

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
個人を自由に移動可能ではない分析のレベルで視覚的学習と記憶の形成を研究に研究を可能にするよう注意を払うアリの古典的条件づけのためのプロトコルを提案します。
Abstract
昆虫のいくつかの種は、学習と記憶の形成を研究するためのモデル システムになっています。多くの研究は動物を自由に移動に焦点を当てる、研究利用された昆虫と古典的条件づけパラダイムを実装する個人を学ぶ正確な手がかりと学習の神経機構を調査するため重要なされていると記憶の形成。ここで古典的条件付けで木蟻の連想学習を想起させるためのプロトコルを提案する.このパラダイムでアリが活かしたし、視覚的な合図 (青い段ボール)、条件刺激 (CS) 飛び廻る砂糖報酬、無条件刺激 (米国) と対になって表示されました。アリは、上顎唇伸展反射 (マラー) 学習の読み出しとして使用することができます無条件の応答 (UR) を実行します。トレーニングは、5 分間試行間間隔 (ITI) で区切られた 10 の試験で構成されます。アリもメモリ保持 10 分または 1 時間のトレーニング後にテストされます。このプロトコルには、高精度かつ制御された方法で、視覚的記憶形成の詳細は、木蟻で学習・記憶形成の神経基盤を分析する研究者を許可する可能性があります。
Introduction
昆虫は、学習・記憶形成1を勉強するためのモデルとして広く使用されています。研究の特に成功した静脈は学んだされている手がかりを正確に制御を可能にする拘束された動物の古典的条件づけを使用し、学習と記憶を支える神経回路メカニズム解明のための研究者を許可します。研究の大半は、ニホンミツバチ、セイヨウミツバチの飛び廻る古典的条件づけに焦点を当てています。ミツバチの労働者は CS を関連付ける UR を連想させる米国に訓練されます。このパラダイムでは、もともと武田2 Bittermanらによって開発されました。3UR はテング拡張反射 (PER)、米国、砂糖で、CS は臭い。蜂は、CS と米国間の関連付けを学ぶし、この協会の長期記憶を形成することができます。
UR としての PER を使用して元のパラダイムは蜂4嗅覚学習を支える神経経路を解明するために使用されています。異なる感覚刺激、視覚刺激5,6、7を含むあたり8またはスティングの拡張機能の抑制による嫌悪学習を組み込むことの学習をテストするいくつかの方法で変更されています。反射 (SER)9 UR とマルハナバチ10と果実ハエ11などの他の種の学習をテストします。記憶の形成の複数のモダリティとは、古典的条件づけを研究されている、視覚的に学習はまだこのアプローチを使用して観察することは困難種でも記載されているがビジュアル学習能力の高い餌中などミツバチ。
最近の研究は, 開口部の応答 (POR)12を実行すると、バッタ、アリ、上顎唇伸展反射 (マラー)13を実行など、テングを持たない昆虫に似たようなアプローチを適用しています。これは 2 つの異なる砂漠バッタの表現型、社交的な solitarious フォームは、記憶の形成に一致する必要がありますアイデアの再強制のフェーズに固有の供給戦略を一致フェーズに固有の学習能力を明らかにしています。エコロジー ニーズ14。さらに、アリの嗅覚学習研究で示しているアリやミツバチの類似性記憶の形成と長期記憶の保持と保持タンパク質合成15に依存することの訓練の後の 72 時間。
元パラダイムのこれらの適応は多くのモダリティといくつかのモデル種勉強する学習と記憶の形成を許可しています。これは昆虫にメモリの形成の共通のメカニズムを識別する学習と異なる種のメモリの特定の生態を反映しての違いを識別するためにも不可欠です。我々 はここで説明するプロトコルの主な目的は、広く研究アリ種、フォーマイカ西隣の視覚条件刺激を用いた古典的条件付け実験を実行する方法を提供します。また視覚的古典的条件づけパラダイムの適応である木アリ16学習の研究から発行されます。
Protocol
1.フォーマイカ西隣の植民地を維持します。
注: 本研究に使用 (フォーマイカ西隣L.) 木製のアリ ・ コロニーは、アシュダウン フォレスト、サセックス、イギリスから収集された (N 51 4.680、E 0 1.800)。英国内で 7 月と 8 月の間F. 西隣のコロニーを収集します。巣の良い割合を削除する必要があります、数百の労働者とひな、植民地を確保するため、持続的なを含むと長期間にわたってアクティブ時間 (1 年間)。巣除去前に関係当局から許可を得なければなりません。
