

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Diese Protokolle werden den Benutzern helfen, den mitochondrialen Energiestoffwechsel in 3D-Krebszelllinien-abgeleiteten Sphäroiden mithilfe der extrazellulären Flussanalyse von Seepferdchen zu untersuchen.
Abstract
Dreidimensionale (3D) zelluläre Aggregate, sogenannte Sphäroide, sind in den letzten Jahren an die Spitze der In-vitro-Zellkultur gerückt. Im Gegensatz zur Kultivierung von Zellen als zweidimensionale, einzellige Monoschichten (2D-Kultur) fördert, reguliert und unterstützt die sphäroide Zellkultur die physiologische Zellarchitektur und -merkmale, die in vivo existieren, einschließlich der Expression extrazellulärer Matrixproteine, Zellsignalisierung, Genexpression, Proteinproduktion, Differenzierung und Proliferation. Die Bedeutung der 3D-Kultur wurde in vielen Forschungsbereichen anerkannt, darunter Onkologie, Diabetes, Stammzellbiologie und Tissue Engineering. In den letzten zehn Jahren wurden verbesserte Methoden entwickelt, um Sphäroide herzustellen und ihre Stoffwechselfunktion und ihr Schicksal zu beurteilen.
Extrazelluläre Flussanalysatoren (XF) wurden verwendet, um die mitochondriale Funktion in 3D-Mikrogeweben wie Sphäroiden zu untersuchen, wobei entweder eine XF24-Insel-Capture-Platte oder eine XFe96-Sphäroid-Mikroplatte verwendet wurden. Unterschiedliche Protokolle und die Optimierung des mitochondrialen Energiestoffwechsels in Sphäroiden unter Verwendung der XF-Technologie wurden jedoch nicht im Detail beschrieben. Dieses Dokument enthält detaillierte Protokolle zur Untersuchung des mitochondrialen Energiestoffwechsels in einzelnen 3D-Sphäroiden unter Verwendung von Sphäroid-Mikrotiterplatten mit dem XFe96 XF-Analysator. Unter Verwendung verschiedener Krebszelllinien kann die XF-Technologie nachweislich zwischen der Zellatmung in 3D-Sphäroiden nicht nur unterschiedlicher Größe, sondern auch unterschiedlicher Volumina, Zellnummern, DNA-Gehalt und -Typ unterscheiden.
Die optimalen mitochondrialen Effektorverbindungskonzentrationen von Oligomycin, BAM15, Rotenon und Antimycin A werden verwendet, um spezifische Parameter des mitochondrialen Energiestoffwechsels in 3D-Sphäroiden zu untersuchen. Dieses Papier diskutiert auch Methoden zur Normalisierung von Daten, die von Sphäroiden erhalten wurden, und befasst sich mit vielen Überlegungen, die bei der Erforschung des Sphäroidstoffwechsels mit der XF-Technologie berücksichtigt werden sollten. Dieses Protokoll wird dazu beitragen, die Forschung in fortschrittlichen In-vitro-Sphäroidmodellen voranzutreiben.
Introduction
Fortschritte bei In-vitro-Modellen in der biologischen Forschung haben in den letzten 20 Jahren rasante Fortschritte gemacht. Solche Modelle umfassen nun Organ-on-a-Chip-Modalitäten, Organoide und 3D-Mikrogewebe-Sphäroide, die alle zu einem gemeinsamen Schwerpunkt geworden sind, um die Translation zwischen In-vitro- und In-vivo-Studien zu verbessern. Die Verwendung fortschrittlicher In-vitro-Modelle, insbesondere von Sphäroiden, umfasst mehrere Forschungsbereiche, darunter Tissue Engineering, Stammzellforschung, Krebs und Krankheitsbiologie 1,2,3,4,5,6,7 und Sicherheitstests, einschließlich genetischer Toxikologie 8,9,10, Nanomaterial-Toxikologie 11, 12,13,14 und Arzneimittelsicherheits- und Wirksamkeitsprüfung 8,15,16,17,18,19.
Die normale Zellmorphologie ist entscheidend für den biologischen Phänotyp und die biologische Aktivität. Die Kultivierung von Zellen zu 3D-Mikrogewebe-Sphäroiden ermöglicht es Zellen, eine Morphologie, phänotypische Funktion und Architektur anzunehmen, die eher der in vivo beobachteten ähnelt, aber mit klassischen Monolayer-Zellkulturtechniken schwer einzufangen ist. Sowohl in vivo als auch in vitro wird die zelluläre Funktion direkt von der zellulären Mikroumgebung beeinflusst, die nicht auf die zelluläre Kommunikation und Programmierung beschränkt ist (z. B. Zell-Zell-Verbindungsbildungen, Möglichkeiten zur Bildung von Zellnischen); Zellexposition gegenüber Hormonen und Wachstumsfaktoren in der unmittelbaren Umgebung (z. B. zelluläre Zytokin-Exposition als Teil einer Entzündungsreaktion); Zusammensetzung physikalischer und chemischer Matrizen (z. B. ob Zellen in einer steifen Gewebekulturplastik oder einer elastischen Gewebeumgebung gezüchtet werden); und vor allem, wie der Zellstoffwechsel durch die Ernährung und den Zugang zu Sauerstoff sowie die Verarbeitung von Stoffwechselabfallprodukten wie Milchsäure beeinflusst wird.
Die metabolische Flussanalyse ist eine leistungsstarke Möglichkeit, den Zellstoffwechsel innerhalb definierter In-vitro-Systeme zu untersuchen. Insbesondere ermöglicht die XF-Technologie die Analyse von Live-Echtzeit-Veränderungen in der zellulären Bioenergetik intakter Zellen und Gewebe. Angesichts der Tatsache, dass viele intrazelluläre Stoffwechselereignisse in der Größenordnung von Sekunden bis Minuten auftreten, sind funktionelle Ansätze in Echtzeit von größter Bedeutung, um Echtzeitänderungen des zellulären Stoffwechselflusses in intakten Zellen und Geweben in vitro zu verstehen.
