

Summary
Häri beskriver vi enkla metoder för framställning av vesiklar, inkapsling av transkription och translation maskiner och övervakning av protein produktion. De resulterande cellfria system kan användas som utgångspunkt för att bygga allt mer komplexa cellulära härmar.
Abstract
Som räntor skiftar från enskilda molekyler till system av molekyler, har ett ökande antal laboratorier försökt bygga underifrån cellulära härmar som bättre representerar komplexa cellulära liv. Hittills finns det ett antal vägar som kan vidtas för att bygga compartmentalized cellulära härmar, inklusive utnyttjande av vatten-i-olja emulsioner, mikrofluidikanordningar och blåsor. Var och en av de tillgängliga alternativen har för-och nackdelar. Till exempel, vatten-i-olja-emulsioner ge hög inkapslingseffektivitet men inte efterlikna väl permeabilitetsbarriären av levande celler. Den primära fördelen med de metoder som beskrivs häri är att de alla är lätt och billig att genomföra. Transkription-translation maskiner är inkapslat inuti fosfolipidvesiklar genom en process som utnyttjar gemensamma instrumentering, såsom en centrifugalindunstare och en extruder. Reaktionerna övervakas av fluorescensspektroskopi. Protokollets kan anpassas för rekombinant proteinuttryck, byggandet av cellulära härmar, utforskandet av minimikrav för cell-liv, eller montering av genetiska kretsar.
Introduction
Cell-fri, in vitro transkription-översättning reaktioner och generering av blåsor från syntetiska lipider är inget nytt. Att kombinera de två till ett cellulärt härma är betydligt mer challenging1-6. E. coli-cellextrakt med eller utan T7 RNA-polymeras kan användas som en källa för transkriptions-translations maskiner 7,8. Cellextrakt gynnas av närvaron av ytterligare cellulära komponenter som kan underlätta proteinexpression och vikning. Alternativt kan en blandning av individuellt renade RNA och proteinmolekyler, dvs PURE-systemet 9, kan användas för att mediera intravesikulär proteinsyntes 4,10-14. PURE system möjliggör konstruktion av helt definierade cellulära härmar och inte lider av nukleasaktivitet hittades i cellextrakt. Praktiskt innebär detta att mycket mindre DNA-templat erfordras, vilket underlättar processer med låg inkapslingseffektivitet 11 16-18.
Det enklaste sättet att övervaka transkriptions-translationsreaktioner är att mäta fluorescensen eller luminescensen av genetiskt kodade element. Typiskt är eldflugeluciferas 19 eller GFP används, även om in vitro reaktioner ofta mäts genom radioaktiv märkning. Fluorescensdetektion tillåter dessutom för övervakning av populationer av vesiklar 20,21 genom cytometry baserade metoder, vilket ger en inblick i den stokastiska karaktären av biologisk-liknande processer. Dessa kontrollprogram metoder har använts för att definiera en liten uppsättning designregler och ett bibliotek av delar för att bygga från, inklusive en samling av fluorescerande proteiner som är kompatibla med in vitro transkription-translation 22, påverkan av genetiska organisation på uttryck 22, aktiviteten av sigma faktorer 16, och effektiviteten av transkriptionella terminatorer 23. Ändå finns det fortfarande mycket som behöver göras för att öka förmågan att bygga förutsägbar in vitro, genetiskt kodade enheter.
