

Summary
在这里,我们介绍简单的方法编制泡,封装的转录和翻译机制,监测蛋白质的生产。将得到的无细胞系统可以用来作为一个起点,从而建立越来越复杂的细胞模仿。
Abstract
从单个分子的分子系统随着利率的变化,越来越多的实验室已设法建立从下往上蜂窝模仿细胞生命的复杂性,更好地代表。迄今为止,也有一些可以采取,建立条块蜂窝模仿的路径,包括开采的油包水型乳状液,微流体装置,和囊泡。可用的选项中的每一个具有特定的优点和缺点。例如,油包水型乳剂给予较高的包封率,但没有模仿活细胞的通透性屏障。本文所述的方法的主要优点是,它们都容易和便宜的实施。转录 - 翻译机械是封装内的磷脂小泡通过一个过程,它利用常见的仪表,如离心式蒸发器和挤出机。荧光光谱反应监测。该协议s可以适于重组蛋白的表达,细胞模仿建设,探索细胞生命的最低要求,或遗传的电路组件的。
Introduction
无细胞, 在体外转录翻译反应和合成脂质囊泡代是什么新鲜事。然而,成蜂窝模拟相结合的两个显着更多challenging1-6 E。带或不带T7 RNA聚合酶的大肠杆菌细胞提取物可用于作为源转录-翻译机器7,8。细胞提取物受益于额外的细胞成分的存在下,可以方便蛋白的表达和折叠。或者,单独纯化的RNA和蛋白质分子的混 合, 即在PURE系统9,可以用于介导合成intravesicular蛋白4,10-14。纯系统可以完全定义蜂窝模仿,建设并不会受到细胞提取物中发现核酸酶活性。实际上,这意味着,需要少得多的DNA模板,从而促进低包封11的流程与16-18。
监测转录 - 翻译反应最简单的方法是测量基因编码的元素的荧光或发光。通常情况下,萤火虫荧光素酶19或GFP的使用,虽然在体外的反应,经常测定放射性标记。荧光检测还可以让囊泡20,21人群的监测,通过基于流式细胞仪的方法,从而提供一些洞察类似生物过程的随机性。这些监测方法已被用来定义一个小的设计规则和部件,从而建立从图书馆,包括荧光蛋白的集合, 与 i体外转录-翻译22,22表达的基因的组织的影响,西格玛因子16的活性,和转录终止子效率23。尽管如此,仍然有许多增加的能力, 在体外建立可预测,基因编码的设备,需要做的事情。
有许多方法可以使囊泡。最常用的方法取决于在玻璃表面上,然后再悬浮在水溶液中solution24薄的脂质膜的生成。如果该水溶液含有转录 - 翻译机器,例如,则形成囊泡的一小部分将包含蛋白质的生产所需的元件。然而,这样的方法包封为低,这意味着,只有一小部分的囊泡是活跃的。许多替代的方法,其特征在于高得多的封装EFF的iciency利用囊泡的油包水型乳状液的液滴转换。虽然这是可能的,这种方法将在未来是司空见惯,目前这些方法存在需要专门的设备和给囊泡有改变膜组合物的25。的油包水型囊泡的方法的一个显着优点是控制膜的层数(lamellarity)的电位。本文所述的方法是根据在薄的脂质膜YOMO实验室11所述的协议,有轻微的修改,包括一个额外的均质化步骤。这个方法很简单,价格便宜,而且提供了强大的囊泡转录翻译机器的封装非常适合。
Protocol
1。准备DNA模板
- 从一个标准的实验室菌株大肠杆菌中纯化质粒大肠杆菌 ,如大肠杆菌大肠杆菌 DH5a中或新商业套件蓝。可替换地,直链的PCR产物,可以类似地用市售的试剂盒纯化。洗脱DNA与H 2 O只。
- 苯酚-氯仿提取的DNA溶液26。
- 确定DNA浓度和纯度, 例如,通过紫外线吸光率或其他合适的方法。高效转录翻译,重要的是要使用高纯度的DNA。
2。准备薄的脂质膜
- 称量干脂质粉末溶解在溶剂中。对于1 -棕榈酰-2 -油酰-SN-甘油-3 -磷酸胆碱(POPC),储备溶液的制备在氯仿中,在浓度为40毫克/毫升。
注:有机溶剂必须始终处理的玻璃吸管和瓶子。换句话说,决不能使用塑料。商店s滴答的解决方案中的气密性,抗溶剂的琥珀色玻璃瓶在-20℃下,优选在氩气下。 - 等分试样12微摩尔到5毫升的圆底烧瓶中加入220微升40毫克/毫升原液(POPC)。
