

Summary
Kombinationen av kromatin immunoprecipitation och ultra-high-throughput sekvensering (chip-punkter) kan identifiera och kartlägga protein-DNA interaktioner i en viss vävnad eller cellinje. Beskrivs är hur man skapar en hög mall kvalitet chip för efterföljande sekvensering, med erfarenhet av transkriptionsfaktor TCF7L2 som ett exempel.
Abstract
Chip-sekvensering (Chip-seq) metoder ger direkt hel-genom täckning, där kombinationen av kromatin immunoprecipitation (chip) och massivt parallell sekvensering kan utnyttjas för att identifiera den repertoar av däggdjurs-DNA-sekvenser som är bundna av transkriptionsfaktorer in vivo. "Nästa generations" genomsekvenseringsprojekt teknik ger 1-2 storleksordningar ökning av mängden av sekvensen som kan vara kostnadseffektivt genererade över äldre teknik vilket möjliggör Chip-Seq metoder för att direkt ge hel-genom täckning för effektiv profilering av däggdjur protein-DNA-interaktioner.
För framgångsrika Chip-Seq metoder, måste man generera hög kvalitet chip DNA-mall för att få bästa sekvensering resultaten. Beskrivningen är uppbyggd kring erfarenheterna med proteinet produkten av genen starkast inblandad i patogenesen för typ 2-diabetes, nämligen transkriptionsfaktor transkriptionsfaktor 7-liknande 2 (TCF7L2). Denna faktor har också varit inblandad i olika cancerformer.
Beskrivs är hur man skapar hög kvalitet mall Chip DNA härlett från kolorektal karcinomcellinjen, HCT116, i syfte att bygga upp en högupplöst karta genom sekvensering för att bestämma de gener som är bundna av TCF7L2, vilket ger ytterligare inblick i dess nyckelroll i patogenesen av komplexa drag.
Introduction
Under många år har det funnits ett uppdämt behov av att identifiera uppsättningen av gener bundna och regleras av ett visst protein genom bred, i synnerhet de i transkriptionsfaktor klassen.
Odom et al. 1 används kromatin immunoprecipitation (chip) i kombination med promotor microarrays för att systematiskt kartlägga generna utnyttjas av pre-specificerade transkriptionsregulatorer i human lever och holmar bukspottkörtel. Därefter utvecklade Johnson et al. 2 en storskalig analys kromatin immunoprecipitation baserad på direkta ultra high-throughput DNA-sekvensering (chip-punkter) i syfte att på ett heltäckande kartlägga protein-DNA interaktioner över hela däggdjursgenom. Som ett testfall, kartlagt de in vivo bindningen av neuron-begränsande ljuddämpare faktor (NRSF) till 1946 platser i det mänskliga genomet. De data som visas skarp upplösning av bindande ställning (+ 50 baspar), vilket underlättade både itröst av motiv och identifieringen av NRSF-bindande motiv. Dessa chip-Seq uppgifter hade också hög sensitivitet och specificitet och statistisk konfidens (P <10 -4), egenskaper som är viktiga för härledning nya kandidatländer interaktioner.
Robertson et al. 3 används också ChIP-punkter för att kartlägga STAT1 mål i interferon-γ (IFN-γ)-stimulerade och ostimulerade humana HeLa S3-celler in vivo. Av Chip-punkter, med 15,1 och 12,9 miljoner unikt kartlagt sekvens läser, och en uppskattad falsk upptäckten av mindre än 0,001, identifierade de 41.582 och 11.004 förmodade STAT1-bindande regionerna i stimulerade och ostimulerade celler, respektive. Av de 34 loci som man vet innehåller STAT1 interferon-responsiv bindningsställen 4-8, fann ChIP-seq 24 (71%). Chip-Seq mål berikades i sekvenser som liknar kända Stat1 bindningsmotiv. Jämförelser med två befintliga Chip-PCR datamängder föreslåsatt chip-seq känslighet var mellan 70% och 92% och specificitet var minst 95%. Dessutom var det tydligt att Chip-seq erbjuder både låg analytisk komplexitet och känslighet som ökar med sekvensering djup.