- ワイド ボックスで木蟻コロニーの家 (約 135 cm × 70 cm × 35 cm) 12 h ライト: 12 下 21 ° C で、エスケープからアリを防ぐために (例えばフロン)、ポリテトラフルオロ エチレン (PTFE) 樹脂で覆われて壁-h 暗いサイクル。
- 水ディスペンサーを使用してアリにショ糖 (333 g/L) と水を提供します。アリの食事が蛋白質で豊富なことを確認、冷凍コオロギ 2 倍 1 週間でコロニーをフィードします。高温多湿に保つために、毎日水と巣をスプレーします。
- 植民地の衛生状態を改善するために松脂必要な場合を追加します。松の樹脂は、木蟻のコロニー17で寄生を低減します。アリはそれで死んだコロニー メンバーを沈殿できるように巣の中小さい容器 (約 15 センチ直径の 5 cm) を配置します。
- 実験を試みる前に収集した後、少なくとも 2 週間は、船首に記載の条件の下でアリを保つため。アリ意欲が向上、実験前に 2 d を餓死に巣からショ糖を削除します。
2 を選択してアリを活用
注: アリを活用するため特注のホルダーが必要です。モデリング粘土を使用してこれが構築でき、段ボールを開くカットの水平方向にそれに接続されています。段ボールの上面は部分的昆虫ピン固定中に添付することを許可するワックスでコーティングする必要があります。慎重な取り扱いは、トレーニング前にストレスの高いレベルにそれらを服従させることを避けるために、アリを活用このプロトコル (メンテナンス、トランスポート、および実験を含む)、しかし特にステップごとに必要です。
- アリを食べる意欲を選択すると、小さなボックス内スクロース (200 g/L) のドロップでスライドに配置 (約 14 cm × 8 cm × 5 cm) 壁脱出を防ぐために PTF 樹脂で覆われています。巣から木材や死んだアリを運んでいるアリを選択しないでください。代わりに、これらは食べ物を探すため巣を残してしようとして受動的探索担当になりやすいので、巣の壁を這っている蟻を取る。
- ボックスでそれぞれのアリを置き、かどうかそれはショ糖のドロップでフィードを参照してくださいするを待ちます。場合は、飽食を防ぐために別の空のボックスにすぐに転送します。
- それぞれのアリを別のチューブに転送します。アリを固定する各チューブに 1-2 分冷凍庫または最大 5 分の砕いた氷を配置します。
- 頭部と胸部がその間のジョイントによって、段ボールのカットをホルダーに固定された ant を配置します。アンテナを側頭部、厚紙の上にワックス層に埋め込まれている昆虫のピンを使用して確保します。
- カスタムメイドの加熱要素を使用して、ant の頭に昆虫ピンの先端を段ボールに平行にワックスします。この中に、それらへの損傷を避けるためにヒーター エレメントとアンテナ配線に触れないで。
- ワックスが乾燥した後は、アンテナを固定する昆虫ピンを取り外してホルダーからアリを慎重に取り外します。
- 修正アリを保持昆虫ピン モデリング粘土でシリンダーと修正アリの確保、その下にオーダーメイド プラスチックのホルダーが典型的な姿勢を維持し、体全体が自由に移動することを除いて頭。
- トレーニング前に少なくとも 2 時間の暗闇の中でよう注意を払うアリを維持します。
3. トレーニング セットとテスト
-
セットアップ、条件刺激と無条件刺激
- 実験者に対してアクセスを許可する前に開いている白いアクリル ボックス (50 × 50 × 50 cm) で実験を行います。カメラがアリの行動を記録します。
- ボックスの上面の穴から、ant 上に直接マクロ レンズとカメラを配置します。
- 任意の余分な視覚的な手掛かりを減らすためには、アクリル ボックス (2 つの 26 W ライト) の上部を直接指している光源を除いて暗闇の中で部屋を保ちます。
- 視覚的な合図 (CS) として明るい青 (スペクトル図 S1) 段ボール四角形を使用 (60 × 45 mm) 中心に針、注射器 (2 mL) に接続に接続されている、米国 (ショ糖 200 g/L) アリ (図 1 aおよび1B に手動で配信されます).