- 减压蒸除溶剂,用旋转蒸发器( 图1),以产生薄的脂质膜。
- 牢固地连接的圆底烧瓶中,蒸馏管,有一个圆形的剪辑,并开始在烧瓶的旋转。
- 启动的循环水通过冷凝器盘管。使用的热浴中,是没有必要的,因为氯仿61.2°C在正常大气压下具有低的沸点。
- 确保检查活塞打开,该系统是开放的气氛。打开真空泵慢慢地关闭活栓到系统应用真空。
- 的薄的脂质膜沉积在圆底烧瓶中的壁形成一个不透明的膜在旋转蒸发这是肉眼可见的。让旋转蒸发持续0.5-2小时。要停止的过程中,慢慢释放真空压力,停止了转动,取下圆底烧瓶中。
3。脂质再悬浮和囊泡同质化
- 薄的脂质膜,直接在圆底烧瓶中加入1毫升18.2MΩH 2 O。溶液剧烈振荡以最大的速度,约每分钟3200转,从玻璃上分离,用肉眼可观察到脂质膜。
- 脂质分散转移到2毫升离心管中。
- 建立了一个环站举行匀浆5毫米的小费。将下面的均质牢牢固定脂质分散离心立场。冲洗的均化分散体浸渍18.2MΩH 2 O中的前端和运行几秒钟。
- 将分散体直接将脂质分散体,以确保次的尖端处的微量离心管的底部不接触。均质功率4级或14000转1分钟。
4。囊泡挤压和冻干
- 的组装部件的小型挤出机( 图2),它由一个外壳和容纳两个聚四氟乙烯内部膜支持,两个O形环,和一个特富龙轴承的保持器螺母。
- 将两个O形圈进入膜内部支持的凹槽。预湿两个过滤器和一个400nm的膜。该过滤器将被放置对聚四氟乙烯O型圈的内侧,与它们之间的膜。
- 将膜放置在支持到挤出机中的外壳与膜包围的两个过滤器之间的O形环。将特富龙轴承外壳内,并附加固定螺母并用手拧紧。
- 冲洗两个注射器,并填补一个18.2MΩ水的。将注射器针头插入到小的holes聚四氟乙烯的挤出机组件的任一端上。针应该很容易滑动,不要强行针。固定注射器进入挤出机挤出住房和紧固。
- 通过挤出机出来的水缓慢地推入一个注射器和成其他的通水。这代表一个通道。重复一共有三个通道,确保不存在任何泄漏。取下注射器水和处置的水。
- 与样品填充一个注射器,连接到挤出机中,并如上所述在步骤4.3中,通过膜囊泡溶液缓慢通过。重复10倍,总的11通道。由于挤压过程的进行,样品会变得混浊,更容易推动穿过膜。突然减少阻力,但是,通常表示一个破裂的膜。
- 挤出囊泡溶液转移到微量离心管中,将40μl等分的囊泡。闪光冻结各等份,在任一干冰或液氮。
- 用离心蒸发器在30℃下过夜冻干各等份存储冻干空的囊泡在-20°C
5。封装转录翻译机械
- 转录 - 翻译反应中混合各成分,并加入20个单位的RNA酶抑制剂。在冰上孵育。
- 添加DNA模板。控制反应,250毫微克质粒编码mVenus的或类似的荧光蛋白后面的T7转录启动子和一个强大的E.大肠杆菌核糖体结合位点的建议。
- 用无RNA酶的水使终体积为25微升。
- 水合物冻干囊泡的等分试样(来自步骤4.6)与10μl的反应组装在步骤5.3中。简言之涡的混合物,直至囊泡悬浮。这应小于30秒。
- 3在冰上孵育反应0分钟,以允许膨胀的小泡。
- 稀释20倍,至终体积为30μl加入到27.0微升的50mM Tris-盐酸,50 mM氯化钠,pH值7.4和20.2毫克/毫升的蛋白酶K DNA酶和RNA酶1.5微升1.5微升囊泡的囊泡混合物也可以被添加在这一点上,作为一种替代蛋白酶K降解extravesicular材料。
- 孵育至少2.5小时,在37℃下
6。显微镜
- 检查在不同时间点的囊泡和荧光蛋白的生产进度。囊泡将有较大的直径大于400nm的孔隙的膜用于囊泡挤出的大小。
- 准备样品室放置一个20×5毫米的硅垫片到一个标准的显微镜幻灯片。吸取10微升囊泡进入样品室。在室放置一个硅化玻璃盖玻片。
- 观察囊泡与63X油分散Ø类似ŗ的明场和荧光显微镜使用适当的过滤器设置为利用荧光蛋白的目的。
Representative Results