Dieses Papier enthält Protokolle für die Kultivierung von Krebs-abgeleiteten Zelllinien A549 (Lungen-Adenokarzinom), HepG2 / C3A (hepatozelluläres Karzinom), MCF-7 (Brust-Adenokarzinom) und SK-OV-3 (ovarielles Adenokarzinom) als In-vitro-3D-Sphäroidmodelle unter Verwendung von erzwungenen Aggregationsansätzen (Abbildung 1). Es beschreibt auch (i) detailliert, wie der mitochondriale Energiestoffwechsel einzelner 3D-Sphäroide mit dem Agilent XFe96 XF-Analysator untersucht werden kann, (ii) zeigt Möglichkeiten zur Optimierung von XF-Assays mit einzelnen 3D-Sphäroiden auf und (iii) diskutiert wichtige Überlegungen und Einschränkungen der Untersuchung des 3D-Sphäroidstoffwechsels mit diesem Ansatz. Am wichtigsten ist, dass dieses Papier beschreibt, wie Datensätze gesammelt werden, die die Berechnung der Sauerstoffverbrauchsrate (OCR) ermöglichen, um die oxidative Phosphorylierung und damit die mitochondriale Funktion in zellulären Sphäroiden zu bestimmen. Obwohl nicht für dieses Protokoll analysiert, ist die extrazelluläre Versauerungsrate (ECAR) ein weiterer Parameter, der neben OCR-Daten in XF-Experimenten gemessen wird. ECAR wird jedoch oft schlecht oder falsch aus XF-Datensätzen interpretiert. Wir kommentieren die Grenzen der Berechnung von ECAR nach grundlegenden Ansätzen des Technologieherstellers.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

Abbildung 1: Grafischer Workflow zur Erzeugung zellulärer Sphäroide, extrazellulärer Flussanalyse und nachgelagerter Assays. Vier Krebszelllinien wurden selektiv als Monoschichten (A) kultiviert, von Gewebekulturkolben gelöst und in Ultralow-Attachment-96-Well-Mikroplatten gesät, um Sphäroide zu bilden (B). A549-Lungenkarzinom, HepG2 / C3A-Leberkarzinom, SK-OV-3-Ovarialadenokarzinom und MCF-7-Mammakarzinomzellen wurden bei 1 × 10 3-8 × 103 Zellen / gut ausgesät und bis zu 7 Tage gezüchtet, um einzelne Sphäroide zu bilden und die Sphäroidaussaatdichte und Kultivierungszeit durch kontinuierliche Beobachtung und planimetrische Messungen zu optimieren. Nach der Bildung wurden einzelne Sphäroide in ein serumfreies XF-Medium gewaschen und sorgfältig in Sphäroid-Assay-Mikrotiterplatten gesät, die mit Poly-D-Lysin (C) vorbeschichtet waren. Sphäroide wurden einer extrazellulären Flussanalyse mit dem XFe96-Analysator unter Verwendung mehrerer Protokolle unterzogen, um Folgendes zu adressieren: (1) optimale Sphäroidgröße für die basale mitochondriale Atmungsreaktion; (2) optimierte Titration von mitochondrialen Atemhemmern; (3) Optimierung der Sphäroidplatzierung in Mikroplattenvertiefungen. (D) Post-XF-Analysen, Phasenkontrastmikroskopie und Sphäroid-DNA-Quantifizierung wurden für die Datennormalisierung und andere nachgelagerte In-vitro-Assays verwendet. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
1. Kultivierung von Krebszelllinien als 3D-In-vitro-Sphäroide
| Zelllinie | Beschreibung | Nährmedium | Quelle |
| A-549 | Lungenkarzinom-Zelllinie | RPMI 1640 | Europäische Sammlung authentifizierter Zellkulturen (ECACC) |
| Natriumpyruvat (1 mM) | |||
| Penicillin- Streptomycin - (100 U / ml - 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| HepG2/C3A | Leberkarzinom-Zelllinie, ein klonales Derivat der Elternzelllinie HepG2 | DMEM | Amerikanische Gewebekultursammlung (ATCC) |
| Penicillin- Streptomycin - (100 U / ml - 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| MCF7 | Brust-Adenokarzinom-Zelllinie | RPMI 1640 | Europäische Sammlung authentifizierter Zellkulturen (ECACC) |
| Natriumpyruvat (1 mM) | |||
| Penicillin- Streptomycin - (100 U / ml - 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| SK-OV-3 | Ovarielle Adenokarzinom-Zelllinie | RPMI 1640 | Europäische Sammlung authentifizierter Zellkulturen (ECACC) |
| Natriumpyruvat (1 mM) | |||
| Penicillin- Streptomycin - (100 U / ml - 100 mg / ml) | |||
| 10 % (v/v) FBS | |||
| Bestandteil | RPMI-Assay-Medium (50 ml Endvolumen) | ||
| Basismedium | Agilent Seepferdchen XF RPMI, pH 7,4 | ||
| Glukose (1 M steriler Vorrat) | 11 mM (0,55 mL Stammlösung) | ||
| L-Glutamin (200 mM steriler Vorrat) | 2 mM (0,5 mL Stammlösung) | ||
| Natriumpyruvat (100 mM steriler Vorrat) | 1 mM (0,5 mL Stammlösung) |
Tabelle 1: Krebszelllinienmedien und XF-Medienzusammensetzungen.
- Kultivieren Sie alle Zelllinien mit der aseptischen Standard-Gewebekulturtechnik und bestätigen Sie, dass sie frei von Mykoplasmen sind, indem Sie ein geeignetes Assay-Kit verwenden.
- Kultivieren Sie die Zelllinien in T75-Gewebekulturflaschen oder einem gleichwertigen Plasma unter Verwendung des empfohlenen Mediums (Tabelle 1). Kultivieren Sie die Zelllinien auf 65-80% Konfluenz und passieren Sie sie regelmäßig bis zu einem Maximum von 25 Passagen.
- Spülen Sie die Zellkulturkolben zweimal in Dulbeccos modifizierter phosphatgepufferter Kochsalzlösung (DBPS) ab.
- Lösen Sie die Zellen aus den Kolben mit 3 ml des Zelldissoziationsreagenzes (siehe Materialtabelle) für 5 min bei 37 °C und bestätigen Sie die Ablösung durch Mikroskopie.
- Saugen Sie die abgelöste Zellsuspension vorsichtig ab, um eine Einzelzellsuspension zu gewährleisten, und deaktivieren Sie das Zelldissoziationsreagenz mit 7 ml vollständigem Gewebekulturmedium.
- Sammeln Sie die Zellen durch Zentrifugation bei 300 × g für 5 min, verwerfen Sie den Überstand und suspendieren Sie die Zellen in vollständigem Medium.
- Zählen Sie die Zellen mit einem Hämozytometer oder einem automatisierten Zellzähler und titrieren Sie auf die gewünschte Zelldichte, die für die Aussaat erforderlich ist.
HINWEIS: Um eine ganze 96-Well-Platte bei 100 μL / Well bei 4 × 103 Zellen / Well zu säen, sollten die Zellen auf 4 × 104 Zellen / ml in einem empfohlenen Volumen von 12 ml titriert werden. - Dekantieren Sie die Zellsuspension in ein steriles Reservoir und geben Sie 100 μL der Zellsuspension mit einem Mehrkanalpipettor in jede Vertiefung einer zellabweisenden Mikrotiterplatte ab.
HINWEIS: Nur die inneren 60 Vertiefungen einer Mikrotiterplatte sollten ausgesät und der Rest mit DPBS gefüllt werden. Dies bildet eine Verdampfungsbarriere, gewährleistet die Sphäroidhomogenität über die Platte und minimiert Plattenrandeffekte. - Zentrifugieren Sie Sphäroid-Mikrotiterplatten bei 300 × g für 15 Minuten, um die Zellen in lose Aggregate zu zwingen.