Det finns många metoder tillgängliga för att göra vesiklar. De vanligaste metoderna är beroende av genereringen av en tunn lipidfilm på en glasyta med påföljande återsuspension i vattenhaltig solution24. Om den vattenhaltiga lösningen innehåller transkriptions-translations maskiner, till exempel, då en fraktion av vesiklarna bildade skulle innehålla nödvändiga komponenter för proteinproduktion. Emellertid är inkapslingseffektiviteten av sådana metoder låg, vilket innebär att endast en liten andel av vesiklar är aktiva. Många av de alternativa metoder som kännetecknas av mycket högre inkapsling efficiency utnyttja omvandlingen av vatten-i-olja emulsionsdroppar till vesiklar. Även om det är troligt att sådana metoder kommer att vara vardagsmat i framtiden, för närvarande dessa metoder lider av behovet av specialiserad utrustning och ge blåsor med förändrade membran kompositioner 25. En tydlig fördel med vatten-i-olja till vesikler metoder är möjligheten att styra membranet lamellarity. Metoden som beskrivs häri är baserad på den tunna lipidfilmen protokollet beskrivet av den Yomo laboratorium 11 med smärre ändringar, inklusive ett ytterligare homogeniseringssteg. Denna metod är enkel, billig, och ger robusta vesiklar väl lämpade för inkapsling av transkriptions-translations maskiner.
Protocol
Ett. Förberedelse av DNA-mall
- Rena plasmid från en standard laboratorium stam av E. coli, såsom E. coli DH5a eller Nova Blue med ett kommersiellt kit. Alternativt kan en linjär PCR-produkt på liknande renas med ett kommersiellt kit. Eluera DNA med H2O bara.
- Fenol-kloroform extrahera DNA-lösning 26.
- Bestäm DNA-koncentration och renhet, t.ex. genom UV-absorbans eller andra lämpliga metoder. Det är viktigt att använda högren DNA för effektiv transkription-translation.
2. Förbereda Thin lipidfilmen
- Väg det torra lipidpulvret och lös i lösningsmedel. För 1-palmitoyl-2-oleoyl-sn-glycero-3-fosfokolin (POPC), är förrådslösningar framställdes i kloroform vid en koncentration av 40 mg / ml.
Obs: Organiska lösningsmedel måste alltid hanteras med glaspipetter och flaskor. Med andra ord måste plast aldrig användas. Affär stock lösningar i lufttäta, lösningsmedelsbeständiga flaskor bärnstensfärgat glas vid -20 ° C, företrädesvis under argon. - Alikvot 12 imol POPC (220 pl av 40 mg / ml stamlösning) i en 5 ml rundbottnad kolv.
- Indunsta lösningsmedlet i en rotationsindunstare (figur 1) för att generera den tunna lipidfilm.
- Säkert fästa rundkolv till destillationsröret med en cirkulär klämma och starta rotationen av kolven.
- Initiera cirkulation av vatten genom kondensorslingan. Med hjälp av en värme-bad är inte nödvändigt, eftersom kloroform har en låg kokpunkt av 61,2 ° C vid normalt atmosfärstryck.
- Kontrollera att systemet är öppen mot atmosfären genom att kontrollera att kranen är öppen. Slå på vakuumpumpen och långsamt stänga kranen för att applicera vakuum till systemet.
- En tunn lipidfilm avsätts på väggarna i rundkolv under rotationsindunstning bildar ett täckande skiktsom är synlig med blotta ögat. Låt rotationsindunstning fortsätta under 0,5-2 timmar. För att stoppa processen, långsamt släppa undertrycket, stoppa rotationen, och ta bort rundbottnad kolv.
Tre. Lipid Resuspension och Vesicle homogenisering
- Tillsätt 1 ml 18,2 MQ H2O till den tunna lipidfilmen direkt i den rundbottnade kolven. Kraftigt virvel lösningen vid maximal hastighet, ca 3.200 rpm, tills Lipidfilmen lossnar från glaset, som kan observeras med ögat.
- Överför lipiddispersion i en 2 ml mikrocentrifugrör.
- Ställ upp en ring står för att hålla en homogenisator med en 5 mm spets. Placera en mikrocentrifug monter under homogeniseraren att säkert hålla lipiddispersion. Skölj dispergerande elementet i homogenisatorn genom nedsänkning av spetsen i 18,2 Mohm H2O och kör i några sekunder.