荧光显微镜揭示了该荧光观察囊泡内,因为extravesicular材料酶降解( 图3)。 ,的intravesicular荧光如果在mVenus表达开始1.5小时后,观察到在37℃下,6小时内达到最大荧光强度。最佳的温度和温育时间可以有所不同,这取决于所使用的特定构造。例如,不同的荧光蛋白的成熟的完全不同的温度依赖性。换句话说,观察蛋白质生产并不仅仅取决于蛋白质的合成和折叠,但对发色基团的形成。总体蛋白质的合成,可以增加通过整合膜蛋白毛孔允许耗尽后,组件27的转录和翻译所必需的大量涌入。
建议开展一个类似的转录的翻译在囊泡的情况下,以确保利用的遗传构建功能重刑反应。该控制反应更容易监测荧光光谱法,而不是显微镜图4示出了在体外转录-翻译一个构建体编码mVenus的反应。未封装的反应给远高于总荧光强度比同类intravesicular反应。这是由于包封率,因为总的切除此体积比extravesicular体积( 即稀释的效果)要少得多。

图1。旋转蒸发器和真空泵等 。 点击这里查看大图 。
图2所示的挤出机的外壳和部件分开 。从左至右黑色O型圈面对彼此,挤出机的外壳,和第二注射器,注射器,固定螺母,聚四氟乙烯轴承,膜内部支持, 点击这里查看大图 。
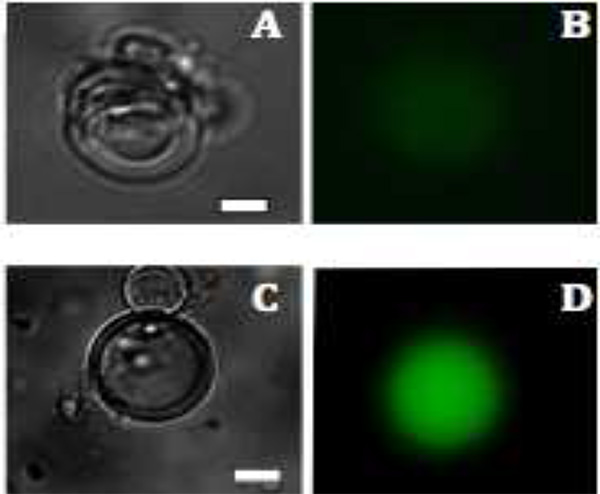
图3。荧光图像,在脂质体中的蛋白质生产mVenus A和C:明场图像多层囊泡1.5小时和2.5小时后,B&#160,D:的生产mVenus是荧光(颜色为绿色)后分别为1.5和2.5小时,可视化。比例尺为20微米。 点击此处查看大图 。
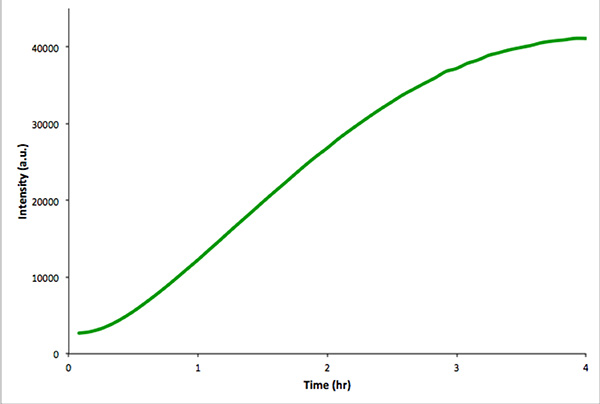
图4。 在体外控制反应的未封装的体外转录和翻译的mVenus中 。测定荧光强度超过4小时,每5分钟。被收购的数据在实时PCR仪。 点击此处查看大图 。
Discussion
虽然无细胞合成生物学是仍处于起步阶段,进展奠定了基础,日益复杂的细胞样系统可以。完全定义的组件内囊泡28的重组转录翻译机器尤为显着,在促进以后努力建设环境敏感人工cells17 18。同样地,人工细胞的研究已被用来探测进化过程4,29,30,RNA和蛋白质的合成22,31,影响代谢load32 33,和组装病毒颗粒34的机理的细节。足够的知识,更重要的是,现在存在里面泡在实验室后,这些先前的报告和本文所描述的协议,可以重组,基本的细胞功能。
除了容易,所描述的封装程序ocedure有几个好处。例如,许多空的,冻干的囊泡等分试样可以预先制造,并储存在-20℃下以供以后使用。该协议并不受生物分子的有机溶剂,温度急剧变化或长时间的透析。我们预计,温柔的程序,会促进将因为需要额外的元件。我们还没有观察到改变的脂质组成的膜封装或转录翻译效率的不利影响。因此,脂质膜蛋白,具体的形态,或可视化纳入更适合可以想见,被人利用。
上述方法的主要限制是,由此产生的囊泡大小和层数(lamellarity)不是同质的。对于许多应用,这些困难不会干扰对数据的解释。然而,如果需要的话,额外的步骤可以并入窄次E尺寸分布和减少层膜,如进一步挤压轮封装后,冻结解冻,或透析35。无疑是更好的方法规避这些和其他问题将得到发展。直到这时,我们发现这里描述的协议非常适合蜂窝模仿建设。
Disclosures
什么都没有透露。
Acknowledgments
作者承认Armenise哈佛基金会,玛丽 - 居里特伦蒂诺COFUND(ACS)自治省特伦托(ECOMM),和CIBIO资金。
Materials
| Name | Company | Catalog Number | Comments |
| Quick Spin Mini-prep kit | Qiagen | 27104 | |
| Spectrometer | NanoDrop 1000 | NDB767ND | |