Som sådan, "next-generation" genomsekvenseringsprojekt teknik ger 1-2 storleksordningar ökning av mängden av sekvensen som kan vara kostnadseffektivt genererade över äldre teknik 9. Chip-Seq metoder därför direkt ge hel-genom täckning för effektiv profilering av däggdjurs protein-DNA interaktioner 3.
År 2006 var en stark sammanslutning av varianter i transkriptionsfaktor 7-liknande 2 (TCF7L2) genen med typ 2-diabetes upptäcktes 10. Andra forskare har redan självständigt replikeras detta fynd i olika etniciteter och, intressant nog, från den första genomet hela föreningen studier av typ 2-diabetes som publiceras i Nature 11,12 Science 13-15 och annorstädes 16,17, var den starkaste föreningen verkligen med TCF7L2, detta är nu anses vara den mest betydelsefulla genetiska fynd i typ 2-diabetes som hittills 18-20. Dessutom har TCF7L2 länkats till cancerrisken 21,22, ja, blev detta sammanhang mer uppenbart när 8q24 locus avslöjades av genomet hela föreningen studier av en rad cancerformer, inklusive kolorektal cancer, visade sig bero på en extrem uppströms TCF7L2-bindande elementet driver transkriptionen av MYC 23,24. Som sådan, det finns ett stort intresse för att fastställa de nedströms gener som regleras av denna nyckel transkriptionsfaktor.
Baserat på erfarenhet med TCF7L2 som ett exempel på den metod, beskriver detta papper hur man skapar hög kvalitet mall Chip DNA. ChIP utfördes i det kolorektala karcinomcellinjen, HCT116, för efterföljande sekvensering för att bygga en hög-resolmepannor karta av generna som är bundna av TCF7L2 25 i en strävan att ge ytterligare inblick i dess nyckelroll i patogenesen av komplexa egenskaper.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Ett. Cross-link Chromatin
- Odla celler i 100x20mm rätter cellkultur. Mängden celler kan variera från 1 till 10 miljoner celler per skål beroende på celltyp. Cirka 2 miljoner celler är tillräckligt för en immunoprecipitation.
- Cross-link celler i 1% formaldehyd under 10 min vid rumstemperatur med tillfällig skakning.
- Quench tvärbindning genom tillsats av en slutlig koncentration av 125 mM Glycin och inkubera i 5 min vid rumstemperatur.
- Tvätta cellerna med 1X Fosfatbuffrad saltlösning (PBS) två gånger, dekantera PBS och tillsätt sedan 0,2 ml PBS.
- Harvest celler med en plast cellskrapa i ett mikrocentrifugrör.
- Centrifugera ner cellerna vid 2000 rpm under 5 min vid 4 ° C.
- Aspirera supematanten. Resuspendera cellerna i SDS lyseringsbuffert (1% SDS, 10 mM EDTA, 50 mM Tris-HCl pH 8,1) för helcellslysat eller hålla dem som en pellet för nukleär extraktion.
- Celler kan sparas vid -80 ° C eller man kan gå vidare immediatEly med ultraljud.
2. Förbered Kärnor (Fortsätt till steg 3,5 för Whole Cell Lysate)
- Supplement Buffer Cell Lysis (5 mM PIPES pH 8,0, 85 mM KCl, 0,5% NP-40) med 1X Proteinase Inhibitor varje experiment.
- Resuspendera tinade cell pelleten i cirka 10 gånger pelleten volym med Cell lyseringsbuffert.
- Dounce-homogenisering 10 gånger med mortelstöt inkubera sedan på is i 10 min.
- Centrifugera provet vid 4000 rpm i 5 minuter vid 4 ° C, kassera supernatanten och spara nukleära pellet.
