The following SOP provides a step-by-step description of the protocol used to resurrect Daphnia magna dormant eggs, including a detailed description of sampling, isolation of ephippia from the sediment, and establishment of clonal cultures (Figure 1).

Figure 1: Step-by-step guide to resurrection of Daphnia magna. Sediment from a natural freshwater habitat (A) is sampled with a piston corer (B). The sediment core (C) is sliced in incremental layers of 1 or 0.5 cm (D). Each layer of sediment is stored in a sample zip lock bag (E) in dark and cold conditions (4 °C). Each layer of sediment is weighed and sieved using geological sieves (1 mm and 125 µm mesh sizes, F). White background trays are used to isolate Daphnia magna ephippia (G). Decapsulated dormant eggs (H) are transferred to Petri dishes and exposed to light and temperature stimuli to induce hatching. Hatchlings are transferred to separate jars (I) to establish isoclonal lines. Please click here to view a larger version of this figure.
1. Sampling of Sediment Cores
- Sample sediment from lakes or ponds using a piston corer. This protocol used Big Ben27, a core tube of approximately 1.5 m in length with an internal tube diameter of 14 cm. Big Ben consists of a piston on a rope and a corer head, to which rods are attached to drive the tube into the sediment. A core catcher aids the support of the core tube when full of sediment. To extrude the sediment, a framework keeps the core tube upright and stationary, and a modified bottle jack is used to push the piston upwards during the extrusion process (Supplementary Video 1).
- For shallow ponds of less than 1 m in depth, use a plexiglass gravity corer of no more than 6 cm of diameter manually pushed into the sediment.
- For deep lakes (>6 m of depth), use Livingston piston corers28 or single-drive Griffith sediment corers with the aids of an anchored pontoon boat. The Livingstone-type drive rod piston corer can be used in water up to about 30 m deep to collect successive one-meter drives of soft to consolidated lake sediment. The single-drive Griffith corer consists of a simple but robust core head that connects standard polycarbonate tubes to Livingstone drive rods. The corers are pushed into the sediment with the rods, and a piston provides the suction needed for the recovery of sediment (Figure 2).
- For retrieving continuous, undisturbed cores, use vibracoring. These corers work on a variety of water depths and can retrieve core samples of different lengths, depending on sediment lithology. Low amplitude vibration that is transferred from the vibracore head down through the attached barrel or core tube liquefies sediments, enabling the core barrel attached to the vibracore unit to penetrate into the liquefied sediments. Some vibracorers are small, lightweight, and portable, others are large heavy units that can only be deployed from large vessels. The choice of corers depends on the lithology of sediment.
- Slice the core horizontally in incremental layers of 1 cm or less using a flat metal surface (Supplementary Video 1). Sediment corers like the one used here are designed to reduce hydrostatic pressures at extrusion, reducing disturbance of the sediment layers. When other corers are used, the outer rind of each sediment layer may be removed with a cookie-cutter sort of blade to limit contamination among layers.
- Collect each sediment layer in a separate sampling bag (Supplementary Video 1), and store in dark and cold (4 °C) conditions.
- Collect a minimum of 5 g of sediment from all layers for radiometric dating. As radiometric dating is an established protocol for a detailed description of the dating assay, refer to existing publications12,13.

Figure 2: Cartoon of the piston coring procedure. Piston corer, a hollow tube with an internal sliding seal (the piston) that produces a weak vacuum. When the piston touches the sediment-water interface, the weight pushes the core barrel into the sediment and the vacuum causes the sediment being cored to enter and move up the tube without disturbing the sediment layers. Please click here to view a larger version of this figure.
2. Sieving of Sediment Layers
- Using a precision scale, weigh each sediment layer for future reference. Use the surface area and weight to calculate the species density in the lake.
- Sieve each sediment layer using two geological sieves piled on top of each other. The first sieve has a mesh size of 1 mm and separates clay, large invertebrates and particulate matter, e.g. seeds, plants, and insects, from the remainder of the sediment. The second sieve has a mesh of 125 µm and separates D. manga ephippia and small particulate from the remainder of the sediment (Supplementary Video 2).
- Transfer small aliquots of the sediment fraction collected onto the 125 µm mesh sieve to a white background tray. Depending on the type of sediment, smaller or larger aliquots of sediment may be transferred at each time.
- Add small volumes (up to 200 mL) of medium to the sampling white tray to re-suspend the transferred sediment fraction and facilitate eye spotting of ephippia (Figure 3). The medium used to resuspend the sediment can be dechlorinated tap water, borehole water, COMBO29, or ADaM medium (Aachener Daphnien)30. Hereafter, the term 'medium' will be used to refer to either or all the listed solutions.

