号重复此操作。关闭窗口。(见 补充图S6)。
注意:其他光学捕获系统或经过用户可靠测试的定制系统上的磁珠校准程序(例如均分法或拖曳力法)也是可以接受的57,58。 - 通过搜索与盖玻片表面的 AF 结合的磁珠来查找肌动肌肽束。
- 当检测到没有其他浮珠拥挤的磁珠时,通过荧光成像观察其周围的AF以验证是否存在束。
- 通过查找共定位的两个荧光AF来验证是否存在束。打开白色光源,并使用适当的滤光片立方体通过转动转塔对每个肌动蛋白丝进行成像(Alexa Fluor 488 和罗丹明激发分别为 488 nm 和 532 nm 激发滤光片立方体)。见 补充图S4。
注意:验证单个AF荧光强度的对照实验可用于识别由单个488和单个罗丹明标记的细丝组成的束,或适用于用户选择使用的任何一组荧光团。 - 验证后,通过单击 陷阱快门 按钮捕获附着在束顶部细丝上的珠子。
- 使用屏幕上的控件通过单击 示波器 按钮来记录数据(补充图 S7)。要在不记录数据的情况下可视化测量,请单击 “开始”。要保存所有数据,请单击 自动保存。要记录测量值,请单击 “开始记录”。通过从下拉菜单中选择 X 信号或 Y 信号,选择要实时可视化的数据(位置、力、x 方向、y 方向)。请记住,xdirection 是从左到右的,y 方向在屏幕上是上下的。见 补充图S7。
注意:数据将另存为 .out 文件,包括每个方向的时间、电压、位移和力。这些文件可以导出到其他软件中进行可视化和分析。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
含有肌动球蛋白束系统的流通池采用标准设计,由显微镜载玻片和蚀刻盖玻片组成,由双面胶带制成的通道隔开(图1)。然后使用协议中所述的分阶段引入从盖玻片开始构建测定。最终测定由模板罗丹明标记的肌动蛋白丝组成;所需的肌球蛋白浓度(图 2 和 图3中的代表性结果使用1μM);生物素化,Alexa Fluor 488标记的肌动蛋白丝;1μm链霉亲和素珠;除氧系统;丁胺类兴奋剂;和 APB 缓冲区。每个流通池将形成多个束,上述肌动蛋白浓度在束之间提供足够的间距,以确保没有不必要的相互作用。这也有助于获得每个流通池的多个力测量值,以提高数据采集效率。力分布应在流通池内以及流通池与流通池之间重现。
虽然上述方案面向使用商业光学捕获装置,但此处介绍的流通池和测定可以很容易地用于不同的商业仪器或定制的光学捕获装置,与显微镜或显微镜载物台相结合,并具有荧光成像能力。根据上述方案完成所有流通池添加后,载玻片上的肌动球蛋白束(图1)即可立即测量。将流通池添加到光学陷阱显微镜载物台上,获取多个磁珠校准测量值,并通过束丝的荧光共定位来识别束。与束绑定的珠子被捕获,并开始位移和相应的力测量。用户可以在计算机监视器上实时观察数据的采集。根据流通池中使用的肌球蛋白浓度,束可以立即开始表现出实质性运动,或者可能需要30 s-1分钟才能有效地看到位移/力的增加。
代表性力曲线如图 3A 所示,其中肌球蛋白电机表现出稳定的力斜坡,然后是平台。通常看到这些类型的痕迹在2-5分钟内发展。然而,也可以测量不产生任何净力的肌动球蛋白束(图3B)。这些迹线显示为基线噪声,或者在90秒内没有表现出实质性的力净增加。这可能是由于电机的局部浓度低,不允许生产滑动,或者束处于不利的平行方向,其中灯丝的正负两端对齐。
由于流通池的内容物容易受到入射照明和捕获激光的降解、随时间推移在载玻片上的局部加热以及自由基氧的产生的影响,因此强烈建议不要使用相同的流通池超过 1 小时。为了获得最大效率,建议在获取数据时进行另一种检测孵育。位移/力迹线可以从光学捕获软件导出到Excel,Matlab,Igor或其他数据管理程序中,以进行进一步的过滤和分析。可以从这种光学捕获集合/束实验中提取的数据包括不同测定条件下不同类型的力生成曲线(基线、斜坡/平台)、力生成速度、最大力生成、集成动力学和步进行为通过步长和步长或步骤组之间的停留时间,以及占空比。用户还可以更改测定条件,以比较添加不同类型的肌球蛋白马达、添加肌动蛋白结合蛋白或改变缓冲条件如何影响这些集合力生成特性。