- Inkubieren Sie die Platten bei 37 ° C, 5% CO2 für mindestens 3 Tage, um die Sphäroidbildung sicherzustellen.
- Führen Sie eine Phasenkontrastmikroskopie mit standardisierten Laborpraktiken durch, um das Wachstum von Sphäroiden zu überwachen. Füllen Sie das Zellkulturmedium alle 3 Tage oder zweimal wöchentlich auf, indem Sie einen halbvolumigen Medienaustausch durchführen.
2. Untersuchung des mitochondrialen Energiestoffwechsels einzelner Sphäroide mit Hilfe der extrazellulären Flussmitteltechnologie (XF)
- Assay-Vorbereitung (einen Tag vorher)
- Überprüfen Sie die Sphäroidlebensfähigkeit mit einem inversen Lichtmikroskop mit Phasenkontrast bei 4-facher Vergrößerung, um eine intakte Sphäroidstruktur, Morphologie und allgemeine Gleichmäßigkeit zwischen den Proben zu gewährleisten.
- Hydratisieren Sie die Sensorpatrone.
- Aliquot ~ 20 ml des Kalibers in ein konisches Rohr.
- Das konische Rohr, das das Kalibrant enthält, wird über Nacht in einen Nicht-CO2 37 °C-Inkubator gegeben.
- Entfernen Sie den Inhalt aus dem Assay-Kit.
- Entfernen Sie die Sensorkassette von der Gebrauchsplatte und legen Sie sie kopfüber auf die Arbeitsplatte neben der Stromplatte.
- Pipette 200 μL steriles ddH2O in jede Vertiefung der Sensorkartuschen-Utility-Platte mit einer Mehrkanal-P300-Pipette.
- Legen Sie die Sensorkassette auf die Utility-Platte.
- Überprüfen Sie, ob der Wasserstand in jedem Brunnen hoch genug ist, um die Sensorsonden einzutauchen.
- Die montierte Sensorkartusche in einen CO2-freien 37 °C-Inkubator geben und über Nacht stehen lassen.
HINWEIS: Dieser Schritt kann 12-72 h vor Beginn des Assays durchgeführt werden.
- Coat Sphäroid Assay Mikrotiterplatte
- Mit aseptischen Techniken 30 μL/Well sterile Poly-D-Lysin-Lösung (0,1 mg/ml) in die Sphäroid-Mikroplatte geben und 30 min bei Raumtemperatur inkubieren.
- Saugen Sie die Lösung aus jeder Vertiefung der Sphäroid-Mikrotiterplatte ab, invertieren Sie die Platte und klopfen Sie sie fest auf Seidenpapier, um die Restlösung zu entfernen.
- Waschen Sie die Platte zweimal mit 200 μL/Well von sterilem ddH2O.
- Kehren Sie nach der letzten Wäsche die Mikrotiterplatte um und klopfen Sie sie fest auf Seidenpapier, um Restwasser zu entfernen.
- Lassen Sie die Platte 30 Minuten an der Luft trocknen, bevor Sie sie verwenden oder bei 4 ° C für die zukünftige Verwendung lagern.
HINWEIS: Die Sphäroid-Assay-Mikrotiterplatte sollte mit einem molekularen Klebstoff beschichtet werden, um sicherzustellen, dass die Sphäroide am Boden der Mikrotiterplatte fixiert sind. Ohne einen molekularen Klebstoff können sich Sphäroide lösen und die Assay-Ergebnisse beeinträchtigen. Andere molekulare Klebstoffe können auch als Alternative zu Poly-D-Lysin für die Vorbeschichtung von Platten verwendet werden. Vorbeschichtete Platten können bei 4 °C gelagert werden, sollten aber vor Beginn des Assays auf Raumtemperatur ausgeglichen werden.
- XF Assay-Medium vorbereiten
- Vorbereiten von XF RPMI-Medium, wie in Tabelle 1 beschrieben, und Sterilfilter mit einem 0,22-μm-Spritzenfilter
- Assay-Vorbereitung (1 h vor dem Assay)
- Das ergänzte XF RPMI Assay medium auf 37 °C vorwärmen.
- Vorwärmen Sie die beschichtete Sphäroid-Assay-Mikroplatte in einem Nicht-CO2 37 °C-Inkubator oder Trockenbad.
- Bereiten Sie die Sensorkassette vor.
- Nehmen Sie das konische Rohr mit dem Kalibrant und der Sensorpatrone aus dem Luftinkubator.
- Entfernen Sie die Sensorkassette von der Gebrauchsplatte und legen Sie sie kopfüber auf die Arbeitsfläche.
- Saugen Sie mit einer P300-Mehrkanalpipette das Wasser von der Utility-Platte ab und entsorgen Sie es.
- Gießen Sie die Kalibrantlösung in ein steriles Reagenzreservoir und geben Sie 200 μL/Vertiefung des vorgewärmten Calibrats mit einer P300-Mehrkanalpipette auf die Gebrauchsplatte.
- Nehmen Sie die Sensorpatrone auf und legen Sie sie wieder auf die Gebrauchsplatte, um sicherzustellen, dass die Sensoren gut in das Kaliber eingetaucht sind.
- Bringen Sie die montierte Sensorkassette zurück in den Nicht-CO2 37 °C-Inkubator, bis sie bereit ist, die Anschlussinjektionslösungen zu laden.
- Waschen Sie die Sphäroide mit dem Assay-Medium.
- Entfernen Sie die Sphäroidkulturplatte aus dem 37 ° C, 5% CO2 -Inkubator und beobachten Sie die Sphäroide unter dem Mikroskop, um ihre Integrität vor den Sphäroidtransferschritten sicherzustellen.
- Laden Sie alle Vertiefungen der Sphäroidplatte mit 180 μL/Vertiefung des vorgewärmten Assay-Mediums, einschließlich aller Hintergrundkorrekturquellen.
- Füllen Sie eine 7 cm große Petrischale teilweise mit 3 ml des Assay-Mediums.
- Mit einer Mehrkanalpipette, die mit breiten Pipettenspitzen beladen ist, werden die Sphäroide von der 96-Well-Kulturplatte in 7 cm große Petrischalen übertragen, indem der Pipetter auf ein Aspirationsvolumen von 10-50 μL eingestellt wird.
- Samen Sie Sphäroide in die vorbeschichtete Sphäroid-Assay-Mikroplatte.
- Übertragen Sie die Sphäroide mit einem Dissektionsmikroskop und einer Lightbox-Vorrichtung von der Petrischale auf die Sphäroid-Assay-Mikroplatte, wie unten beschrieben.