- Placera den spridande elementet direkt i lipiddispersion säkerställer thvid spetsen inte vidrör botten av mikrocentrifugrör. Homogenisera under 1 min vid effektnivå 4 eller 14.000 rpm.
4. Vesikel Extrudering och lyofilisering
- Montera de delar av mini-extruder (figur 2), som består av ett yttre hölje och en hållare mutter som rymmer två Teflon inre membran stödjer, två O-ringar, och en Teflon bäring.
- Placera de två O-ringarna in i spåren i de inre membran stöden. Prewet två filter och en 400 nm-membran. Filtren ska placeras mot Teflon insidan av O-ringarna med membranet mellan dem.
- Placera membranet stöd i strängsprutmaskinen ytterhölje med membran som omges av två filter i mellan O-ringarna. Placera Teflon lagret inuti höljet och fäst hållaren muttern och dra åt för hand.
- Skölj två sprutor och fylla en med 18,2 Mohm vatten. Sätt in kanyler i den lilla holes i teflon på vardera änden av strängpressningsenheten. Nålarna ska glida in lätt, inte tvinga nålarna. Säkra extruder med sprutor i den extruder bostäder och sy.
- Passera vattnet genom strängsprutmaskinen genom att långsamt trycka vatten ur en spruta och in i den andra. Detta utgör en passage. Upprepa för totalt tre passager som säkerställer att det inte finns några läckor närvarande. Avlägsna sprutan med vatten och kasta vattnet.
- Fyll en spruta med provet, fäst till extrudern, och långsamt passera vesikel lösningen genom membranet såsom beskrivits ovan i steg 4.3. Upprepa 10x för totalt 11 passager. Som behållningen strängsprutningsprocess, kommer provet att bli mindre grumliga och lättare att trycka över membranet. En plötslig minskning i motstånd, dock oftast indikerar en bristning av membranet.
- Överför extruderade vesikellösning och göra 40 pl alikvoter av blåsor i mikrocentrifugrör.Flash frysa varje portion i torris eller i flytande kväve.
- Lyofilisera varje alikvot med en centrifugalindunstare natten vid 30 ° C. Förvara frystorkade tomma blåsor vid -20 ° C.
Fem. Encapsulating Transkription-translation Machinery
- Blanda komponenterna i transkriptions-translationsreaktion och tillsätt 20 enheter av RNas-inhibitor. Inkubera på is.
- Tillsätt DNA-mallen. För en kontrollreaktion, 250 ng av en plasmid som kodar mVenus eller en liknande fluorescerande protein bakom en T7 transkriptionspromotor och en stark E. coli ribosombindningsställe är rekommenderas.
- Bringa den slutliga volymen till 25 | il med RNas-fritt vatten.
- Hydrat en alikvot av lyofiliserade vesiklar (från steg 4,6) med 10 | il av reaktionen samman i moment 5.3. Kortfattat virvel blandningen tills vesiklarna suspenderas. Detta bör ta mindre än 30 sekunder.
- Inkubera reaktionen på is under 30 min för att tillåta vesiklarna att svälla.
- Späd vesikel blandning 20-faldigt till en slutlig volym på 30 pl genom tillsats av 1,5 | il av vesiklar i 27,0 | il av 50 mM Tris-HCl, 50 mM NaCl, pH 7,4 och 1,5 | il av 20,2 mg / ml proteinas K. DNas och RNas kan också tillsättas vid denna punkt som ett alternativ till proteinas K att nedbryta extravesicular material.
- Inkubera under minst 2,5 h vid 37 ° C.
6. Mikroskopi
- Undersök blåsor och framstegen i fluorescerande proteinet produktionen vid olika tidpunkter. Vesiklarna kommer att ha en större diameter än 400 nm porstorlek hos membranet som används för vesikel extrudering.
- Förbered en provkammare genom att placera en 20 x 5 mm spacer kisel på en standard objektglas. Pipettera 10 | il av vesiklar in i provkammaren. Placera en silikoniserad slip glaskupa över kammaren.