| POPC | Avanti Polar Lipids | 770557 | MW 760 g/mol Transition Temp -2 °C CAS# 26853-31-6 |
| Ethanol | Sigma Aldrich | 459836 | Anhydrous, >99.5% |
| Phenol-choloroform-isoamyl alcohol 25:24:1, for molecular biology use | Sigma Aldrich | P3803-100mL | Saturated with 10 mM Tris, pH 8.0, 1 mM EDTA |
| Chloroform | Biotech Grade Fluka | 496189-1L | Contain ethanol at 0.5-1.0% v/v as stabilizer |
| Brown amber glass bottles | VWR | 89043-518 | 55X 48 mm |
| Rotary evaporator | Buchi Rotovapor R-210/Sigma | Z563846EU-1EA | With jack and water bath, 29/32 joint 240 V |
| Analog vortex mixer | VWR | 945300 | Speed 1,000-3,200 rpm |
| Homogenizer | IKA T10 Basic Ultra-Turbax | 3420000 | |
| Mini-extruder | Avanti Polar Lipids | 610020 | |
| Extruder filters | Whatman | 610014 | drain disc 10 mm |
| Extruder polycarbonate membrane 400 nm | Whatman | 61007 | nuclepore polycarbonate |
| Speed vacuum | Labconco | 7970011 | Centritrap DNA concentrator |
| PURExpress kit | New England Biolabs | NRM #E6800S | |
| RNAse inhibitor (40,000 U/ml) | New England Biolabs | #M0307S | |
| Proteinase K (20.2 mg/ml) | Fermentas | #EO0491 | |
| Microscope | Zeiss Observer Z1with a AxioCam MRm camera | ||
| RealTime | CFX96 Real time PCR Detection System (Biorad) | ||
| Silicon press to seal -Molecular Probe | Life Technologies | P18174 | Resistant from -25-30 °C |
| Siliconized glass circle cover slides | Hampton Research | HR3-231 | Diameter= 22 mm |
| ImageJ | NIH |
References
- Forster, A. C., Church, G. M. Towards synthesis of a minimal cell. Mol. Syst. Biol. 2, 1-10 (2006).
- Noireaux, V., Maeda, Y. T., Libchaber, A. Development of an artificial cell, from self-organization to computation and self-reproduction. Proc. Natl. Acad. Sci. U.S.A. 108, 3473-3480 (2011).