3. Sonication *
- Värm upp SDS lyseringsbuffert och komplettera mängd buffert för att användas med proteinasinhibitor.
- Omsuspendera nukleär pellet i SDS lysbuffert (ca 0,5 ml buffert per 1-10.000.000 celler)
- Inkubera på is under 10 min.
- Lägg 0,5 ml alikvoter av prov till mikrocentrifugrör.
- Låt ligga på våt is med Misonix sonikator med 30 sekunder på och 45 sekunderav vid en amplitud inställning av två. Antal cykler för ideal fragment storlek kan bestämmas genom att först prova olika cykel nummer (ex. 2, 4, 8, 12, 16, och 20 eller fler cykler). Ett annat märke av sonikator kan användas, dock förutsättningarna varierar. Experiment med antalet cykler och mängden tid på och av måste genomföras för att fastställa de ideala förhållanden.
- Samla 20 pl av varje prov för att kontrollera sonication resultat och för att göra kvantifiering,. Resten av provet kan lagras vid -80 ° C.
- Späd 20 il prov genom att tillsätta 30 | il 0,1 X TE-buffert.
- Behandla provet med 1 pl RNas A vid 37 ° C under 1 timme och tillsätt sedan 1 | il proteinas K och inkubera vid 62 ° C under 2 timmar.
- Kör 20 | il av provet på en 2% agarosgel.
- Rena resterande mängd prov med QIAquick PCR Rening kit sedan kvantifiera använda NanoDrop spektrofotometer.
* För infödda CHIP, kan microccocal nukleasdigerering alternativt användas för att skjuva DNA.
4. Block agarospärlor *
- Om pärlorna är redan blockerad, gå vidare till steg 5.1.
- Använd protein A eller protein G agaros. För 5 immunoutfällningar (IP), använd 600 pl 50% pärla slurry (300 pl pärlpelleten)
- Att tvätta pärlorna, snurra av dem vid 800 rpm under 1 min vid 4 ° C och kasta bort supernatanten. Lägg något mer än 2 ml ChIP spädningsbuffert (0,01% SDS, 1,2 mM EDTA, 167 mM NaCl, 1,1% Triton X-100, 16,7 mM Tris-HCl pH 8,1) och blanda genom att långsamt vända röret 10X. Centrifugera ner igen vid 800 rpm under 1 min vid 4 ° C och kasta supernatanten. Upprepa tvätta två gånger till.
- Blockera pärlor genom att rotera vid 4 ° C över natten i blockerande lösningen. Se tabell 1 för receptet av den blockerande lösningen.
Fem. Pre-klar Chromatin
- Thaw sonikerades kromatin på is.
- Spinn ner vid 12.000 rpm feller 10 min vid 4 ° C sedan lägga på is direkt för att ta bort SDS (vit pellet).
- Samla supernatanten, kassera pellet, och kombinera prover vid behov.
- Ta ut de belopp som behövs för experimentet baserat på beräkningar (1-10 ug av kromatin per IP).
- Späd kromatin 10X chip Spädningsbuffert kompletterad med proteinasinhibitor.
- Tillsätt 100 ul blockerade pärlor per IP.
- Rotera vid 4 ° C under 1 timme.
6. Immunoutfällning
- Centrifugera ner prover vid 800 rpm under 1 min och överför supernatanten till ett nytt rör.
- Centrifugera ner supernatanten vid 800 rpm under 1 min och överföring till ett annat rent rör.
- Spara 20 ìl av supernatanten, för att tjäna som input kontroll, vid -20 ° C.
- Alikvotera kromatinet till antalet IP-adresser som skall göras vid experimentet.
- Lägg 2 ug antikropp per 1-10 ug av kromatin till varje prov.
- Inkubera över natten 4 ° C med rotation.
- Tillsätt 100 mikroliter av blockerade pärlor till varje IP prov.
- Inkubera under en timme vid 4 ° C med rotation.