- Observera blåsor med en 63x olja-dispersion or liknande mål med ljusa fält och fluorescens mikroskopi med lämpligt filter set för exploaterade fluorescerande proteinet.
Representative Results
Fluorescens mikroskopi avslöjar att fluorescens endast observeras inuti av vesiklarna, eftersom extravesicular material är enzymatiskt nedbrutna (fig 3). För expression av mVenus börjar intravesikulär fluorescens som skall passera efter 1,5 h vid 37 ° C och når maximal fluorescensintensiteten inom 6 tim. Den optimala temperaturen och inkubationstiden kan variera beroende på de använda specifika konstruktionerna. Exempelvis kan olika fluorescerande proteiner mogna helt annorlunda i en temperatur beroende sätt. Med andra ord, är observation av proteinproduktion inte enbart beroende av proteinsyntes och vikning utan också på kromofor bildas. Total proteinsyntes kan ökas genom att införliva porer membranprotein för att möjliggöra ett inflöde av utarmat komponenter som krävs för transkription och translation 27.
Det rekommenderas att utföra en analog transkription-översätttion reaktionen i frånvaro av vesiklar för att säkerställa att de utsugna genetiska konstruktionen är funktionell. Denna kontroll reaktion är lättare övervakas av fluorescensspektroskopi snarare än mikroskopi. Visar en in vitro transkription-translationsreaktion av en konstruktion som kodar mVenus Figur 4. Oinkapslade reaktioner ger mycket högre totala fluorescensintensiteter än liknande intravesikulär reaktioner. Detta beror på inkapslingseffektivitet och eftersom den totala intravesikulär volym är mycket mindre än den extravesicular volym (dvs. en utspädningseffekt).

Figur 1. Den roterande förångare och vakuumpump. Klicka här för att se större bild .
Figur 2. Höljet och delar av extruder visas separat. Från vänster till höger, stöder spruta, hållare mutter, teflon lagret, inre membran med svarta O-ringar inför varandra, extruder yttre höljet och andra sprutan. Klicka här för att se större bild .
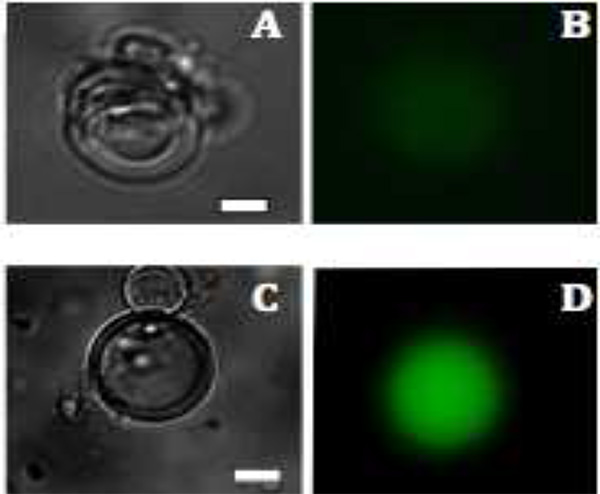
. Figur 3 fluorescens bilder av mVenus protein produktion i liposomer A och C:.. Ljusfält bilder av multilamellära vesiklar efter 1,5 h och 2,5 h B &# 160, D: Produktionen av mVenus visualiseras genom fluorescens (grönfärgad) efter 1,5 och 2,5 timmar, respektive. Skala bar är 20 um. Klicka här för att se större bild .