- Harris, D. C., Jewett, M. C. Cell-free biology: Exploiting the interface between synthetic biology and synthetic chemistry. Curr. Opin. Biotech. 23, (2012).
- Nishikawa, T., Sunami, T., Matsuura, T., Yomo, T. Directed Evolution of Proteins through In Vitro Protein Synthesis in Liposomes. J. Nucleic Acids. 2012, 1-11 (2012).
- Forlin, M., Lentini, R., Mansy, S. S.
- Chiarabelli, C., Stano, P., Anella, F., Carrara, P., Luisi, P. L. Approaches to chemical synthetic biology. FEBS Lett. 586, 2138-2145 (2012).
- Noireaux, V., Shin, J. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. J. Biol. Eng. 4, 1-9 (2010).
- Fujiwara, K., Nomura, S. -iM. Condensation of an Additive-Free Cell Extract to Mimic the Conditions of Live Cells. PLoS ONE. 8, e54155 (2013).
- Shimizu, Y., et al. Cell-free translation reconstituted with purified components. Nat. Biotechnol. 19, 751-755 (2001).
- Hosoda, K., et al. Quantitative Study of the Structure of Multilamellar Giant Liposomes As a Container of Protein Synthesis Reaction. Langmuir. 24, 13540-13548 (2008).
- Sunami, T., Matsuura, T., Suzuki, H., Yomo, T.
- Murtas, G., Kuruma, Y., Bianchini, P., Diaspro, A., Luisi, P. L. Protein synthesis in liposomes with a minimal set of enzymes. Biochem. Biophys. Res. Commun. 363, 12-17 (2007).
- Pereira de Souza, T., Stano, P., Luisi, P. L. The Minimal Size of Liposome-Based Model Cells Brings about a Remarkably Enhanced Entrapment and Protein Synthesis. ChemBioChem. 10, 1056-1063 (2009).
- Caschera, F., et al. Programmed Vesicle Fusion Triggers Gene Expression. Langmuir. 27, 13082-13090 (2011).