Figure 3: Daphnia magna ephippium. Dormant Daphnia magna eggs immediately after decapsulation. The ephippium (A), the inner egg membrane (B), and the dormant eggs (C) are shown. Scale bar = 500 µm.
3. Decapsulation of Ephippia and Hatching
- With a disposable Pasteur pipette or using microdissection forceps, transfer individual ephippia to Petri dishes filled with 10 mL of medium. Use at least a Petri dish per layer of sediment.
- Under a stereo microscope, decapsulate each ephippia using microdissection forceps by forcing open the chitin case (Supplementary Video 3). Remove the resting egg inner membrane delicately, paying attention to not disrupt the eggs, and transfer them to the Petri dish filled with medium using a Pasteur pipette. Decapsulation increases hatching success in D. magna; however, it may not be required or it may be challenging for other Daphnia species producing smaller ephippia.
- Expose the decapsulated eggs to a full spectrum long day photoperiod light (16:8 light:dark) and high temperature (20 ± 1 °C) to induce hatching in a controlled temperature device (incubator) or room. Hatching occurs between 48 h and several weeks (up to four; Supplementary Video 4). In absence of decapsulation, directly expose the ephippia to hatching stimuli (long day photoperiod light (16:8 light:dark) and high temperature (20 ± 1 °C)).
4. Establishing Isoclonal Lines of Daphnia magna
- Establish isoclonal lines from single hatchlings by transferring individual D. magna from step 3.3 to separate jars filled with 200 mL of medium using a disposable Pasteur pipette. Each individual is genetically distinct, being the result of sexual recombination.
- Maintain isoclonal lines indefinitely in stock conditions consisting of 10 ± 1 °C, 16:8 light:dark regime, fed weekly with 0.2 mg C/L of Chlorella vulgaris or other green algae (e.g., Scenedesmus obliquus). Renew the medium every third week. Stock conditions may change with temperature, feeding regimes, and species.
5. Key Studies
NOTE: A description of two key studies is provided in which resurrected D.magna (sub)populations from the sedimentary archive of Lake Ring (Denmark) are used to assess the evolutionary response to warming. Three (sub)populations were resurrected from the following time periods: 1960-1970, 1970-1985, and >1999. D. magna hatching success from the sedimentary archive ranged between 11 and 58% (Figure 4). From the hatchlings obtained from each timeperiod, a random subset was chosen for the two key studies described here. These studies were designed to assess whether (sub)populations resurrected from different time periods along the temperature gradient showed differences in fitness-linked life history traits (5.1), and whether they had different competitive abilities (5.2) after exposure to warming.