- Stellen Sie das Volumen eines Einkanalpipettors mit einer breiten Pipettenspitze auf 20 μL ein und saugen Sie vorsichtig ein einzelnes Sphäroid ab. Platzieren Sie die Spitze direkt in der Mitte jeder Vertiefung der Sphäroid-Assay-Mikroplatte und lassen Sie die Schwerkraft ein einzelnes Sphäroid in die Mitte jedes Vertiefungslochs eluieren, d.h. stoßen Sie kein Medium aus der Pipettenspitze aus und lassen Sie die Kapillarwirkung das Sphäroid aus der Pipettenspitze entfernen. Um die Elution zu bestätigen, kann der Inhalt des Pipettierers unter dem Mikroskop zurück in die 7 cm große Petrischale pipettiert werden.
HINWEIS: Die Schwerkraftelution eines einzelnen Sphäroids dauert typischerweise 15-30 s, abhängig von der Größe / Dichte des Sphäroids. Während dieser Zeit sollte der Pipetter nicht entfernt werden. Alle Hintergrundkorrekturvertiefungen sollten frei von Sphäroiden sein und nur Assay-Medium enthalten. Bestätigen Sie unter dem Mikroskop die Position jedes Sphäroids. Jedes Sphäroid sollte idealerweise in der Mitte jedes Brunnens positioniert werden. - Sobald alle Sphäroide auf die Sphäroid-Assay-Mikroplatte übertragen wurden, wird die Platte vor dem Assay mindestens 1 h lang bei 37 °C in einen Nicht-CO2-Inkubator überführt.
- Stellen Sie das Volumen eines Einkanalpipettors mit einer breiten Pipettenspitze auf 20 μL ein und saugen Sie vorsichtig ein einzelnes Sphäroid ab. Platzieren Sie die Spitze direkt in der Mitte jeder Vertiefung der Sphäroid-Assay-Mikroplatte und lassen Sie die Schwerkraft ein einzelnes Sphäroid in die Mitte jedes Vertiefungslochs eluieren, d.h. stoßen Sie kein Medium aus der Pipettenspitze aus und lassen Sie die Kapillarwirkung das Sphäroid aus der Pipettenspitze entfernen. Um die Elution zu bestätigen, kann der Inhalt des Pipettierers unter dem Mikroskop zurück in die 7 cm große Petrischale pipettiert werden.
- Übertragen Sie die Sphäroide mit einem Dissektionsmikroskop und einer Lightbox-Vorrichtung von der Petrischale auf die Sphäroid-Assay-Mikroplatte, wie unten beschrieben.
3. Aufbereitung und Laden von Verbindungen in die Sensorkartusche für XF-Assays
| Injektionsstrategie | Verbindung (Port) | XFe96 Mikrowellen-Startvolumen (μL) | Gewünschte Endbohrlochkonzentration | Anschlussvolumen (μL) | Endgültiges XFe96-Mikrowellenvolumen nach der Injektion (μL) | Konzentration des Arbeitsmaterials |
| 1 | Oligomycin (A) | 180 | 3 μg/ml | 20 | 200 | 30 μg/ml |
| Rotenon (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimycin A (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 2 | BAM15 (A) | 180 | 5 μM | 20 | 200 | 50 μM |
| Rotenon (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimycin A (B) | 200 | 2 μM | 20 | 220 | 22 μM |
Tabelle 2: Mitochondriale Verbindungskonzentrationen zur Untersuchung des mitochondrialen Energiestoffwechsels einzelner 3D-Sphäroide mit dem XFe96 Analyzer.
- Bereiten Sie die Arbeitsstoffkonzentrationen jeder Verbindung gemäß Tabelle 2 unter Verwendung eines vollständig ergänzten, vorgewärmten XF-RPMI-Assay-Mediums vor.
- Richten Sie die Patronenplatte (gekoppelt an die Utility-Platte) spaltenweise aus, 1-12 von links nach rechts.
- Wenn Sie eine Ladeführung verwenden, legen Sie sie gemäß dem Well-Loading-Verfahren auf die Patronenplatte, z. B. wenn Port A zuerst geladen wird, stellen Sie sicher, dass A in der oberen linken Ecke der Führung sichtbar ist.
- Die Arbeitslösung jeder Verbindung in ein geeignetes Reservoir geben und mit einer kalibrierten P100-Mehrkanalpipette 20 μL in alle entsprechenden Ports dosieren. Wiederholen Sie den Vorgang für jede Verbindung in die verbleibenden Ports.
HINWEIS: Wenn keine Anschlüsse an der Sensorpatronenplatte verwendet werden, können diese leer gelassen oder mit Assay-Medium gefüllt werden. Wenn nur eine Auswahl eines bestimmten Portbuchstabens verwendet wird, stellen Sie sicher, dass die anderen Ports, die diesem Buchstaben entsprechen, mit Assay-Medium geladen sind. Andernfalls wird Luft in das Bohrloch injiziert, wodurch die Ergebnisse in diesen Bohrlöchern beeinträchtigt werden. - Entfernen Sie nach dem Laden des Anschlusses die Plattenladeführungen (falls verwendet) und bereiten Sie den Analysator für das Laden der Sensorpatrone vor.
HINWEIS: Wenn der Assay nicht sofort nach dem Laden der Anschlüsse durchgeführt wird, setzen Sie den Deckel wieder auf die Sensorkassette und legen Sie die Platte wieder in den 37 °C Luftinkubator, bis sie bereit ist, in die Maschine zu laden.
4. Assay-Design, Injektionsstrategien und Datenerfassung
- Ausführen des Assays
- Schalten Sie den Analysator ein und stellen Sie eine Verbindung zum Controller (Computer) her.
HINWEIS: Dies kann durch den Geräteverbindungsstatus im Widget-Panel der Wave Controller-Software überprüft werden. - Navigieren Sie zur Vorlagenseite in der WAVE-Software, suchen Sie die Assay-Vorlagendatei für das Experiment und doppelklicken Sie, um sie zu öffnen.
HINWEIS: Wenn die Assay-Vorlage nicht in der Vorlagenansicht angezeigt wird, importieren Sie die Vorlagendatei von einem freigegebenen Netzlaufwerk oder USB-Flashlaufwerk in den Vorlagenordner . - Um den Assay zu starten, klicken Sie auf die Registerkarte Assay ausführen .
HINWEIS: Wenn die Gruppendefinitionen innerhalb der Plattenkarte korrekt zugeordnet wurden, kann der Assay ausgeführt werden, wie durch das grüne Häkchen auf der rechten Seite der Seite angezeigt. Zu diesem Zeitpunkt können alle zusätzlichen Informationen auf der Seite mit der Zusammenfassung des Assays oder auf der leeren Seite eingegeben werden. Fahren Sie mit dem nächsten Schritt fort. Aufgrund der verzögerten Penetration von mitochondrialen Modulatoren in 3D-Mikrogewebe-Sphäroide (Abbildung 2) sind die in Tabelle 3 beschriebenen Messprotokollinformationen zu verwenden.