注: ショ糖液可能任意の白い砂糖で色と水に溶解したときの臭気があるないこと。 - 並行して、ペア、無作為の順序で (以下の説明を参照)、対になっていないトレーニングの 2 種類を行います。
注: この研究では、我々 注意を払っていない、視覚の性質を。手玉と報酬間の関連付けのみが考慮されました。色と形がテストされなかった、他の機能と CS と同様の結果を生成可能性があります。それにもかかわらず、同じ属からアリがカバー青色18の波長に敏感であると示されているので、青い色が選ばれました。
- 実験者に対してアクセスを許可する前に開いている白いアクリル ボックス (50 × 50 × 50 cm) で実験を行います。カメラがアリの行動を記録します。
-
ペアのトレーニング
注: ペアの研修では、CS と米国として記載してすべての試験お互い (図 1) に関連付けられています。- アリが自発的に実行していないことマラー プレゼンテーションの前に確実に CS を提示する前にアリ 10 s を録音を開始します。アリは、この期間中にマラーを行う、数秒のための試験を延期します。任意の ant は、継続的にこの動作を示している場合は、解析から除外します。
- 〜 10 の ant の前に注射器 + CS を移動 s、針の先端を隔てて ant の顔と最大 5 mm 上記のそれぞれの側に。この時間の間に、(図 1 b) アンテナに触れることがなく蟻の頭に針の先端をできるだけ近くに移動します。
注: モーションは、ミツバチ6の視覚の連合学習の役割を果たすに示されているために、CS を提示するとき含まれていた。 - シリンジ先端からショ糖の一滴をしみ出させるし、触角と口器、ショ糖を検出するための ant を許可するの横にあるスクロースのドロップを配置する圧力を適用します。約 5 フィードに ant を許可する s。
注: 各試行で消費されるショ糖アリの量はだったこれらの実験では制御されません。ショ糖をアリにフィードが約 5 自由にフィードを許可するのに十分な大きさにする必要がありますのドロップ s。 - この手順を 10 x 5 分の ITI。
メモ: これらの実験でアクリル ボックスの開放側に近い右目の右には常にアリが電源。したがって、CS は常に、右側からアリを近づいた。けれどもこれは学習を無効にしない場合、これらの実験が可能な側の任意の効果を避けるために、右に左にアリの半 々 にすることで実施できます。
-
対になっていないトレーニング
注: このトレーニングはアリに CS または米国別に献上、されて、これらの 2 つの刺激、したがって、時間 (図 1) をかけてお互いから分離しました。- 各試験前にアリ 10 s の記録を開始します。
- ペアになっているトレーニングと同様の方法で CS を提示、アリにショ糖を提供しません。
- 2.5 分後に、注射器を使用して接続されている CS なし (アンテナ) にも触れる口器に直接米国を提供します。
- CS の体験版で起動し、この手順 10 を繰り返します 2.5 分の ITI とそれらの間をインターカ レート試験の種類ごとに x。
-
テスト
注: テストの最後のトレーニング試験後実施のみ 1 x、どちらか 10 分または 1 時間をする必要があります。CS ではなく、米国にアリが表示されるテストは以前に形成された連想記憶の絶滅を引き起こす可能性があります。- テスト前にアリ 10 s の記録を開始します。
- 約 10 のための ant に CS を提示 s。
- 蟻はショ糖のテストの後に提供することによってフィードにまだやる気を確認します。
4. データの収集と分析
- 上記、10 のアリの行動を記録 s 各試験前に、CS と米国のプレゼンテーション中に。事後分析のためすべての試験とテストが記録されることを確認します。
- トレーニングとテスト中に行われた録音を使用してスコア、10 の間にアリのレスポンス CS プレゼンテーションの s。
- 3 種類の拡張子、口器の動きに応じて動作に CS プレゼンテーション中に別のアリの応答: 完全伸展運動 (有限要素法) と供給、動き (FE) のない完全な拡張や部分的な拡張 (PE) の場合と同様、上顎唇や上顎 palps (図 2 a - 2 D)。分析のため 1 つの応答 (図 2 e) としてすべてのマラーの種類をグループ化します。
- あらゆる試験およびテストのないフィードは ant を除外します。
注: 分析のため個々 の蟻を考慮した統計的検定をお勧め、それにより個人内変動の会計します。バイナリの方法 (肯定応答の 1 と応答なしの場合は 0) で訓練中にアリの反応を分類するとき、混合効果とロジスティック回帰はお勧め16,19です。各訓練試験またはテスト、 Gの応答アリの割合を比較するため-テストまたはフィッシャーの正確なテストが推奨、アリ分析16,20数に応じて。
Representative Results