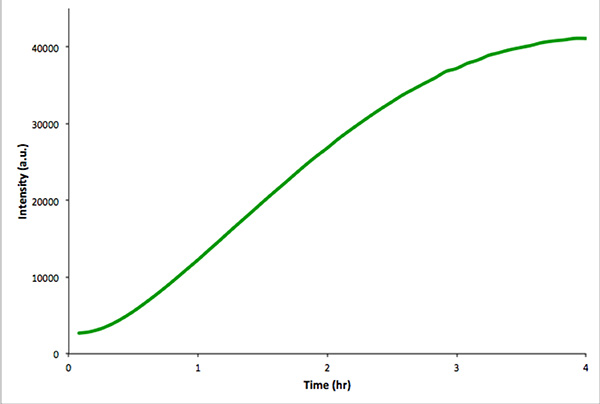
Figur 4. In vitro kontrollreaktion av nonencapsulated in vitro transkription och translation av mVenus. Fluorescensintensitet mättes varje 5 min under 4 timmar. Uppgifterna har förvärvats med en Real-Time PCR instrument. Klicka här för att se större bild .
Discussion
Även cell-fri syntetisk biologi är fortfarande i sin linda, har framsteg lagt en grund från vilken alltmer komplexa cell-liknande system kan göras. Den beredning av transkription-translation maskiner från helt definierade komponenter 9 inuti vesiklar 28 var särskilt viktig för att underlätta senare ansträngningar att bygga miljömässigt lyhörd artificiell cells17, 18. På liknande sätt har konstgjorda cellstudier använts för att proba evolutionära processer 4,29,30, de mekanistiska detaljerna av RNA-och proteinsyntes 22,31, inflytande av metabolisk load32, 33, och montering av virala partiklar 34. Huvudsakligen finns tillräckligt med kunskap nu när grundläggande cellulär funktion kan beredas insidan av blåsor i laboratoriet efter dessa tidigare rapporter och protokoll som beskrivs här.
Förutom att vara lätt, den beskrivna inkapsling procedure har flera fördelar. Till exempel kan många tomma, frystorkade vesikler portioner göras i förväg och förvaras vid -20 ° C för senare användning. Protokollet inte omfattas biologiska molekyler för organiska lösningsmedel, drastiska temperaturförändringar, eller långa perioder av dialys. Vi förväntar oss att mildhet av förfarandet kommer att underlätta införandet av ytterligare komponenter som behövs. Vi har också inte har observerat negativa effekter till förändrade lipidkompositionen av membranet på inkapsling eller transkriptions-translations effektivitet. Därför kunde lipider mer mottagliga för införlivandet av membranproteiner, särskilda morfologier, eller visualisering tänkas utnyttjas.
Den största begränsningen av den beskrivna metoden är att de resulterande vesiklarna inte är homogena i storlek eller lamellarity. För många tillämpningar, stör dessa svårigheter inte med tolkning av data. Men, om så behövs, kan ytterligare steg införlivas för att smala the storleksfördelning och minska lagren av membran, såsom ytterligare omgångar strängsprutning efter inkapsling, frysning-tining, eller dialys 35. Otvivelaktigt bättre metoder att kringgå dessa och andra problem kommer att utvecklas. Tills dess, finner vi det protokoll som beskrivs här för att vara väl lämpad för byggandet av cellulära härmar.
Disclosures
Författarna har ingenting att lämna ut.
Acknowledgments
Författarna erkänner Armenise-Harvard Foundation, Marie-Curie Trentino Cofund (ACS), den autonoma provinsen Trento (Ecomm), och CIBIO för finansiering.