Figure 4: Hatching success in a sedimentary archive sampled from Lake Ring. The proportion of successful hatchlings along the sedimentary archive of Lake Ring used in the key studies. Please click here to view a larger version of this figure.
- Common garden experiments
- Transfer ten hatchlings per (sub)population from stock cultures to common garden conditions: 16 °C; long photoperiod (16:8 light:dark regime); feed daily with 0.8 mg C/L of Chlorella vulgaris, and renew medium every second day. Maintain the hatchlings in common garden conditions for at least two generations (ca. 45 days). Common garden conditions reduce interference from the maternal effect and synchronize reproduction among hatchlings.
- Upon release of the second brood into the brood chamber, transfer adult females from the second generation to 500 mL jars filled with medium until they release the second brood of juveniles.
- Randomly transfer individual juveniles of 24–48 h, born from the second brood of the second generation, to 100 mL jars filled with medium and exposed to the following experimental conditions: 18 °C (current temperature in the lake) and 24 °C (warming, temperature forecasted by the IPCC 26 for the upcoming 100 years), 16:8 light:dark regime, and feed daily 0.8 mg C/L of Chlorellavulgaris.
- On each experimental animal, measure the size at maturity with a stereomicroscope as the distance between the head and the base of the tail spine (Figure 5). Photograph each animal and subsequently analyze its size using a suitable image software.
- Measure the age at maturity: the day in which eggs are observed for the first time in the brood chamber.
- Measure the mortality: the number of extinct individuals during the experiment.
- Measure the fecundity: the total number of offspring released in first and second clonal reproduction.
- Measure the population growth rate, estimated using the Euler's equation (1):
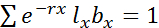 (1)
(1)
Where lx is the proportion of survival at age x, bx is the number of neonates produced per surviving individual at age x, and r is the intrinsic rate of natural increase. - Perform statistical analysis using commercially available software. Here, use R31 to plot reaction norms (phenotypic expression of a single genotype across environments) for each life history trait, using the ggplot2 package. Calculate mortality rates per population via survival analysis (R package rms; https://cran.r-project.org/web/packages/rms/rms.pdf). Finally, perform an analysis of variance (ANOVA, Table 2) in R31 to assess whether the effect of temperature on traits can be explained by evolution (differences among populations), plasticity (response to treatment), or evolution of plasticity (population x treatment).
- Competition experiment
- Transfer ten hatchlings per population from stock cultures to common garden conditions: 20 °C, long photoperiod (16:8 light:dark regime), feed daily with 0.8 mg C/L of Chlorella vulgaris, and renew medium every second day. Maintain common garden conditions for at least two generations (ca. 45 days) to reduce interference from maternal effects.
- Randomly assign five juveniles of 24 to 48 h from the second brood of the second generations of each hatchling to experimental mesocosms (20 L plastic aquaria filled with medium), in triplicates, at a density of 10 animals/L.
- Expose the mesocosms to 24 °C, 16:8 L:D regime, and feed daily with 0.8 mg C/L of Chlorellavulgaris in a controlled temperature chamber or incubator for a minimum of four weeks (>3 clonal generations).
- Cull each mesocosm weekly, refreshing 10% of the medium to simulate a population dynamic that Daphnia may encounter in the natural environment. Impose culling by removing a known volume of medium and animals from each aquarium (1.2 L in this case), and by replacing the culled volume with fresh medium. Start the culling regime after the experimental animals reach maturity (day 10).
- At the end of the fourth week, sample 32 animals from each mesocosm to assess shifts in genotypic composition as compared to the initial inoculum.
- Place individual Daphnia in microcentrifuge tubes and remove excess liquid with the help of a Pasteur pipette.
- Flash freeze individual tubes in liquid nitrogen and store at -80 °C.
- Extract genomic DNA from single individuals using available protocols and following the manufacturer's instructions.
- Amplify the extracted DNA using a number of genetic markers sufficient to provide a unique multilocus genotype per hatchling. Here, 8 microsatellites arranged in a single multiplex (M01, Table 1) were used following established protocols32,33.
- Genotype amplified fragments on a fragment analyzer.
- Conduct fragment analysis with a commercial or freely available software using a suitable size standard.
- Assess genotypic composition at the end of the experiment by genotyping the 32 individuals with a set of microsatellite loci as described33, and calculating the frequency of each genotype at the end of the experiment as compared to the initial inoculum.

Figure 5: Adult female Daphnia magna. Adult female Daphnia magna with parthenogenetic eggs in the brood chamber. The distance between the head and the base of the tail spine is used to measure size of the animal. The red lines indicate the size measurements. Scale bar = 500 µm.
Long-term empirical data are critical to the understanding of evolutionary dynamics and persistence of natural populations. Such data are generally challenging to obtain because of logistic difficulties associated with accessing temporal samples and the requirement of committing long-term to data collection. In the two key studies presented here, empirical evidence of the response to temperature of a central zooplankter in freshwater ecosystems is provided over evolutionary times. This is enabled by the use of layered dormant egg banks that provide the opportunity to study the response of historical populations and their modern descendants to environmental stress in common experimental settings.
Common garden experiment
The common garden experiment showed that all life history traits responded to temperature (Figure 6 and Figure 7). The ANOVA analysis revealed that all (sub)populations respond to temperature via plasticity (Table 2), except for mortality, which is unresponsive. Evidence of evolutionary changes (differences among (sub)populations) was observed only in population growth rate (Table 2), which significantly increased in two of the three (sub)populations at 24 °C (Figure 6).