古典的条件づけの実験中に CS は動物の自発の応答を誘発する必要があります。行った実験でアリのわずか 3%-4% はトレーニングする前に、最初の試験で視覚的な手掛かりに応答マラーを実行しました。ペアの訓練を受けていたアリは、CS に応答ますますマラーを実行 (図 3 a; ロジスティック回帰、 N = 51、自由度 (df) = 507、z = 5.949 p < 0.01)。CS に対応した対アリの割合 50%、3 番目の試験以降から頭打ちになった頃。それどころか、対になっていないアリも有意増加訓練中にマラー (図 3 b; ロジスティック回帰、 N = 29、df = 287、z 0.758、 p = = 0.45)。マラーのビジュアル キューへの応答で発生トレーニング ペアよりもペアになって中に高値を示した (ロジスティック回帰、 N = 80、df = 796、z =-5.306、 p < 0.01)、最初 (表 1) を除くすべての試験の本当だった。
アリがテストされた、短・中期的メモリ15を調べるため、10 分または最後のトレーニング試験後 1 h。(図 3および3 D; のペアではなく、対になっていない訓練を受けていたとき両方のテスト マラーを CS への応答で実行するアリの割合が有意に高かった表 1)。
トレーニング、個々 の蟻の間に数と種類のマラー (図 4 aおよび4 b) 表示されることで実質的な変動を示した。彼らは答えた、彼らはしなかった試験の間ほとんどのアリに交代しながら、対アリの 14% だけは 2 番目または 3 番目の試験以降からすべての試験で答えた。アリが答えたそれら訓練試験の拡張程度彼らの口器を移動が変化。したがって、我々 は 3 種類にマラーを分かれています: 有限要素法、鉄、または PE。通常、アリでは FE が多い以上、有限要素法または PE が実行されています。ただし、いくつかのアリだけ一貫した応答の同じ種類を行うほとんどの場合、アリは、彼ら実行 (図 4) をマラーの種類に少し一貫性を示した。

図 1: 実験のセットアップとトレーニング方式。虫ピンにワックスで ant がモデルに接続されている (A) 修正 (オレンジ) 粘土の円柱と自然主義的なスタンスを許可するそれの下に特注のホルダーの場所。カメラのすぐ下に、2 つの光源に照らされた白いアクリル ボックス内に配置します。無条件刺激 (米国)、砂糖の報酬を提供する注射器に接続されている段ボール (青い四角) を条件刺激 (CS) として使用します。インセットは、ホルダーにアリのクローズ アップ ビューを示しています。(B) 注射器の先端の針は、この回路図で示すように、アンテナに触れることがなくが、蟻の頭にできるだけ近くに移動されます。いずれか、(C) を通じて鉄道アリのペアまたは (D) 対トレーニングになっていません。この図は、フェルナンデスらから変更されています。16.この図の拡大版を表示するのにはここをクリックしてください。

図 2: 木蟻の上顎陰唇拡張応答します。個々 のフレームのビデオ録画からは、トレーニング中にアリの口の部分の動きを表示します。(A) このパネルは応答を示しています。(B) このパネルは、バルセロナの完全な拡張子 (FE) 上顎唇を終了するを示しています。このパネル (C) 目に見える上顎 palpus のみの部分拡張 (PE) を示します。(D) このパネルは、上顎唇構造の部分拡張 (PE) を示しています。(E) アリ対訓練を受けている (N = 51) FE (半ばブラウン) や PE (ライトブラウン) (FEM; ダークブラウン) の動きと完全な拡張子を実行します。この図は、フェルナンデスらから変更されています。16.この図の拡大版を表示するのにはここをクリックしてください。

図 3: 木の砂糖報酬と共に視覚手がかりの連想記憶アリ フォーム。(A) 対アリの割合 (N = 51) 研修を通して増加大幅 CS プレゼンテーションへの応答でマラーを実行します。(B) アリ マラーの実行率大幅に増加しなかった不対トレーニング全体 (N = 29)。アリだったテスト (C) 10 分 (ペア: N = 15; ペア: N = 15) 又は (D) 1 時間 (ペア: N = 15; ペア: N = 14) 最後の訓練試験の後。アリのペアまたは対になっていないトレーニング後 CS への対応の割合は、それぞれ茶色または灰色で表されます。マラーの 3 つのタイプは、暗い (FEM)、媒体 (FE)、光 (PE) トーンで表されます。この図は、フェルナンデスらから変更されています。16.この図の拡大版を表示するのにはここをクリックしてください。

図 4: トレーニング中にアリの個々 のパフォーマンス。これらのパネルは、個々 の公演を表示 (A) の対アリ、(B) 登録がアリ。マラーの 3 つのタイプは、暗い (FEM)、媒体 (FE) とライトブラウン (PE) またはグレーで表されます。この図は、フェルナンデスらから変更されています。16.この図の拡大版を表示するのにはここをクリックしてください。