- Pellet pärlorna genom att snurra ner vid 800 rpm i 1 minut och kassera så mycket av supernatanten som möjligt.
- Tvätta pärlor gång med låg salthalt immunkomplex tvättbuffert. Tillsätt 1 ml buffert till varje rör, rotera vid rumstemperatur under 5-8 min; spinn ner vid 800 rpm under 1 min och därefter kassera supernatanten. Upprepa tvätta en gång med High Salt immunkomplex tvättbuffert och LiCl immunkomplex tvättbuffert och två gånger med TE-buffert för totalt fem tvättar (tabell 2).
7. Eluering
- Tina insampel från föregående dag som skall bearbetas med eluanter.
- Gör färskt elueringsbuffert (tabell 3).
- Gör en mästare blandning av tillräckligt elueringsbuffert behövs för IP-adresser och input kontrollprover plus 1-2 extra prover.
- Tillsätt 100 | il elueringsbuffert till varje IP-prov och inkubera vid rumstemperatur under 15 min med rotation.
- Centrifugera ner vid 800 rpm i 1 minut och tillsätt supernatanten till ett nytt rör.
- Lägg till ytterligare 100 pl av Elueringsbuffert till varje rör av pärlor och inkubera vid rumstemperatur under 15 min med rotation.
- Vortex i 15 sek efter inkubation, spinn ner vid 5000 rpm under 1 min, sedan kombinera supernatanten med supernatanten från den första elueringen. (Se till att det inget finns kvar kulor i de överstående. Om du är osäker, spinn ner supernatanten igen vid 5000 rpm i 1 minut och samla in supernatanten i ett nytt rör.
- Lägg 180 pl elueringsbuffert till 20 pl prover ingående kontroll.
8. Omvänd Cross-link
- Till 200 pl eluanter och kontroller insatsvaror, tillsätt 8 pl 5 M NaCl.
- Slutna rör med parafilm och inkubera i vattenbad vid 65 ° C över natten.
9. DNA Rening
- Behandla varje prov med 1 pl RNas A under 1 timme vid 37 ° C.
- Lägg 4 μl av 0,5 M EDTA, 8 | il 1 M Tris-HCl, blanda, tillsätt sedan en pl av proteinas K till varje prov och inkubera vid 45 ° C under 2 timmar.
- Rena proverna med QIAquick PCR Rening kit. Proverna kan sparas vid -20 ° C och PCR-kontroll kan göras vid en senare tidpunkt.
* Alternativt kan ChIP-grade magnetiska kulor användas i stället för agaros för immunoprecipitation delen.
10. PCR check
- För PCR-kontroll, använda primrar för regioner som är kända att vara bunden av proteinet av intresse. Dessutom använder primers för icke-bindande regioner som negativa kontroller.
- Blanda reagensen för reaktionen. Späd Input provet vid 1:100 (tabell 3).
- Kör reaktionen. PCR-program:
Steg 1: 94 ° C 3 min
Steg 2: 94 ° C 20 sek
59 ° C 30 sek
72 ° C 30 sek
(Upprepa steg 2 under minst 30 cykles)
Steg 3: 72 ° C 2 minuter
- Kör prover på 1% agaros-gel.
- Berikning kan också bestämmas kvantitativt med realtids-PCR.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
När kromatin har sonikerades och har behandlats med RNas och proteinas bör de prov som körs på 2% agarosgel presentera ett utstryk med huvuddelen av DNA vid den önskade storleken. Om flera olika cykler testas, bör en gradvis minskning i storlek ses som antalet cykler ökar (Figur 2).
Efter genomgången immunoprecipitation delen av protokollet anrikningen kan antingen kontrolleras av PCR eller realtids-PCR. För PCR-prov som körs på en agarosgel det bör finnas band i Input och chip (med antikroppar för proteinet av intresse, vilket är TCF7L2 i detta fall) körfält prov och ingenting eller högst ett mycket svagt band (bakgrundsljud) i IgG (negativ) kontroll körfält för positiva bindande regionen. För den negativa bindande regionen bör det vara mycket svagt eller inget band för IgG-kontroll och körfält chip. Det bör finnas ett band i Input körfält (Figur 3).