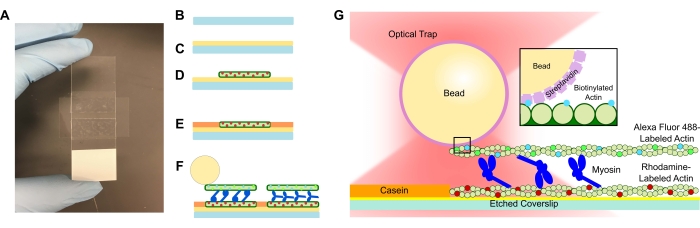
图1:测定示意图。 (A-C)蚀刻的盖玻片涂有聚-L-赖氨酸,并使用双面胶带和显微镜载玻片形成流通池。协议中描述的定时引入和孵育步骤导致罗丹明标记的鬼笔环肽稳定的肌动蛋白作为模板或底部细丝(D),然后酪蛋白阻断以防止非特异性结合(E)和(F)Alexa Fluor 488鬼笔环肽稳定的生物素化肌动蛋白作为货物或顶部细丝,以及肌球蛋白II团队在引入ATP时将细丝滑开并产生力。电机的几何形状和束内交联的性质在不同的条件下可能会有所不同,例如盐浓度59。先前的研究表明,肌球蛋白尾部结构域具有与肌动蛋白丝相互作用的能力和缓慢的集合运动46。然而,重度肌球蛋白实验中的肌球蛋白头证明每个头与相邻肌动蛋白丝60的结合。(G)链霉亲和素珠用作陷阱的光学手柄,仅与货物生物素化的肌动蛋白丝结合,这有助于验证载玻片上是否形成适当的束。请点击此处查看此图的大图。

图2:荧光肌动肌肽束。 肌动蛋白丝和束在束测定中的四种不同遭遇如图 1所示。左侧显示了带有Alexa Fluor 488鬼笔环肽通道的顶部货物生物素化肌动蛋白丝,右侧显示了带有罗丹明鬼笔环肽通道的底部模板肌动蛋白丝。在底部,显示相同的图形,并叠加彩色线条以帮助引导眼睛。(A)顶部肌动蛋白丝在底部肌动蛋白丝附近发现,但覆盖不完整。这不会用于捆绑实验。(B)顶部和底部肌动蛋白丝是共定位的,并且每根细丝的强度确认它们是束内的每根单丝。这将是捆绑实验的一个很好的候选者。(C)在底部发现了一大束自组装的罗丹明细丝。虽然有相应的顶部肌动蛋白丝是共定位的,但存在太多的底部细丝;因此,它不会用于捆绑实验。这也是当捆绑多个相同类型的肌动蛋白丝时,荧光强度增加的一个例子。用户可以将其用作判断单丝与相同灯丝类型的束的量规。(D)存在没有相应顶部灯丝的底部灯丝,也确认没有渗漏。这不会用于捆绑实验。我们注意到Alexa Fluor 488通道中的灯丝强度较低,并认为这是由于正在使用的过滤器组(蔡司的过滤器组09)。用于罗丹明通道的过滤器组是蔡司的过滤器组 43。 请点击此处查看此图的大图。

图3:肌球蛋白II集成力生成。 骨骼肌球蛋白II马达在 构建的体外 肌动蛋白结构层次结构内产生力的代表性痕迹。肌球蛋白马达协同工作,集体有效地产生力,直到达到平台期并且力持续(A)或在基线附近经历拮抗(B)。 请点击此处查看此图的大图。
补充图S1:布鲁克/ JPK纳米跟踪器2光学陷阱。 (A) 计算机显示器。(B) 电脑键盘。(三)计算机塔。(D) 控制器盒。(五)激光电源。(F)光学陷阱光学箱。(G) 倒置显微镜。(H) 显微镜载物台的门。(I) 偏振片滑块,用于在明场和微分干涉对比成像之间切换。 请点击此处下载此文件。
补充图S2:光学陷阱的遥控器。 (A) 用于定位电动载物台的键盘。(乙-丙)调整疏水阀位置。(D) A、X 和 B 分别打开和关闭主快门、陷印 1 快门和陷印 2 快门。(E) 罗技按钮用于唤醒控制器。(F) 用于定位陷印目标的向上和向下按钮。(G) 用于定位检测目标的向上和向下按钮。请注意,不需要遥控器,所有这些操作都可以在软件中完成。但是,在观察显微镜载物台环境的同时能够控制物镜和载物台位置很方便。 请点击此处下载此文件。
补充图S3:用于光学陷阱的荧光模块。 89North PhotoFluor荧光白光源耦合到倒置显微镜的背面。它通过拨动开关(箭头)打开和关闭。 请点击此处下载此文件。
补充图S4:荧光滤光片立方体转盘。 转塔(箭头)可以转动以使用在DIC,罗丹明或Alexa Fluor 488染料中成像所需的滤光片立方体。请注意,可以切换滤光片立方体以自定义使用不同荧光团的设置。 请点击此处下载此文件。