| 試用版 | N | df | G (調整) | P |
| 1 | 80 | na | na | > 0.1 |
| 2 | 80 | 1 | 3.86 | < 0.05 |
| 3 | 80 | 1 | 8.41 | < 0.01 |
| 4 | 80 | 1 | 6.63 | < 0.01 |
| 5 | 80 | 1 | 8.41 | < 0.01 |
| 6 | 80 | 1 | 7.5 | < 0.01 |
| 7 | 80 | 1 | 10.69 | < 0.01 |
| 8 | 80 | 1 | 11.76 | < 0.01 |
| 9 | 80 | 1 | 17.13 | < 0.01 |
| 10 | 80 | 1 | 17.13 | < 0.01 |
| 10 分 | 59 | 1 | 5.5 | < 0.05 |
| 1 h | 59 | 1 | 4.42 | < 0.05 |
表 1: ペアし、トレーニング、試験および試験ごとに対になっていないマラーを受けていた蟻間 CS レスポンスの比較アリ (N)、自由度 (df)、 G数-独立 (G) とpのテストの値が表示されます。フィッシャーの正確確率検定で最初の試験を行った。このテーブルは、フェルナンデスらから変更されています。16。
Discussion
古典的条件付けは、学習・記憶の研究に最もよく確立されたパラダイムのひとつです。私たちがここで提示されたプロトコルはパラダイムの適応蜂蜜蜂労働者2,3設計され、その結果、マルハナバチなど他のいくつかの種で読み出しとしてあたりを使用しても10 を学習のためのショウジョウバエを使用 ,11とバッタ、アリ、POR とマラー、それぞれ12,13を使用します。このプロトコルを使用して、それは視覚と砂糖報酬間の連合を学び、この短い - (10 分)、中期 (1 時間) の保持を分析するよう注意を払う木蟻を訓練することが可能メモリ16。
行動実験、動物の応答の可変性を最小限に抑えることが重要なステップを考慮する必要が。ここで提示されたプロトコル、いくつかの対策前に、とトレーニング中に変動を最小限に抑えるために取られます。実験開始前に植民地は少なくとも 2 日不足する必要があります、シュガー ドロップ保持ボックスから食べる意欲に基づくアリを選択する必要があります。アリを選択するこの方法では、フィードに意欲のある研修アリのチャンスを最大化するものです。慎重に処理はストレス レベルを低減することができますので、重要な考慮事項でも、中断学習それがあまりにも強烈な21場合。このため、宿泊施設動かずに活かされている、この手順中に (脱出) の任意の動きがストレスの源であるため寒さにアリを麻酔する必要があります。さらに、アリとワックスとの接触は最小限に抑え、アンテナとホット ワックスまたはワイヤは、損傷を引き起こす可能性の間の接触を避ける必要があります。これらの観察が正式に分析されていないがアンテナ学習中に特定のパターンで移動するように見えた。
実験中に砂糖の慎重な配信もアリのモチベーションを維持する重要です。再度、これは正式に分析されているではないながら急激な食品配送の動機と学習不足につながったアリに追加の圧力を引き起こすように見えた。さらに、訓練中に配信されたショ糖は集中力の低下 (200 g/L) の訓練の終わりの前に、の飽食を避けるためにする必要がありますおよびテストします。これは、一緒にこの視覚的応答の低自発的なパフォーマンス、それも飽和しない場合試験を巡るので適して無条件応答にマラーをことができます。最後に、ほとんどの古典的条件付けに反して研究2,3,5,6,7,8,9,10,11,12,13、私たちは訓練 1 つアリ時に実験が終了するまで別の ant をテストするそれを削除するのではなく、試験の間の場所でそれを残してします。ストレスや風景の完全な変更によって引き起こされる視覚の情報間の競合の増加が原因である可能性がありますより変数の結果を生成するように見えたいくつかのアリを一緒にトレーニングします。各実験の期間を減らすためには、我々 はほとんど古典的条件づけ研究16で使用 10 分 ITI の代わりに 5 分 ITI を使用しました。すべてのこれらの考慮事項を助ける必要があります確実にアリをフィードし、トレーニング中に学ぶ動機がいくつかの変動が避けられない。通常の社会、飛び廻る、および歩行の動作を持っているようだアリを使用し、訓練試験やテストにフィードに失敗した瞬間の分析からアリを除くをお勧めします。