Figur 4 visar samma prov som undersökts genom realtids-PCR. I likhet med den föregående figuren, bör det finnas en betydande faldig anrikning av de positiva bindningsregionen för chipsampeldata över IgG kontrollen. Dessutom bör det finnas mycket lite anrikning, om någon, sett i den negativa bindande regionen.

Figur 1. Flödesschema av chip process. Klicka här för att visa en större bild .
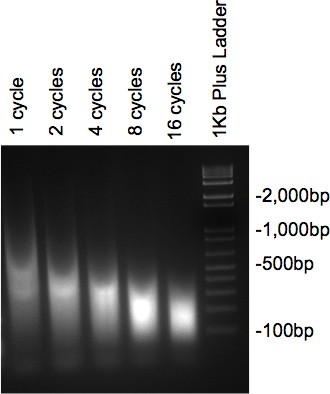
Figur 2. Gel kontrollera av DNA ultraljudsbehandling.

Figur 3. PCR Kontroll av chip.
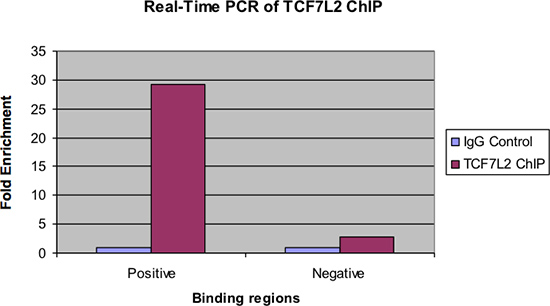
Figur 4. Realtids-PCR av TCF7L2 chip.
| Reagens | Volym |
| Bead pellet | 300 | il |
| BSA (50 mg / ml) | 30 | il |
| 100X proteinasinhibitor | 10 pl |
| ChIP Spädningsbuffert | 660 | il |
| Totalt | 1000 | il |
Tabell 1. Recept för blockering agaros.
| Buffert | Komponenter |
| Låg Salt immunkomplex tvättbuffert | 0,1% SDS 1% Triton X-100 2 mM EDTA 20 mM Tris-HCl pH 8,1 150 mM NaCl |
| Hög Salt immunkomplex tvättbuffert | 0,1% SDS 1% Triton X-100 2 mM EDTA 20 mM Tris-HCl pH 8,1 500 mM NaCl |
| LiCl immunkomplex Wash Buffer | 0,25 M LiCl 1% NP-40 1% deoxicholat 1 mM EDTA 10 mM Tris-HCl pH 8,1 |
| TE-buffert | 10 mM Tris-HCl pH 8,1 1 mM EDTA pH 8,0 |
Tabell 2. CHIP tvättbuffertar.
| Reagens | Volym |
| 10 pl | |
| 1 M NaHCO 3 | 20 | il |
| H2O | 170 | il |
Tabell 3. Elueringsbuffert för en IP.
| Reagens | 50 pl reaktion | 20 pl reaktion |
| Vatten | 27 | il | 10,8 | il |
| 5X PCR-reaktionsbuffert | 10 pl | 4 | il |
| MgCl2 | 4 | il | 1,6 | il |
| dNTP (10 mM) | 1 il | 0,4 | il |
| Primer mix (5 uM vardera) | 2 pl | 0,8 | il |
| Taq (Promega Hotstart) | 1 pl </ Td> | 0,4 | il |
| ChIP DNA | 5 | il | 2 pl |
Tabell 4. PCR-reaktionsvolymer.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Det är nu möjligt att utföra en genomet hela profilen av protein-DNA interaktioner association med Chip-punkter, som har mycket nyligen visats med andra transkriptionsfaktorer 2,3. Nyckeln till en lyckad sekvensering resultat är skapandet av en hög kvalitet kromatin immunoprecipitation DNA-mall.