- Noireaux, V., Bar-Ziv, R., Godefroy, J., Salman, H., Libchaber, A. Toward an artificial cell based on gene expression in vesicles. Phys. Biol. 2, P1-P8 (2005).
- Shin, J., Noireaux, V. An E. coli Cell-Free Expression Toolbox: Application to Synthetic Gene Circuits and Artificial Cells. ACS Synth. Biol. 1, 29-41 (2012).
- Kobori, S., Ichihashi, N., Kazuta, Y., Yomo, T. A controllable gene expression system in liposomes that includes a postive feedback loop. Mol. Syst. Biol. 9, 1282-1285 (2013).
- Martini, L., Mansy, S. S. Cell-like Systems with Riboswitch Controlled Gene Expression. Chem. Commun. 47, 10734-10736 (2011).
- Noireaux, V., Bar-Ziv, R., Libchaber, A. Principles of cell-free genetic circuit assembly. Proc. Natl. Acad. Sci. U.S.A. 100, 12672-12677 (2003).
- Sunami, T., et al. Detection of Association and Fusion of Giant Vesicles Using a Fluorescence-Activated Cell Sorter. Langmuir. 26, 15098-15103 (2010).
- Saito, H., et al. Time-Resolved Tracking of a Minimum Gene Expression System Reconstituted in Giant Liposomes. ChemBioChem. 10, 1640-1643 (2009).
- Lentini, R., et al. Fluorescent Proteins and in Vitro Genetic Organization for Cell-Free Synthetic Biology. ACS Synth. Biol. , (2013).
- Du, L., Villarreal, S., Forster, A. C. Multigene Expression In Vivo: Supremacy of Large Versus Small Terminators for T7 RNA Polymerase. Biotechnol. Bioeng. 109, 1043-1050 (2011).
- Trochilin, V. P., Weissig, V. Liposomes: A Practical Approach. , 2nd edn, Oxford University Press. (2003).
- Walde, P., Cosentino, K., Engel, H., Stano, P. Giant vesicles: preparations and applications. ChemBioChem. 11, 848-865 (2010).
- Sambrook, J., Russell, D. W. Molecular Cloning. , 3rd edn, Cold Spring Harbor Laboratory Press. (2001).
- Noireaux, V., Libchaber, A. A vesicle bioreactor as a step toward an artificial cell assembly. Proc. Natl. Acad. Sci. U.S.A. 101, 17669-17674 (2004).
- Yu, W., et al.
- Caschera, F., et al. Stable vesicles composed of monocarboxylic or dicarboxylic fatty acids and trimethylammonium amphiphiles. Langmuir. 27, 14078-14090 (2011).
- Pereira de Souza, T., Steiniger, F., Stano, P., Fahr, A., Luisi, P. L. Spontaneous crowding of ribosomes and proteins inside vesicles: a possible mechanism for the origin of cell metabolism. ChemBioChem. 12, 2325-2330 (2011).
- Niederholtmeyer, H., Xu, L., Maerkl, S. J. Real-Time mRNA Measurement during an in Vitro Transcription and Translation Reaction Using Binary Probes. ACS Synth. Biol. 10, (2012).
- Stögbauer, T., Windhager, L., Zimmer, R., Rädler, J. Experiment and mathematical modeling of gene expression dynamics in a cell-free system. Integr. Biol. 4, 494-501 (2012).
- Lazzerini-Ospri, L., Stano, P., Luisi, P., Marangoni, R. Characterization of the emergent properties of a synthetic quasi-cellular system. BMC Bioinformatics. 13, 1-10 (2011).
- Shin, J., Jardine, P., Noireaux, V. Genome Replication, Synthesis, and Assembly of the Bacteriophage T7 in a Single Cell-Free Reaction. ACS Synth. Biol. 1, 408-413 (2012).
- Zhu, T. F., Szostak, J. W.