Materials
| Name | Company | Catalog Number | Comments |
| Quick Spin Mini-prep kit | Qiagen | 27104 | |
| Spectrometer | NanoDrop 1000 | NDB767ND | |
| POPC | Avanti Polar Lipids | 770557 | MW 760 g/mol Transition Temp -2 °C CAS# 26853-31-6 |
| Ethanol | Sigma Aldrich | 459836 | Anhydrous, >99.5% |
| Phenol-choloroform-isoamyl alcohol 25:24:1, for molecular biology use | Sigma Aldrich | P3803-100mL | Saturated with 10 mM Tris, pH 8.0, 1 mM EDTA |
| Chloroform | Biotech Grade Fluka | 496189-1L | Contain ethanol at 0.5-1.0% v/v as stabilizer |
| Brown amber glass bottles | VWR | 89043-518 | 55X 48 mm |
| Rotary evaporator | Buchi Rotovapor R-210/Sigma | Z563846EU-1EA | With jack and water bath, 29/32 joint 240 V |
| Analog vortex mixer | VWR | 945300 | Speed 1,000-3,200 rpm |
| Homogenizer | IKA T10 Basic Ultra-Turbax | 3420000 | |
| Mini-extruder | Avanti Polar Lipids | 610020 | |
| Extruder filters | Whatman | 610014 | drain disc 10 mm |
| Extruder polycarbonate membrane 400 nm | Whatman | 61007 | nuclepore polycarbonate |
| Speed vacuum | Labconco | 7970011 | Centritrap DNA concentrator |
| PURExpress kit | New England Biolabs | NRM #E6800S | |
| RNAse inhibitor (40,000 U/ml) | New England Biolabs | #M0307S | |
| Proteinase K (20.2 mg/ml) | Fermentas | #EO0491 | |
| Microscope | Zeiss Observer Z1with a AxioCam MRm camera | ||
| RealTime | CFX96 Real time PCR Detection System (Biorad) | ||
| Silicon press to seal -Molecular Probe | Life Technologies | P18174 | Resistant from -25-30 °C |
| Siliconized glass circle cover slides | Hampton Research | HR3-231 | Diameter= 22 mm |
| ImageJ | NIH |
References
- Forster, A. C., Church, G. M. Towards synthesis of a minimal cell. Mol. Syst. Biol. 2, 1-10 (2006).
- Noireaux, V., Maeda, Y. T., Libchaber, A. Development of an artificial cell, from self-organization to computation and self-reproduction. Proc. Natl. Acad. Sci. U.S.A. 108, 3473-3480 (2011).
- Harris, D. C., Jewett, M. C. Cell-free biology: Exploiting the interface between synthetic biology and synthetic chemistry. Curr. Opin. Biotech. 23, (2012).
- Nishikawa, T., Sunami, T., Matsuura, T., Yomo, T. Directed Evolution of Proteins through In Vitro Protein Synthesis in Liposomes. J. Nucleic Acids. 2012, 1-11 (2012).
- Forlin, M., Lentini, R., Mansy, S. S.
- Chiarabelli, C., Stano, P., Anella, F., Carrara, P., Luisi, P. L. Approaches to chemical synthetic biology. FEBS Lett. 586, 2138-2145 (2012).
- Noireaux, V., Shin, J. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. J. Biol. Eng. 4, 1-9 (2010).
- Fujiwara, K., Nomura, S. -iM. Condensation of an Additive-Free Cell Extract to Mimic the Conditions of Live Cells. PLoS ONE. 8, e54155 (2013).
- Shimizu, Y., et al. Cell-free translation reconstituted with purified components. Nat. Biotechnol. 19, 751-755 (2001).
- Hosoda, K., et al. Quantitative Study of the Structure of Multilamellar Giant Liposomes As a Container of Protein Synthesis Reaction. Langmuir. 24, 13540-13548 (2008).
- Sunami, T., Matsuura, T., Suzuki, H., Yomo, T.
- Murtas, G., Kuruma, Y., Bianchini, P., Diaspro, A., Luisi, P. L. Protein synthesis in liposomes with a minimal set of enzymes. Biochem. Biophys. Res. Commun. 363, 12-17 (2007).
- Pereira de Souza, T., Stano, P., Luisi, P. L. The Minimal Size of Liposome-Based Model Cells Brings about a Remarkably Enhanced Entrapment and Protein Synthesis. ChemBioChem. 10, 1056-1063 (2009).
- Caschera, F., et al. Programmed Vesicle Fusion Triggers Gene Expression. Langmuir. 27, 13082-13090 (2011).