Figure 6: Common garden experiment. Reaction norms for life history traits (fecundity, size, and age at maturity) and population growth rate (r) are shown for each (sub)population under temperature warming (24 °C) as compared to the common garden and current temperature regime (18 °C). The population growth rate 'r' is calculated using the Eulerian equation (1). Confidence intervals are shown. (Sub)populations are color coded: (i) blue: 1960-1970; (ii) green: 1970–1985; (iii) red: >1999. Please click here to view a larger version of this figure.

Figure 7: Mortality. Mortality rates per (sub)population (1960-1970; 1970-1985; >1999) are shown under warming (24 °C) as compared to modern temperature regimes (18 °C). Please click here to view a larger version of this figure.
Mesocosm experiment
After four weeks of selection, represented by warming at 24 °C, the frequency of the three (sub)populations did not change significantly (χ2 = 0.55, P = 0.76) as compared to the initial inoculum (Figure 8). Among the 30 genotypes inoculated in the mesocosm experiment, the majority was identified after four weeks of selection (Figure 9). Specifically, 70% of the inoculated genotypes were recovered, compatible with Poissonian expectation of recovering at least one representative of each genotype in a sample of 32 individuals.

Figure 8: Competition experiment – population frequency. Population-averaged median and quartiles (25th and 75th), is shown for the three (sub)populations of D. magna after four weeks of selection in mesocosm competition experiments (24 °C), as compared to an initial equal frequency (at the start). (Sub)populations are color coded as shown in Figure 6. Please click here to view a larger version of this figure.