本研究では、CS の性質はテストされませんでした。青い視覚刺激を用いて同じ属のアリ18これらの波長に敏感であるのでが、報酬に関連他の色も学習しました可能性があります。それ以上の実験は、完全に見られていると、このセットアップで学んだ色を特徴づける必要があります。これはまた異なった形および視覚手がかりのサイズの場合は true です。アリの空間分解能でそれ発表された蟻の目からの距離でここで紹介する視覚刺激を区別するために十分になる場合我々 はテストしていません。木蟻の複眼は、ファセット22、我々 の知識のサイズと数の面で記載されているが、空間分解能は完全にまだ記載されていません。ただし、これはMelophorus magoti23について計算されています。木蟻の同じような特性、または他のテストの昆虫の目に貢献する視覚の機能の明確な調査観察し、動物によって学んだ。さらに、運動は、古典的条件付け6中にミツバチの連合学習の役割を果たすに示されているために、アリに視覚刺激を提示するとき含まれています。ただし、ミツバチとアリ visual 古典的条件づけの違いはこれもテストは行いませんでしたこの研究では、飛行昆虫の歩行に比べると昆虫の性質上異なる動き観察できます。
最終的なノートでは、アリでは、トレーニング後のような長い期間のために活かされているが存続しなかったので、長期記憶の保持を確認することができませんでした。しかし、実験のそれに続くセットで我々 は生きているアリを保持している、(それらの上にボックスを配置する)、暗くて湿気の多い環境で夜間に食べるし、活かしたし、左を学ぶ動機。したがって、このパラダイムを利用して短、中期記憶に加えて、木蟻の長期記憶の保持が解明される可能性があります。
この簡単な手順は、一般的な古典的条件づけパラダイムから適応で獲得と動物の自由移動を使用してパラダイムの中で広く研究されているよう注意を払うの木蟻で視覚的な記憶の維持を研究することが可能です。このパラダイムには、昆虫のナビゲーションのための非常によく確立されたモデルの学習の神経基盤を分析するために使用される可能性があります。
Disclosures
著者が明らかに何もありません。
Acknowledgments
著者らは、コレクションや木製のアリ ・ コロニーのメンテナンスに関する貴重な情報を共有するためトム コレットとコーネリア ・ Buehlmann をありがとうございます。著者らはまた、この資料の以前のバージョンでコメントをジャスティン クレヴェルを感謝します。
Materials
| Name | Company | Catalog Number | Comments |
| Fluon | Blades Biological Ltd, Edenbridge, UK | ACS 109; ACS 112; ACS 114 | For preventing insects from scaping |
| Crickets | Blades Biological Ltd, Edenbridge, UK | LZJ 217 | Given to the ant colonies as protein source |
| Natural Pine Rosin/Resin | Minerals-water Ltd, Rainham, UK | 500g | Given to the ant colonies for sanitation |
| Austerlitz Insect Pin | Fine Science Tools GmbH, Heidelberg, Germany | 26000-40 | For harnessing ants |
| High speed camera | Edmund Optics Inc., Barrington, USA | eo-13122M | MaLER recordings during training and testing |
| Macrolens | Cannon, Surrey, UK | EF 100 mm f/2.8 L Macro IS USM | MaLER recordings during training and testing |
| Software | IDS Imaging Development Systems GmbH | uEye64 | MaLER recordings during training and testing |
| Blue Cardboard | john smith's at Union Store, University of Sussex | JACK-PJM41358 | Constitutes de conditional stimulus |
| Syringe | Fisher Scientific LTD, Loughborough, UK | BD Plastipak 300185case; Product Code.12369289 | US and CS (attached) presentation |
| Needle (0.5 x 16 mm) | Fisher Scientific LTD, Loughborough, UK | BD Microlance 300600; Product Code:10442204 | US and CS (attached) presentation |