När DNA-mallen har genererats och konstaterats vara tillräckligt berikas, kan man sedan ta det i biblioteket förberedelse för efterföljande sekvensering. Till exempel kan man använda sekvensering biblioteket protokollet tillhandahållet av säljaren, Illumina. Storleken urval av detta bibliotek kan utföras genom gelelektrofores och efterföljande excision och rening av DNA i det ungefär 200 - till 700-bp-intervallet. Minska storleken och minska storleken intervallet DNA som samlats in från gelrening syftar till att förbättra positionella upplösningen av chip-punkter. Genom berikande för mindre bitar av ingående DNA bunden till faktor interest, skulle man förvänta sig att platsen platsen kommer att få upplösningen. Hårdare storleksselektion förbättrar också storleken likformigheten av molekylära kolonier produceras på Illumina plattformen. Sådan kolonistorlek enhetlighet ökar också effektiva lästa numret erhållits. Kortare input DNA storlek producerar också mer robusta kolonier på Illumina plattformen, och detta kan innebära att kortare DNA-bitar i varje given insamplet distributionen kommer att vara representerade på ett effektivare sätt i den sista sekvensen produktion än är längre in bitar från samma fördelning.
De bioinformatiska metoder för "nästa generations" sekvensanalys fortsätter att utvecklas, med många leverantörer gör sin programvara med öppen källkod för vidare förädling. Man kan förändra läser denna kartan till unika genomiska platser i ett DNA-fragment överlappning profil. Väsentliga toppar kan identifieras genom threshholding profiler på en höjd som motsvarar en beräknad falsk upptäckt takt. Positionen specific frekvens matriser som härrör från detta arbete kan användas för att identifiera och lokalisera DNA bindningsställen hela det mänskliga genomet för en given faktor.
Men man måste vara försiktig när det gäller vilka faktorer man vill studera med Chip-punkter. Innan man börjar på en sådan studie bör man bedöma om en antikropp är tillgänglig på marknaden som är användbar i chippet inställning, som en dålig antikropp kan få mycket negativa konsekvenser för sina experimentella resultat. Dessutom bör man överväga om det finns skarva isoformer av proteinet som studeras, ja, är TCF7L2 känd för att ha många isoformer så vi var särskilt försiktig i valet av en antikropp som binds till aminosyror kontinuerligt förekommer i alla huvudsakliga isoformer av denna transkriptionsfaktor 25.
Sammanfattningsvis kan kombinationen av kromatin immunoprecipitation och ultra-high-throughput sekvensering (chip-seq) identifiera och kartlägga protein-DNA interaktioner i en viss vävnad eller cell linje. Vi har beskrivit hur man skapar en hög mall kvalitet chip för efterföljande sekvensering.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Författarna förklarar att de inte har några konkurrerande ekonomiska intressen.
Acknowledgments
Arbetet stöds av ett institut Development Award från Barnens Sjukhus av Philadelphia.
Materials
| Name | Company | Catalog Number | Comments |
| QIAquick PCR Purification Kit | Qiagen | 28104 | |
| EZ-ChIP Kit | Millipore | 17-371 | |
| GoTaq Hot Start Polymerase | Promega | M5001 | |
| Misonix Sonicator | Qsonica | XL-2000 | |
| NanoDrop 1000 Spectrophotometer | Thermo-Scientific | ||
| Positive control primer sequences (TCF7L2-1) Forward- 5'-TCGCCCTGTCAATAATCTCC-3' Reverse- 5'-GCTCACCTCCTGTATCTTCG-3' Negative control primer sequences (CTRL-1) Forward-5'-ATGTGGTGTGGCTGTGATGGGAAC-3' Reverse- 5'-CGAGCAATCGGTAAATAGGTCTGG-3' |
|||
References
- Odom, D. T., et al. Control of pancreas and liver gene expression by HNF transcription factors. Science. 303, 1378-1381 (2004).