补充图S5:纳米跟踪器2软件。 (A)激光电源按钮和控制。(二)客观定位窗口。方向箭头用于移动检测物镜(顶部)和陷印物镜(底部)。双箭头以更高的速度移动目标。左下角的蓝色和红色按钮可解开物镜并将它们缩回原始位置。这对于将样品移入和取出显微镜载物台时是必要的。左起第三个按钮带有物镜和挂锁图标,将物镜“耦合”在一起,这样当它们都对焦并实现科勒照明时,用户可以在z轴上上下移动陷印物镜和检测物镜。(C)用于在x轴和y轴上移动显微镜载物台的样品定位窗口。双箭头以更高的速度移动载物台。通过单击顶部菜单中的向上/向下和向左/向右箭头图标激活此窗口。(D) 相机可视化窗口。扳手图标可用于设置自定义成像条件。此窗口通过单击顶部菜单中的 相机 图标激活。(E)显微镜照明窗口。此窗口通过单击顶部菜单中的 灯泡 图标激活。 请点击此处下载此文件。
补充图S6:校准窗口。 (一)此窗口用于磁珠校准,通过单击顶部菜单中的 Cal 图标激活。为了校准磁珠,需要在x、y和z信号中实现转折频率的最佳拟合。(B) 对于每个信号,选择左上角的相应信号按钮。(C) 单击 运行 并通过在绿色窗口 (D) 内单击并拖动来优化拟合。(E)一旦对贴合度感到满意,请单击“ 使用它” 以获取灵敏度和刚度。这将允许记录纳米位移和皮孔顿的力。(F)然后,单击左下角的 接受值 。对 y 和 z 方向重复此操作。 请点击此处下载此文件。
补充图S7:数据采集窗口。 此窗口用于获取位置和力数据,并允许用户实时查看测量值。(A) 通过单击顶部菜单中的 x,t 图标激活此窗口。(B) 用户可以在查看 x 和 y 信号之间切换。(C) 单击“开始” 开始 可视化数据。单击自动保存以 保存 数据。单击开始记录以开始 记录 和保存数据。 请点击此处下载此文件。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
使用光镊结合荧光成像进行体 外 研究,以研究肌球蛋白集合与肌动蛋白丝相互作用的动力学。肌动蛋白-肌球蛋白-肌动蛋白束使用肌球蛋白II,罗丹明肌动蛋白在束底部和盖玻片表面组装,488标记的生物素化肌动蛋白丝在束的顶部组装。来自兔肌肉的肌动蛋白使用通用肌动蛋白缓冲液(GAB)和肌动蛋白聚合缓冲液(APB)聚合和稳定。GAB 和 APB 必须每天在实验室中使用 ATP、FC 缓冲液和 TC 缓冲液新鲜制备。肌肉肌球蛋白II用于形成肌动蛋白-肌球蛋白-肌动蛋白三明治。鬼笔环肽用于肌动蛋白丝的荧光染色,以及 体外稳定。
肌球蛋白活性可以通过执行之前发表的标准滑动丝测定来确认46,47.肌球蛋白II及其亚片段可以以各种方向与盖玻片表面结合,与使用重美肌苷46,48,49的测定相比,尾部结构域的存在可以减缓细丝滑动。然而,仍然可以观察到滑行和表面运动。肌球蛋白活性的一个更明显的证明是活跃的肌动蛋白丝断裂,可以观察到较长的肌动蛋白丝被分解成更小的片段,然后向多个方向滑去。这是由于表面上高浓度的有源马达而发生的,已被多个实验室观察到,并且如果没有存在42,50,51,52,53,54的有源肌球蛋白马达,就不会发生。此外,这里介绍的束测定有助于缓解主要与滑动丝测定相关的运动性问题,例如玻璃盖玻片上的各种电机结合方向,因为束测定涉及酪蛋白阻断玻璃表面,以便电机结合在束内47,55,56。
第一步是将罗丹明肌动蛋白丝作为底部或模板丝添加到流通池中的聚-L-赖氨酸包被的盖玻片中。聚-L-赖氨酸用于促进肌动蛋白结合,因为聚赖氨酸带正电荷,而肌动蛋白带负电荷,并且已用于以前的细胞骨架体外测定制剂61,62,63。在束形成之前,将不同的肌动蛋白稀释液添加到流通池中以优化肌动蛋白浓度。在这种情况下,从库存中取出600倍是最佳稀释度,可产生足够数量的模板细丝用于束的形成,但具有足够的间距,以便束是个性化的。使用APB缓冲液进行稀释。添加罗丹明肌动蛋白后加入一层酪蛋白以阻断表面并避免非特异性结合。将流通池孵育30分钟,并在用缓冲液孵育后洗涤以洗出任何未结合的肌动蛋白丝。最后,将肌球蛋白、488/生物素肌动蛋白和链霉亲和素包被的磁珠组合添加到流通池中,以促进肌动蛋白-肌球蛋白束的形成。磁珠浓度应足以结合表面结合的束,并且有足够的悬浮液以促进校准。然而,磁珠浓度过高会导致捕获实验过程中的困难,因为相邻的磁珠落入激光陷阱并破坏测量。肌球蛋白马达在注射到载玻片之前被添加到组合中,这样肌球蛋白马达就不会与货物或顶部生物素化的肌动蛋白丝预先聚集,从而将底部罗丹明结合以捆绑生物素化的肌动蛋白丝。