Figure 9: Competition experiment – genotype frequency. Genotype frequencies — averaged median and quartiles (25th and 75th), are shown after four weeks of exposure to warming (24 °C) as compared to an initial equal frequency of genotypes (dotted line). Names on the x-axis are the inoculated genotypes ID, grouped per (sub)population (blue, 1960-1970; green, 1970-1985; red, >1999). Please click here to view a larger version of this figure.
| Locus | AN | Size range (bp) | Primers (5’-3’) | Dye label | Repeat motif | Tm | |
| B008 | HQ234154 | 150–170 | F: TGGGATCACAACGTTACACAA | VIC | (TC)9 | 56 | |
| R: GCTGCTCGAGTCCTGAAATC | |||||||
| B030 | HQ234160 | 154–172 | F: CCAGCACACAAAGACGAA | PET | (GA)11 | 56 | |
| R: ACCATTTCTCTCCCCCAACT | |||||||
| B045 | HQ234168 | 118–126 | F: GCTCATCATCCCTCTGCTTC | NED | (TG)8 | 56 | |
| R: ATAGTTTCAGCAACGCGTCA | |||||||
| B050 | HQ234170 | 234–248 | F: TTTCAAAAATCGCTCCCATC | 6FAM | (GAA)6 | 56 | |
| R: TATGGCGTGGAATGTTTCAG | |||||||
| B064 | HQ234172 | 135–151 | F: CTCCTTAGCAACCGAATCCA | 6FAM | (TC)8 | 56 | |
| R: CAAACGCGTTCGATTAAAGA | |||||||
| B074 | HQ234174 | 196–204 | F: TCTTTCAGCGCACAATGAAT | NED | (GT)9 | 56 | |
| R: TGTGTTCCTTGTCAACTGTCG | |||||||
| B096 | HQ234181 | 234–240 | F: GGATCTGGCAGGAAGTGGTA | VIC | (AC)15 | 56 | |
| R: TTGAACCACGTCGAGGATTT | |||||||
| B107 | HQ234184 | 250–274 | F: GGGGTGAAGCATCAAAGAAA | PET | (CT)8 | 56 | |
| R: TGTGACCAGGATAAGAGAAGAGG | |||||||
Table 1: Microsatellite multiplex. The NCBI Accession Number (AN), the multiplex information, the PCR primer sequences, the PCR size range, the repeat motif, the dye used to label the forward primer, and the annealing temperature (Tm) are shown.
| Pop Growth rate (r) | Df | F | P | |
| Evolution (Pop) | 2 | 30.309 | <0.001 | |
| Plasticity (Temp) | 1 | 531.546 | <0.001 | |
| Evol. Plasticity (Pop x Temp) | 2 | 65.137 | <0.001 | |
| Mortality | Df | F | P | |
| Evolution (Pop) | 2 | 2.234 | 0.1162 | |
| Plasticity (Temp) | 1 | 2.679 | 0.1071 | |
| Evol. Plasticity (Pop x Temp) | 2 | 1.8657 | 0.164 | |
| Fecundity | Df | F | P | |
| Evolution (Pop) | 2 | 1.8852 | 0.1633 | |
| Plasticity (Temp) | 1 | 6.8934 | 0.0117 | |
| Evol. Plasticity (Pop x Temp) | 2 | 1.6511 | 0.203 | |
| Size at maturity | Df | F | P | |
| Evolution (Pop) | 2 | 0.211 | 0.8106 | |
| Plasticity (Temp) | 1 | 11.1361 | 0.0017 | |
| Evol. Plasticity (Pop x Temp) | 2 | 0.6586 | 0.5225 | |
| Age at maturity | Df | F | P | |
| Evolution (Pop) | 2 | 0.7811 | 0.4637 | |
| Plasticity (Temp) | 1 | 8.0764 | 0.0066 | |
| Evol. Plasticity (Pop x Temp) | 2 | 0.088 | 0.9159 | |
Table 2: Analysis of variance(ANOVA). Analysis of variance testing whether changes in life history traits and population growth rate of the resurrected (sub)populations exposed to warming are explained by evolutionary adaptation (populations), plasticity (temperature treatment), and their interaction term (evolution of plasticity). Significant p-values (p<0.05) are shown in bold.
Supplementary Video 1: Sampling of sediment cores. The use of a Big Ben corer is shown. Big Ben is a core tube of approximately 1.5 m in length with an internal tube diameter of 14 cm. It consists of a piston on a rope and a corer head, to which rods are attached to drive the tube into the sediment. A core catcher is used to support the core tube that is deployed from a small vessel. The piston is pushed down into the sediment by gravitational pressure. A framework is used to support the core tube during the extrusion process carried out using a modified bottle jack that pushes the piston upwards. Each sediment layer is collected on a flat metal surface and transferred to transparent sampling bags for long term storage [dark and cold (4 °C) conditions]. Please click here to download this file.
Supplementary Video 2: Sediment sieving. The equipment required for sieving sediment is a precision scale, white sampling trays and geological sieves. From each sediment layer, at least 5 g are retained for radiometric dating. The remainder of the sediment is used to isolate ephippia. The sediment is sieved through two geological sieves, one with 1 mm and a second with 125 µm mesh size, piled on top of each other. Medium is poured on the 1 mm mesh sieve to separate clay, large invertebrates, and particulate matter. Medium poured on the second sieve with 125 µm mesh separates D. magna ephippia and small particulate matter. Aliquots of sediment are then transferred to a white sampling tray. D. magna ephippia are spotted by eye in the white background tray. Ephippia from each layer are collected in separate Petri dishes. Please click here to download this file.
Supplementary Video 3: Decapsulation. Under a stereomicroscope, D. magna ephippia are opened with microdissection forceps by applying pressure on the spine of the chitin case. The inner egg membrane is delicately removed and resting eggs gently transferred with a Pasteur pipette to a Petri dish containing 10 mL of medium. Please click here to download this file.
Supplementary Video 4: Hatching. After exposure to a long photoperiod and 20 °C, embryo development resumes between 48 h and few weeks. When development is complete, the embryos break free from the egg shell and freely swim in the medium. Please click here to download this file.