References
- Chittka, L., Niven, J.
- Takeda, K. Classical conditioned response in the honey bee. Journal of Insect Physiology. 6, 168-179 (1961).
- Bitterman, M. E., Menzel, R., Fietz, A., Schäfer, S. Classical conditioning of proboscis extension in honeybees (Apis mellifera). Journal of Comparative Psychology. 97 (2), 107-119 (1983).
- Menzel, R., Muller, U. Learning and memory in honeybees: from behavior to neural substrates. Annual Review of Neuroscience. 19 (1), 379-404 (1996).
- Hori, S., Takeuchi, H., Kubo, T. Associative learning and discrimination of motion cues in the harnessed honeybee Apis mellifera L. Journal of Comparative Physiology A. 193 (8), 825-833 (2007).
- Balamurali, G. S., Somanathan, H., De Ibarra, N. H. Motion cues improve the performance of harnessed bees in a colour learning task. Journal of Comparative Physiology A. 201 (5), 505-511 (2015).
- Mujagić, S., Würth, S. M., Hellbach, S., Dürr, V. Tactile conditioning and movement analysis of antennal sampling strategies in honey bees (Apis mellifera L). Journal of Visualized Experiments. (70), e50179 (2012).
- Wright, G., et al. Parallel reinforcement pathways for conditioned food aversions in the honeybee. Current Biology. 20 (24), 2234-2240 (2010).
- Vergoz, V., Roussel, E., Sandoz, J. C., Giurfa, M. Aversive learning in honeybees revealed by the olfactory conditioning of the sting extension reflex. PloS One. 2 (3), e288 (2007).
- Laloi, D., et al. Olfactory conditioning of the proboscis extension in bumble bees. Entomologia Experimentalis et Applicata. 90 (2), 123-129 (1999).
- Chabaud, M. A., Devaud, J. M., Pham-Delègue, M. H., Preat, T., Kaiser, L. Olfactory conditioning of proboscis activity in Drosophila melanogaster. Journal of Comparative Physiology A. 192 (12), 1335-1348 (2006).
- Simões, P., Ott, S. R., Niven, J. E. Associative olfactory learning in the desert locust, Schistocerca gregaria. Journal of Experimental Biology. 214 (15), 2495-2503 (2011).
- Guerrieri, F. J., d'Ettorre, P. Associative learning in ants: conditioning of the maxilla-labium extension response in Camponotus aethiops. Journal of Insect Physiology. 56 (1), 88-92 (2010).
