The generation of transgenic flies expressing human Tar-Binding Protein of 43 kDa mutant (TDP-43M337V) is represented by the schematic (Figure 1A). This demonstrates the application of the binary Gal4/UAS system in Drosophila27. The illustration depicts a hemithorax with six muscle fibers, A‒F going from the most dorsal fiber A to the most ventral F (Figure 1B)11,12. To assess synaptic integrity, NMJs were stained with HRP and Phalloidin (Figure 1C‒E). Motor neurons in TDP-43M337V mutants (Figure 1F) have little to no HRP staining by Day 21, while WT (Oregon-R) remains intact (Figure 1C). There are no visible differences in muscle staining (Figure 1D,G). The changes in gross morphology observed in TDP-43M337V mutants demonstrates how synaptic integrity can be implicated in a neurodegenerative disease model of amyotrophic lateral sclerosis (ALS) using the adult DLM model. In addition to structural staining, staining the DLM NMJs can also provide an assessment of synaptic integrity with presynaptic (Figure 2A‒R) and post synaptic (Figure 2S‒X) markers. Together, these results illustrate how this dissection protocol could be applied to studying DLM tissue in neurodegenerative diseases.
One key aspect of this dissection is the application of liquid nitrogen to flash freeze the tissue to make the bisection easier. The utility of the liquid nitrogen is demonstrated in WT flies with liquid nitrogen where muscle tissue has no damage or nicked fibers (Figure 3A‒C). Without liquid nitrogen, the tissue can be more difficult to dissect. For example, following this protocol and skipping the liquid nitrogen flash freezing step allows the tissue to be more susceptible to damage from the dissection tools such as damaged neurons (Figure 3D) or damaged muscle fibers (Figure 3E). The application of liquid nitrogen helps to prevent tissue damage that could occur when working with DLM tissue regardless of the genotype of the specimen (Figure 3C and 3F).
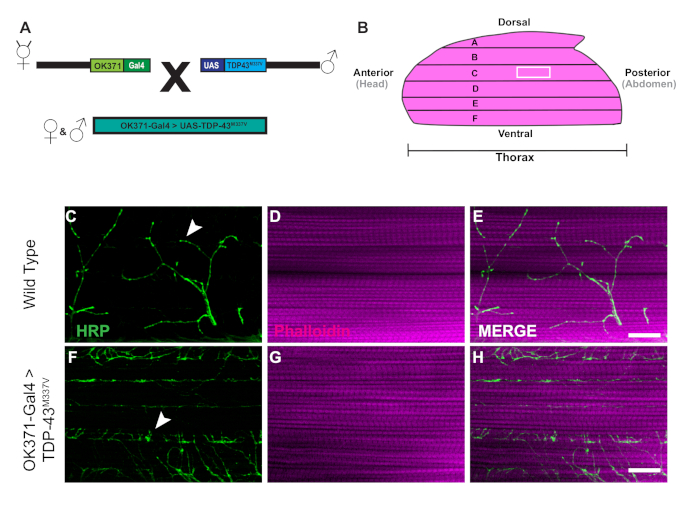
Figure 1: Progressive denervation of DLM synapses in a Drosophila model of ALS. (A) The generation of ALS transgenic flies expressing a human mutant form of Tar-Binding Protein of 43 kDa (TDP-43) are shown in the schematic. (B) The illustration depicts the shape and orientation of a hemithorax in an adult Drosophila. Using the protocol, we can observe the progressive loss of synaptic integrity of DLM NMJ synapses through structural staining of motor neurons with HRP (green) and muscle tissue with Phalloidin (magenta). Our model depicts the loss of synaptic integrity in an adult model of ALS through the generation of adult flies expressing a mutant from of human TDP-43M337V in motor neurons (Figure 1F‒H) in comparison to WT (Figure 1C‒E) flies in muscle fiber C. Arrows highlight examples of a WT synapse (Figure 1C) and an example of loss of synaptic integrity. Scale bar =20 µm at 63x magnification. Please click here to view a larger version of this figure.
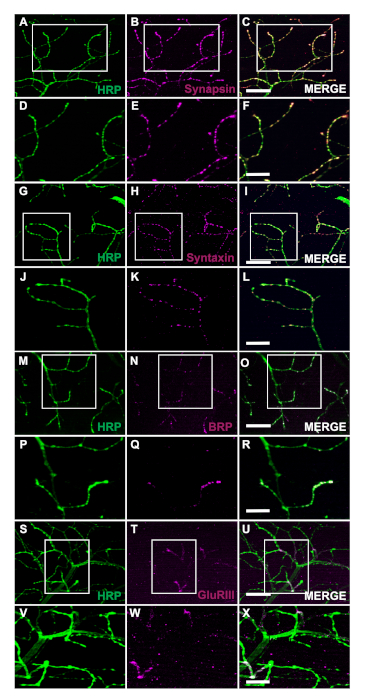
Figure 2: Assessing synaptic integrity using presynaptic markers at adult NMJs. Synaptic integrity can also be assessed using presynaptic and postsynaptic markers in WT flies that are 14 days old in muscle fiber C. The presynaptic markers Synapsin (B), Syntaxin (H), and Bruchpilot (BRP) (N) are co-stained with HRP (A, G, M). The staining depicts the localization of these markers to the presynaptic terminals (C, I, O). At higher magnification, the images illustrate the localization of Synapsin (E), Syntaxin (K), and BRP (Q) with HRP (D, J, and P) in more detail (Figure F, L, and R). We also show a postsynaptic marker Glutamate Receptor III (GluRIII) (T) co-stained with HRP (S). The co-staining demonstrates the utility of these markers (U). At higher magnification the representative images exemplify the localization (X) of GluRIII (W) and HRP (V) to the postsynaptic muscle tissue and the presynaptic terminals, respectively. Scale bar for panels A‒C, G-I, M‒O, S‒U represent 20 µm at 63x magnification. Scale bar for panels D‒F, 2J-2L, 2P-2R, and 2V-2X represent 10 µm at 63x magnification. Please click here to view a larger version of this figure.
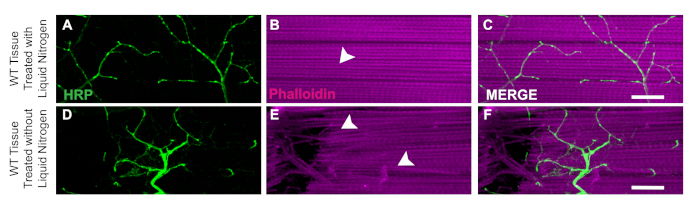
Figure 3: Utility of liquid nitrogen for DLM dissections. To demonstrate the utility of liquid nitrogen for the DLM dissections, we show a comparison of day 21 WT flies with and without liquid nitrogen from muscle fiber C. With liquid nitrogen, Phalloidin (B) remains intact and does not compromise the HRP staining (A, C). Without liquid nitrogen, muscle tissue becomes stringy and difficult to bisect (E) and HRP staining (D, F) becomes compromised due to technical error. White arrows show an area of no muscle damage in with liquid nitrogen (B) and damaged muscle tissue (E). Scale bar = 20 µm at 63x magnification. Please click here to view a larger version of this figure.