- Johnson, D. S., Mortazavi, A., Myers, R. M., Wold, B. Genome-wide mapping of in vivo protein-DNA interactions. Science. 316, 1497-1502 (2007).
- Robertson, G., et al. Genome-wide profiles of STAT1 DNA association using chromatin immunoprecipitation and massively parallel sequencing. Nature Methods. 4, 651-657 (2007).
- Reich, N. C., Liu, L.
- Lodige, I., et al. Nuclear export determines the cytokine sensitivity of STAT transcription factors. The Journal of Biological Chemistry. 280, 43087-43099 (2005).
- Schroder, K., Sweet, M. J., Hume, D. A. Signal integration between IFNgamma and TLR signalling pathways in macrophages. Immunobiology. 211, 511-524 (2006).
- Vinkemeier, U. Getting the message across, STAT! Design principles of a molecular signaling circuit. The Journal of Cell Biology. 167, 197-201 (2004).
- Brierley, M. M., Fish, E. N.
- Bentley, D. R.
- Grant, S. F., et al. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nature Genetics. 38, 320-323 (2006).
- Sladek, R., et al. A genome-wide association study identifies novel risk loci for type 2 diabetes. Nature. 445, 881-885 (2007).
- Wellcome Trust Case Control Consortium. Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature. 447, 661-678 (2007).
- Saxena, R., et al. Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science. 316, 1331-1336 (2007).
- Zeggini, E., et al. Replication of genome-wide association signals in UK samples reveals risk loci for type 2 diabetes. Science. 316, 1336-1341 (2007).
- Scott, L. J., et al. A genome-wide association study of type 2 diabetes in Finns detects multiple susceptibility variants. Science. 316, 1341-1345 (2007).
- Steinthorsdottir, V., et al. A variant in CDKAL1 influences insulin response and risk of type 2 diabetes. Nature Genetics. 39, 770-775 (2007).
- Salonen, J. T., et al. Type 2 Diabetes Whole-Genome Association Study in Four Populations: The DiaGen Consortium. American Journal of Human Genetics. 81, 338-345 (2007).
- Zeggini, E., McCarthy, M. I. TCF7L2: the biggest story in diabetes genetics since HLA. Diabetologia. 50, 1-4 (2007).
- Weedon, M. N.
- Hattersley, A. T. Prime suspect: the TCF7L2 gene and type 2 diabetes risk. The Journal of Clinical Investigation. 117, 2077-2079 (2007).
- Yochum, G. S., et al. Serial analysis of chromatin occupancy identifies beta-catenin target genes in colorectal carcinoma cells. Proceedings of the National Academy of Sciences of the United States of America. 104, 3324-3329 (2007).
- Duval, A., Busson-Leconiat, M., Berger, R., Hamelin, R. Assignment of the TCF-4 gene (TCF7L2) to human chromosome band 10q25.3. Cytogenet. Cell Genet. 88, 264-265 (2000).
- Pomerantz, M. M., et al. The 8q24 cancer risk variant rs6983267 shows long-range interaction with MYC in colorectal cancer. Nature Genetics. 41, 882-884 (2009).
- Tuupanen, S., et al. The common colorectal cancer predisposition SNP rs6983267 at chromosome 8q24 confers potential to enhanced Wnt signaling. Nature Genetics. 41, 885-890 (2009).
- Zhao, J., Schug, J., Li, M., Kaestner, K. H., Grant, S. F. Disease-associated loci are significantly over-represented among genes bound by transcription factor 7-like 2 (TCF7L2) in vivo. Diabetologia. 53, 2340-2346 (2010).
- Benjamini, Y., Yekutieli, D. Quantitative trait Loci analysis using the false discovery rate. Genetics. 171, 783-790 (2005).