- Schalten Sie den Analysator ein und stellen Sie eine Verbindung zum Controller (Computer) her.
| Messzeitraum | Einspritznummer und Port | Details zur Messung | Periodendauer (h:min:s) |
| Kalibrierung | Nicht zutreffend | XF-Analysatoren führen diese Kalibrierung immer durch, um sicherzustellen, dass die Messungen genau sind | 00:20:00 Uhr (dies ist ein Durchschnitt und kann zwischen den Maschinen variieren) |
| Equillibration | Nicht zutreffend | Das Gleichgewicht tritt nach der Kalibrierung auf und wird empfohlen. | 00:10:00 |
| Basal | Nicht zutreffend | Zyklen = 5 | 00:30:00 |
| Mix = 3:00 | |||
| Wartezeit = 0:00 | |||
| Maß = 3:00 | |||
| Oligomycin / BAM15 | Einspritzung 1 (Anschluss A) | Zyklen = 10 | 01:00:00 |
| Mix = 3:00 | |||
| Wartezeit = 0:00 | |||
| Maß = 3:00 | |||
| Rotenon + Antimycin A | Einspritzung 2 (Anschluss B) | Zyklen = 10 | 01:00:00 |
| Mix = 3:00 | |||
| Wartezeit = 0:00 | |||
| Maß = 3:00 | |||
| Gesamtzeit: | 03:00:00 | ||
Tabelle 3: Protokollaufbau zur Untersuchung des mitochondrialen Energiestoffwechsels einzelner 3D-Sphäroide mit dem XFe96 Analyzer.
- Klicken Sie auf Ausführen starten , um das Dialogfeld Speicherort aufzurufen.
- Geben Sie den Speicherort für die Ergebnisdatei ein, und legen Sie die montierte Sensorkassette auf das Thermofach, das an der Tür an der Seite des Analysators angezeigt wird. Warten Sie, bis sich das Thermofach automatisch öffnet und auf dem Bildschirm die Meldung Load Calibrant Utility Plate angezeigt wird. Bevor Sie den Anweisungen auf dem Bildschirm folgen, stellen Sie sicher, dass i) die Sensorpatrone ordnungsgemäß auf der Utility-Platte sitzt, ii) der Deckel von der Sensorpatrone entfernt wird und iii) die Sensorpatrone auf der Utility-Platte richtig ausgerichtet ist.
- Befolgen Sie die Anweisungen auf dem Bildschirm, um die Kalibrierung der Sensorpatrone zu starten.
HINWEIS: Die Zeit bis zum Abschluss der Kalibrierung beträgt ca. 10-20 min (bei Assays bei 37 °C). - Laden Sie nach der Kalibrierung der Sensorkartusche die Sphäroid-Mikroplatte in den Analysator, indem Sie den Anweisungen auf dem Bildschirm auf dem Wave Controller folgen, um den 12-minütigen Gleichgewichtsschritt einzuleiten.
HINWEIS: Grüne Kästchen mit weißen Häkchen zeigen eine "gute" Kalibrierung für diesen Brunnen an. Wenn Bohrlöcher keine "gute" Kalibrierung bieten, werden sie mit einem roten Kasten und einem weißen Kreuz gekennzeichnet. Solche Vertiefungen sollten notiert und von jeder Analyse ausgeschlossen werden, nachdem der Assay mit der Registerkarte Modifikationsassay abgeschlossen wurde. - Warten Sie, bis der Analysator automatisch mit der Erfassung der Basismessungen beginnt, nachdem die Maschine den Gleichgewichtsschritt abgeschlossen hat (wie im Geräteprotokoll beschrieben).
- Um das Experiment abzuschließen, folgen Sie den Bildschirmbefehlen auf dem WAVE-Controller.
HINWEIS: Sobald die Sphäroid-Mikroplatte aus dem Analysator entfernt wurde, entsorgen Sie die Sensorpatrone und legen Sie die Sphäroidplatte für weitere Analysen beiseite, falls erforderlich (z. B. doppelsträngige (ds) DNA-Quantifizierung). Wenn die Mikrotiterplatte nicht für die weitere Analyse benötigt wird, kann sie zusammen mit der Sensorpatrone entsorgt werden. - Warten Sie, bis das Assay-Dialogfeld angezeigt wird, und zeigen Sie die Ergebnisse an oder kehren Sie zur Vorlagenansicht zurück.
5. Datennormalisierungs- und Analysestrategien - Post-Assay-Normalisierung und Downstream-Assays (optionale Schritte)
- Datennormalisierung
- Um Sphäroiddaten zu normalisieren, beziehen Sie sich auf die Reihe von Protokollen, die für Datennormalisierungsstrategien zur Berechnung der Sphäroidgröße und des Sphäroidvolumens und zur Quantifizierung von dsDNA in Sphäroidassays relevant sind. Diese wurden als ergänzende Dateien aufgenommen; siehe Zusatzdatei 1 und Ergänzungsdatei 2.
- Datenanalyse
- Um Daten in einen der automatisierten Analysegeneratoren zu exportieren, folgen Sie den Datenexportbefehlen auf dem WAVE-Controller und wählen Sie den Exportgenerator aus, der dem Assay-Typ entspricht. Alternativ können Sie die Datendatei exportieren und in Seahorse Analytics hochladen.
HINWEIS: Der Nachteil von Berichtsgeneratoren und Seahorse-Analysen besteht darin, dass die Datenanalyse auf die Art und Weise beschränkt ist, wie der XF-Assay konzipiert ist, und keine Mittelwerte über Messzyklen hinweg zulässt. Der manuelle Export von Datensätzen aus der Gerätesoftware ermöglicht diesbezüglich eine Benutzerpräferenz. Angesichts der Tatsache, dass sich die Injektionsstrategie zur Beurteilung der mitochondrialen Atmung von 3D-Sphäroiden wahrscheinlich von der eines typischen "MitoStress" -Tests unterscheidet, wurde eine Reihe von Tabellenvorlagen entwickelt, um die Analyse dieser Datensätze zu unterstützen, die für 3D-Zellkulturen spezifisch sind und auf Anfrage zur Verfügung gestellt werden. Diese Datenvorlagendateien liefern Daten zu den wichtigsten mitochondrialen Atmungsparametern, die in Abbildung 2 detailliert beschrieben und erläutert werden. - Um die Daten zu analysieren, exportieren Sie die Daten als Tabellenkalkulationsbericht aus der WAVE-Controller-Software und verwenden Sie eine unabhängige Tabellenkalkulationsvorlage für die Analyse.
- Um Daten in einen der automatisierten Analysegeneratoren zu exportieren, folgen Sie den Datenexportbefehlen auf dem WAVE-Controller und wählen Sie den Exportgenerator aus, der dem Assay-Typ entspricht. Alternativ können Sie die Datendatei exportieren und in Seahorse Analytics hochladen.