NT2 光学捕获系统是一种商用光学捕获器,具有明场、微分干涉对比 (DIC) 和落射荧光成像模式。它与蔡司AxioObserver 3倒置显微镜配合使用,具有100x/NA 1.46和63x/NA 1.0水浸陷捕和检测物镜。该系统配备了一个激光陷阱的点击和拖动捕获功能,可以在以前列出的任何模式成像时使用。通过使用荧光成像检测和确认形成的束。具有具有适当滤光片立方体(GFP/FITC和TRITC/CY3)的白光源允许在灯丝成像之间快速切换。在使用光镊进行每次力测量之前,通过可视化不同激发波长下的自动对焦来验证共定位自动对焦。由于即使使用除氧试剂,灯丝也可以快速光漂白,因此建议研究人员在进行束实验之前优化可视化参数,例如强度和曝光时间。
采用光学捕获进行力测量,在ATP存在下使用链霉亲和素珠结合生物素化的货物肌动蛋白丝并激活肌球蛋白力生成作为力传感器。从捕获软件中提取通过光学捕获获得的位移和力与时间的关系数据进行分析。但是,商业诱捕软件还提供了可以利用的分析例程,或者用户可以对其他程序中的自定义算法进行编程,以可视化和分析诱捕数据。在定制的光学捕获系统上,用户可能使用激发激光器而不是带滤光片的白光源,这也是可以使用的。此外,如果发射光谱不重叠并导致渗漏,则可以更改荧光染料以适合用户可能拥有的现有设备。
我们注意到,所提供的测定是一种基线测定,用户可以根据他们在actomyosin集合力学领域的研究问题进一步定制。一般工作流程也可以应用于可能感兴趣的其他体外细胞骨架集合系统,例如形成有丝分裂纺锤体32,61,63,64,65,66的最小模型的微管束测定。修改可能包括但不限于更改适合用户现有设置的荧光团标记;改变肌球蛋白浓度、构建体或同种型;以及滴定缓冲液条件等方面。
进行此测定时可能存在潜在挑战。当形成肌动蛋白-肌球蛋白束时,肌动蛋白束内的肌球蛋白浓度在整个载玻片上可能不均匀。为了适应这种情况,将测量整个滑块上的多个管束,以确保对电机分布和力生成曲线进行正确采样。如果解释力数据需要了解束方向,那么了解束方向也具有挑战性。因此,应为每个捆绑包进行多次试验。还可以通过荧光凝胶索林或比光学捕获手柄更小的凝胶索林包被的珠子来结合肌动蛋白丝末端标记。荧光成像也可用于观察x和y分量力,以推断束取向。此外,由于肌球蛋白聚集状态受到缓冲液离子强度的高度影响,在KCl快速稀释时会形成粗丝,因此应适当监测缓冲盐浓度67,68。
以前使用其他体外方法(如滑动测定)的研究有助于确定肌球蛋白结构域的作用,并研究肌球蛋白与其他肌动蛋白结合蛋白之间的结构和相互作用。然而,这些方法有一个缺点,即将肌球蛋白结合到刚性表面上将限制肌球蛋白电机之间的协调潜力,从而限制机械传感反馈,以确定电机集合是否处于高占空比模式33,35,41,69。此外,使用单肌球蛋白运动网络进行光学捕获并不能清楚地了解肌球蛋白马达如何相互相互作用以及与肌动蛋白丝相互作用。这里开发的协议允许在兼容的分层肌动蛋白网络中研究肌球蛋白运动集合动力学。它还可以根据电机-灯丝集合特性(如浓度、亚型和缓冲环境等)进行定制,以便进行系统研究。所提出的协议是未来研究更复杂的肌动肌肽网络的平台,并保持了传统上用于单分子研究的光学捕获促进的位移和力产生测量的精度。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者没有利益冲突需要声明。
Acknowledgments
这项工作得到了密西西比大学研究生会研究奖学金(OA),密西西比大学Sally McDonnell-Barksdale荣誉学院(JCW,JER),密西西比州太空资助联盟(资助号NNX15AH78H)(JCW,DNR)和美国心脏协会(资助号848586(DNR)的部分支持。
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