- Noireaux, V., Bar-Ziv, R., Godefroy, J., Salman, H., Libchaber, A. Toward an artificial cell based on gene expression in vesicles. Phys. Biol. 2, P1-P8 (2005).
- Shin, J., Noireaux, V. An E. coli Cell-Free Expression Toolbox: Application to Synthetic Gene Circuits and Artificial Cells. ACS Synth. Biol. 1, 29-41 (2012).
- Kobori, S., Ichihashi, N., Kazuta, Y., Yomo, T. A controllable gene expression system in liposomes that includes a postive feedback loop. Mol. Syst. Biol. 9, 1282-1285 (2013).
- Martini, L., Mansy, S. S. Cell-like Systems with Riboswitch Controlled Gene Expression. Chem. Commun. 47, 10734-10736 (2011).
- Noireaux, V., Bar-Ziv, R., Libchaber, A. Principles of cell-free genetic circuit assembly. Proc. Natl. Acad. Sci. U.S.A. 100, 12672-12677 (2003).
- Sunami, T., et al. Detection of Association and Fusion of Giant Vesicles Using a Fluorescence-Activated Cell Sorter. Langmuir. 26, 15098-15103 (2010).
- Saito, H., et al. Time-Resolved Tracking of a Minimum Gene Expression System Reconstituted in Giant Liposomes. ChemBioChem. 10, 1640-1643 (2009).
- Lentini, R., et al. Fluorescent Proteins and in Vitro Genetic Organization for Cell-Free Synthetic Biology. ACS Synth. Biol. , (2013).
- Du, L., Villarreal, S., Forster, A. C. Multigene Expression In Vivo: Supremacy of Large Versus Small Terminators for T7 RNA Polymerase. Biotechnol. Bioeng. 109, 1043-1050 (2011).
- Trochilin, V. P., Weissig, V. Liposomes: A Practical Approach. , 2nd edn, Oxford University Press. (2003).
- Walde, P., Cosentino, K., Engel, H., Stano, P. Giant vesicles: preparations and applications. ChemBioChem. 11, 848-865 (2010).
- Sambrook, J., Russell, D. W. Molecular Cloning. , 3rd edn, Cold Spring Harbor Laboratory Press. (2001).
- Noireaux, V., Libchaber, A. A vesicle bioreactor as a step toward an artificial cell assembly. Proc. Natl. Acad. Sci. U.S.A. 101, 17669-17674 (2004).
- Yu, W., et al.
- Caschera, F., et al. Stable vesicles composed of monocarboxylic or dicarboxylic fatty acids and trimethylammonium amphiphiles. Langmuir. 27, 14078-14090 (2011).
- Pereira de Souza, T., Steiniger, F., Stano, P., Fahr, A., Luisi, P. L. Spontaneous crowding of ribosomes and proteins inside vesicles: a possible mechanism for the origin of cell metabolism. ChemBioChem. 12, 2325-2330 (2011).
- Niederholtmeyer, H., Xu, L., Maerkl, S. J. Real-Time mRNA Measurement during an in Vitro Transcription and Translation Reaction Using Binary Probes. ACS Synth. Biol. 10, (2012).
- Stögbauer, T., Windhager, L., Zimmer, R., Rädler, J. Experiment and mathematical modeling of gene expression dynamics in a cell-free system. Integr. Biol. 4, 494-501 (2012).
- Lazzerini-Ospri, L., Stano, P., Luisi, P., Marangoni, R. Characterization of the emergent properties of a synthetic quasi-cellular system. BMC Bioinformatics. 13, 1-10 (2011).
- Shin, J., Jardine, P., Noireaux, V. Genome Replication, Synthesis, and Assembly of the Bacteriophage T7 in a Single Cell-Free Reaction. ACS Synth. Biol. 1, 408-413 (2012).
- Zhu, T. F., Szostak, J. W.