Abbildung 2: Schematische Deskriptoren für Parameter, die aus extrazellulären Flussdatenanalysen abgeleitet wurden. Abkürzung: OCR = Sauerstoffverbrauchsrate. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Um wohlgeformte, kompakte Sphäroide zu erhalten, wurde jede Zelllinie einzeln für die Aussaatdichte und die Kultivierungsdauer optimiert (Abbildung 3). A549-, HepG2/C3A- und SK-OV-3-Zelllinien bildeten zunächst lose Aggregate, die erst nach 7 Tagen in Kultur zu runden Sphäroiden mit klar definierten Umfängen übergingen. Umgekehrt könnten MCF-7-Zellen innerhalb von 3 Tagen Sphäroide bilden. Es gab eine klare Korrelation zwischen der anfänglichen Zellaussaatdichte und dem Sphäroidvolumen nach der Kulturperiode für alle Sphäroidmodelle. Die Größe und Morphologie der Sphäroide wurden auf die Aussaatdichte optimiert. Morphologie und Zirkularität begannen mit zunehmender Sphäroidgröße in allen Modellen abzunehmen. Seeding-Strategien für Zelllinien wurden bei 4 ×10 3 Zellen/Well für A549 und SK-OV-3 Zellen optimiert; HepG2/C3A-Zellen wurden zuvor an anderer Stelle auf 1 × 10 3 Zellen/Well optimiert, und MCF-7-Zellen wurden bei 4 × 103 Zellen/Well in allen Assays verwendet. Bei optimierten Seeding-Strategien lag das Sphäroidvolumen zwischen 5,46 × 107 μm3 (SK-OV-3) und 1,45 × 108 μm3 (A549) (Abbildung 3B). Alle Sphäroidtypen hatten eine lineare Korrelation zwischen der anfänglichen Aussaatdichte und dem Sphäroidvolumen, wobei A549 und HepG2/C3AR2-Werte von 0,957 bzw. 0,947 aufwiesen. Es wurde festgestellt, dass die Sphäroidvolumina von MCF-7 und SK-OV-3 eine größere Korrelation mit der anfänglichen Aussaatdichte R2=0,977 aufweisen (Abbildung 3A).
Die Sphäroidzirkularität wurde mithilfe der Bildplanimetrie innerhalb der FIJI-Analysesoftware unter Verwendung der langen und kurzen Sphäroiddurchmesser berechnet. Die perfekte Sphäroidsymmetrie hatte eine Zirkularität = 1,0; Die Abweichung von 1,0 deutete auf einen Verlust der Zirkularität hin (Abbildung 3C). Die Zirkularität war bei MCF-7-Sphäroiden größer als bei anderen Modellen, bei denen die Zirkularität bei allen Seeding-Dichten zwischen 0,83 und 0,9 gehalten wurde. Im Vergleich dazu war der äußere Umfang der SK-OV-3-Sphäroide nicht so klar definiert, und das Sphäroidvolumen war auch nach 7 Tagen in Kultur signifikant kleiner, was zu Sphäroiden mit einer maximalen Zirkularität von 0,61 bei einer Aussaatdichte von 4 × 103 / Well führte. Es wurde auch festgestellt, dass HepG2 / C3A-Zellen enge, wohlgeformte Sphäroide mit einer gleichmäßigen Morphologie über die Oberfläche aller Sphäroide bilden, wobei die Zirkularität bei 0,79 für Zellen beibehalten wird, die bei 1 × 103 Zellen / Well ausgesät sind. A549-Zellen schienen einem Trend zu folgen, bei dem die sphäroide Zirkularität und Morphologie mit der Aussaatdichte verbessert wurden; Die Zirkularität war jedoch bei der in diesen Experimenten verwendeten Dichte nicht größer als 0,63.
Die basale mitochondriale Atmung wurde als OCR berechnet, die aus Sphäroiden gemessen wurde, die bei 1 × 10 3, 2 × 10 3, 4 × 10 3 oder 8 × 10 3 Zellen / Well in Ultralow Attachment Sphäroidkultur-Mikroplatten ausgesät waren (Abbildung 3D). Für alle Sphäroidtypen stieg die OCR mit der Sphäroidgröße an und korrelierte linear mit dem Sphäroidvolumen, wobei R2 bei MCF-7-Sphäroiden bei 0,988 am höchsten und bei SK-OV-3-Sphäroiden bei 0,744 am niedrigsten war (Abbildung 3E). Die gemessene OCR unterschied sich statistisch zwischen allen experimentellen Gruppen. A549 hatte die niedrigste OCR und erreichte nur 18 pmol/min/well bei der größten Sphäroidgröße (Abbildung 3D). Umgekehrt ergaben MCF-7-Sphäroide nach nur 3 Tagen in Kultur eine ähnliche OCR bei der kleinsten Sphäroidgröße und erreichten eine maximale Ausgangs-OCR von 53 pmol/min/well für die größte Sphäroidgröße (Abbildung 3D). HepG2/C3A lieferte OCR-Daten, die in hohem Maße mit der Größe und Morphologie des Sphäroids übereinstimmen. In HepG2/C3A-Sphäroiden, die aus 1 × 103 Zellen/Well gesät wurden, erreichte die Baseline-OCR einen Durchschnitt von 15 pmol/min/well und stieg in den größten Sphäroiden auf maximal 52 pmol/min/well an (Abbildung 3D). OCR in SK-OV-3-Sphäroiden war nur zwischen Sphäroiden signifikant, die aus 4 × 10 3 Zellen / Well und 8 × 10 3 Zellen / Well gezüchtet wurden, wobei bei Sphäroiden zwischen 1 × 10 3, 2 ×10 3 oder 4 × 10 3 Zellen kaum Unterschiede zu beobachten waren. Trotz Größenunterschieden waren die OCR-Daten zwischen HepG2/C3A- und MCF-7-Sphäroiden an allen Größenpunkten sehr ähnlich. Im Verhältnis zur Sphäroidgröße (μm3) war die Baseline-OCR durch MCF-7-Sphäroide vergleichbar mit der von HepG2/C3A-Sphäroiden, die über 7 Tage aus 1.000 Zellen pro Bohrloch gezüchtet wurden.