- Simões, P. M., Niven, J. E., Ott, S. R. Phenotypic transformation affects associative learning in the desert locust. Current Biology. 23 (23), 2407-2412 (2013).
- Guerrieri, F. J., d'Ettorre, P., Devaud, J. M., Giurfa, M. Long-term olfactory memories are stabilised via protein synthesis in Camponotus fellah ants. Journal of Experimental Biology. 214 (19), 3300-3304 (2011).
- Fernandes, A. S. D., Buckley, C. L., Niven, J. E. Visual associative learning in wood ants. Journal of Experimental Biology. 221 (3), 173260 (2018).
- Christe, P., Oppliger, A., Bancala, F., Castella, G., Chapuisat, M. Evidence for collective medication in ants. Ecology Letters. 6 (1), 19-22 (2003).
- Aksoy, V., Camlitepe, Y. Behavioural analysis of chromatic and achromatic vision in the ant Formica cunicularia (Hymenoptera: Formicidae). Vision research. 67, 28-36 (2012).
- Bates, D. M. lme4: Mixed-Effects Modeling with R. , Springer. New York, NY. (2010).
- Sokal, R. R., Rohlf, F. J. Biometry (3rd edn). , W.H. Freman and Company. New York, NY. (1995).
- Bateson, M., Desire, S., Gartside, S. E., Wright, G. A. Agitated honeybees exhibit pessimistic cognitive biases. Current Biology. 21 (12), 1070-1073 (2011).
- Perl, C. D., Niven, J. E. Differential scaling within an insect compound eye. Biology letters. 12 (3), 20160042 (2016).
- Schwarz, S., Narendra, A., Zeil, J. The properties of the visual system in the Australian desert ant Melophorus bagoti. Arthropod Structure & Development. 40 (2), 128-134 (2011).
Tags
動作、問題 140、視覚的な手掛かり、飛び廻るエアコン、学習、記憶、マラー、フォーマイカ西隣Erratum
Formal Correction: Erratum: Visual Classical Conditioning in Wood Ants
Posted by JoVE Editors on 12/31/1969.
Citeable Link.
An erratum was issued for: Visual Classical Conditioning in Wood Ants. The Acknowledgments section was updated, and a supplemental figure was added.
The Acknowledgments sections was updated from:
The authors thank Tom Collett and Cornelia Buehlmann for sharing valuable information regarding collection and maintenance of wood ant colonies. The authors also thank Justine Crevel for commenting on previous versions of this article.
to:
The authors thank Tom Collett and Cornelia Buehlmann for sharing information regarding collection and maintenance of wood ant colonies. The authors also thank Justine Crevel for commenting on previous versions of this article, and Nora Nevala for measuring the spectrum intensity of the visual stimulus. This work was supported by a BBSRC grant to JEN (grant number BB/R005036/1). All the data pertaining to this manuscript are published in the University of Sussex Research Data Repository online database (10.25377/sussex.5794386).
The following supplemental figure was added to the end of the Representative Results section:
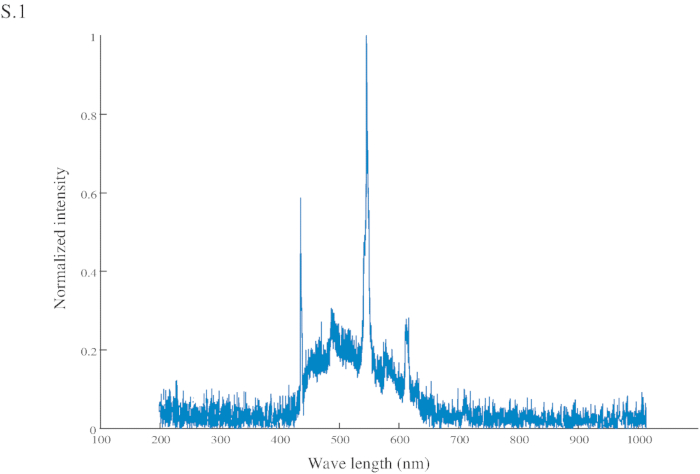
Figure S1: Normalized intensity of the conditional stimuli (CS). The CS has a peak intensity at 545 nm (in the green range) and another at 435 nm (in the blue range).