Abbildung 3: Sphäroidwachstumsparameter bestimmen die mitochondriale Atmung zu Studienbeginn. (A) A549-, HepG2/C3A-, MCF-7- und SK-OV-3-Sphäroide wurden für die Aussaatdichte optimiert und ihr Wachstum bei 1 × 10 3, 2 × 10 3, 4 × 10 3 und 8 ×10 3 Zellen / Well in jeder der Mikroaufnahmen von oben links nach unten rechts überwacht; Maßstabsstäbe = 500 μm. (B) Die Sphäroidgröße wurde unter Verwendung planimetrischer Daten aus gesammelten Mikroaufnahmen berechnet und unter Verwendung der Korrelationsstatistik von Pearson verglichen. Gepunktete Linien stellen die Verteilung des 95%-Konfidenzintervalls dar. (C) Die Sphäroidmorphologie wurde durch Berechnung der Zirkularität verglichen. (D) Die OCR wurde 5x gemessen, woraufhin Rotenon-Antimycin A hinzugefügt wurde, um die nicht-mitochondriale Atemfrequenz mit dem Agilent Seahorse XFe96-Analysator zu berücksichtigen. OCRbasal gemessen als OCR - OCRr/a wurde zwischen Aussaatdichte (D) und Sphäroidvolumen (E) verglichen. Die Daten sind Durchschnittswerte ± SEM von 5-8 Well-Replikaten pro Sphäroidtyp und Zellaussaatdichte. Abkürzungen: OCR = Sauerstoffverbrauchsrate; OCRbasal = Basale mitochondriale Atmung; OCRr/a = OCR nach Zugabe von Rotenon-Antimycin A. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Der Konzentrations- und Zeitverlauf für die Exposition gegenüber respiratorischen Modulatoren in der XF-Analyse ist ein kritischer Schritt in der Assay-Optimierung. Atemmodulatorverbindungen - Oligomycin, BAM15, eine Mischung aus Rotenon-Antimycin A- oder eine DMSO-Fahrzeugkontrolle - wurden nacheinander über die Einspritzöffnungen der Sensorpatrone in Mikrotiterplattenvertiefungen injiziert, die die MCF-7-Sphäroide enthielten (Abbildung 4A). Vier Messzyklen wurden durchgeführt, um die durchschnittliche basale OCR von 30-40 pmol/min/well für alle Probengruppen zu bestimmen. Für den Rest der Assay-Dauer wurden alle 5 Messzyklen nacheinander Atemmodulatoren zugegeben, um nach Injektion 1 endgültige Bohrlochkonzentrationen von 0,5 μM (0,5 μg/ml Oligomycin) zu erreichen; 2,0 μM (2 μg/ml Oligomycin) nach Injektion 2; 5 μM (5 μg/ml Oligomycin) nach Injektion 3; und schließlich eine maximale Wellkonzentration von 11 μM (11 μg/ml Oligomycin) nach der vierten sequenziellen Addition. MCF-7-Sphäroide reagierten während des gesamten Experiments nicht auf die Fahrzeugsteuerung (Abbildung 4A). Die basale OCR veränderte sich unmittelbar nach der ersten Injektion der jeweiligen Verbindung bei der niedrigsten Konzentration von 0,5 μM oder 0,5 μg/ml Oligomycin (Abbildung 4B). Die OCR in MCF-7-Sphäroiden wurde mit Oligomycin nach 5 Messzyklen nach der ersten Injektion von 0,5 μg/ml von 41 pmol/min/well auf 23 pmol/min/well gesenkt (Abbildung 4B).
Als Reaktion auf 0,5 μm BAM15 wurde die OCR vor der zweiten Injektion von 33 auf 41 pmol/min/gut erhöht (Abbildung 4C). Im Vergleich dazu senkte die Kombination von Rotenon plus Antimycin A die OCR vor der zweiten Injektion von 37 auf 13 pmol/min/gut (Abbildung 4D). Kinetische Spuren zeigten außerdem eine stetige lineare Abnahme (Oligomycin und Rotenon-Antimycin A) oder einen Anstieg (BAM15) der OCR. Für alle Dosierungsschemata von Verbindungen wurde innerhalb von 10-12 vollständigen Messzyklen (60-72 min) eine stationäre OCR bei einer Gesamtmuldenkonzentration von 2 μM BAM15, 2 μM Rotenon, 2 μM Antimycin A und 2 μg/ml Oligomycin erreicht (Abbildung 4A). Die Sauerstoffverbrauchsrate erreichte ein stationäres Plateau bei ~ 19 pmol / min / well (Oligomycin), 52 pmol / min / well (BAM15) und 10 pmol / min / well (Rotenon-Antimycin A) (Abbildung 4A). Eine Erhöhung der zusammengesetzten Konzentration von Oligomycin, BAM15 oder Rotenon plus Antimycin A hatte darüber hinaus keine offensichtliche Wirkung auf die OCR, die während des restlichen Gehalts konstant blieb. Diese Daten zeigen, dass sowohl die Wirkstoffkonzentration als auch der zeitliche Verlauf der Exposition gegenüber Atemmodulatorverbindungen für die Assay-Optimierung bei der Verwendung von 3D-Sphäroiden in Betracht gezogen werden sollten.

Abbildung 4: Titration von respiratorischen Modulatorverbindungen als wichtiger Schritt zur Optimierung der extrazellulären Flussanalyse. (A) MCF-7-Sphäroide wurden an 4 × 10 3 Zellen/Well ausgesät und über 3 Tage kultiviert, bevor sie in Vertiefungen einer Sphäroid-Assay-Mikroplatte, die XF RPMI enthielt, platziert und mit dem XFe96-Analysator auf OCR ± mitochondriale Modulatoren untersucht wurden. OCR wurde 5x gemessen, wonach Titrationen von entweder Vehikelkontrolle, Oligomycin (B), BAM15 (C) oder Rotenon-Antimycin A (D) hinzugefügt wurden, um die mitochondriale ATP-Synthase zu hemmen, die maximale Atmungskapazität zu bestimmen bzw. die nichtmitochondriale Atemfrequenz zu bestimmen. Die Konzentration jedes mitochondrialen Modulators wurde über 4 individuelle Titrationsinjektionsstrategien (0,5 μM, 1,5 μM, 3 μM und 6 μM; Einheiten für Oligomycin sind μg/ml) erhöht, um die maximale stationäre OCR als Reaktion auf die optimale Verbindungskonzentration zu bestimmen. Die OCR wurde für 5 Messzyklen zwischen jeder Injektion gemessen. Daten sind Durchschnittswerte ± SEM von 5-8 einzelnen Bohrlochreplikationen. Abkürzung: OCR = Sauerstoffverbrauchsrate. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Einer der Hauptvorteile der XF-Technologie ist die Möglichkeit, die mitochondriale Funktion in intakten Zellen und Geweben zu untersuchen. Um spezifische Aspekte der mitochondrialen Funktion in Zellen und Geweben zu untersuchen, werden mitochondriale Modulatoren nacheinander über die 4 verfügbaren Injektionsanschlüsse an der Sensorpatrone zu den Vertiefungen der Probenmikrotiterplatte hinzugefügt. Die typische Sequenz von Modulatoren, die zur Untersuchung mitochondrialer Parameter in XF-Assays verwendet werden, sind Oligomycin, ein Protonophor (z. B. FCCP oder BAM15) und eine Kombination aus Rotenon plus Antimycin A, die sequentiell hinzugefügt werden, um die mitochondriale ATP-Synthase zu hemmen, die maximale Atmungskapazität zu bestimmen bzw. die nichtmitochondriale Atemfrequenz zu korrigieren. Diese typische Abfolge von Modulatorzusätzen wird vom Hersteller der Assay-Technologie als MitoStress-Test bezeichnet. Angesichts der Tatsache, dass Oligomycin die vom Entkoppler stimulierte Atmung in einigen Zellmonoschichtenhemmen kann 20, untersuchten wir dies mit krebsabgeleiteten 3D-Sphäroiden, indem wir die entkoppelte stimulierte OCR (OCRmax) vor (einzeln) und nach der (sequentiellen) Oligomycin-Injektion maßen (Abbildung 5A-D). OCRmax war nicht signifikant durch die Zugabe von Oligomycin in Sphäroide aus HEPG2/C3A oder SK-OV-3 begrenzt (Abbildung 5E und Abbildung 5G). OCR max war jedoch in A549- und MCF-7-Sphäroiden nach einer sequentiellen Injektion von BAM15 nach Oligomycin im Vergleich zu OCRmax, das durch eine einzige Injektion von BAM15 erreicht wurde, signifikant gesenkt (Abbildung 5F und Abbildung 5H). Sofern nicht anders bekannt, wird daher empfohlen, separate Vertiefungen zur Behandlung mit Oligomycin und Entkoppler zu verwenden, mit einem letzten Zusatz von Rotenon und Antimycin A bei der Erforschung des mitochondrialen Energiestoffwechsels von 3D-Sphäroiden. Dieser Ansatz ermöglicht immer noch die Berechnung aller mitochondrialen Parameter wie bei einem typischen MitoStress-Test, bei dem Verbindungen sequenziell hinzugefügt werden.

Abbildung 5: Einmalige oder sequentielle Injektion mitochondrialer respiratorischer Verbindungen. Von Krebszellen abgeleitete Sphäroide von MCF-7, HEPG2/C3A, SK-OV-3 und A549 wurden in Vertiefungen einer XFe96-Sphäroid-Mikrotiterplatte in XF RPMI platziert und mit dem Agilent Seahorse XFe96-Analysator auf OCR untersucht. Die OCR wurde 5x gemessen, danach 2 μg/ml Oligomycin (Injektionsport A: grüne Spur) oder 5 μM BAM15 (Injektionsport A: blaue Spur oder Injektionsport B: grüne Spur), um die mitochondriale ATP-Synthase zu hemmen bzw. die maximale Atmungskapazität zu bestimmen. Kinetische OCR-Daten werden in % basal (A-D) ausgedrückt. Die maximale Beatmungskapazität (OCRmax) wurde als Faktor der basalen OCR nach der Gleichung OCRmax = OCRBAM15 / OCRbasal berechnet. OCRmax wurde aus OCR-Durchschnittswerten über Messzyklen 8-10 nach BAM15-Injektion mit (grüne Balken) und ohne (blaue Balken) Oligomycin erhalten. Die Daten sind Durchschnittswerte ± REM von 3-8 einzelnen Bohrlochreplikationen über die Sphäroid-Assay-Mikrotiterplatte. Abkürzungen: OCR = Sauerstoffverbrauch. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Unter Verwendung optimaler Zellaussaatdichten, Verbindungskonzentrationen, Injektionsstrategie und Messzyklusdauer, die in diesen Optimierungsexperimenten bestimmt wurden (Tabelle 3), haben wir ein detailliertes Protokoll für die genaue Untersuchung der basalen mitochondrialen Atmung entwickelt: OCRbasal (Abbildung 6A), ADP Phosphorylierungsatmung: OCRADP (Abbildung 6B), Leckatmung: OCRomy (Abbildung 6C), Kopplungseffizienz (Abbildung 6D ), maximale Atmungskapazität: OCRmax (Abbildung 6E) und freie Atmungskapazität: OCRspare (Abbildung 6F) unter Verwendung von krebsabgeleiteten 3D-Sphäroiden.

Abbildung 6: Untersuchung der OCR mit der XF-Technologie zur Etablierung des mitochondrialen Energiestoffwechsels von Krebs-abgeleiteten Sphäroiden. Krebszell-abgeleitete Sphäroide von MCF-7, HEPG2/C3A, SK-OV-3 und A549 wurden in Vertiefungen einer Sphäroid-Assay-Mikroplatte in XF RPMI platziert und mit dem Agilent Seahorse XFe96-Analysator auf OCR untersucht. OCR wurde 5x gemessen, wonach 2 μg/ml Oligomycin oder 5 μM BAM15 und RA hinzugefügt wurden, um die mitochondriale ATP-Synthase zu hemmen, die maximale Atmungskapazität zu bestimmen und die nichtmitochondriale Atemfrequenz zu berechnen. (A) Die basale mitochondriale Atmung (OCRbasal) wurde als Durchschnitt der OCR aus den 3 Messzyklen vor der Port-A-Injektion berechnet. (B) Die Kopplungseffizienz der oxidativen Phosphorylierung wurde angenähert, indem OCRADP (OCR Basal- OCRLeak) als Prozentsatz von OCRBasal exprimiert wurde. (C) Die ADP-Phosphorylierungsrespiration (OCR ADP) wurde als oligomycinsensitive OCR gemessen, berechnet aus der gemittelten OCR über die Messzyklen 11-13 vor der BAM15-Injektion. (D) DieLeckatmung (OCR-Leck) wurde als OCR-unempfindlich gegen Oligomycin gemessen, berechnet aus der mittleren gemittelten OCR über die Messzyklen 11-13. (E) Die maximale Atmungskapazität (OCRmax) wurde als durchschnittliche maximale OCR gemessen, die nach der Injektion von BAM15 gemessen wurde. (F) Die freie Atemkapazität wurde berechnet, indem OCRSpare (OCR max - OCRbasal) als Prozentsatz der OCRbasal ausgedrückt wurde. OCR nach Rotenon-Antimycin-A-Injektion (OCRr/a) wurde von allen Raten subtrahiert, um die nichtmitochondriale OCR zu korrigieren. Die Daten sind Durchschnittswerte ± SEM von 3-8 einzelnen Bohrlochreplikaten über die XFe96-Sphäroidplatte. Abkürzungen: OCR = Sauerstoffverbrauchsrate; RA = 2 μM Rotenon-2 μM Antimycin A. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
MCF-7-Sphäroide, die aus 4 × 10 3 Zellen / gut über3 Tage gezüchtet wurden, wurden als Modell verwendet, um den optimalen Transfer, die Platzierung und die Analyse in Sphäroid-Assay-Mikroplatten zu bestimmen. Unter Verwendung der für die Sphäroid-Mikroplatte vom Hersteller vorgesehenen Abmessungen wurde die Bohrlochoberfläche für eine optimale Sphäroidplatzierung in drei Zonenbereiche aufgeteilt (Abbildung 7A), wobei Zone 1 als optimale Zone in der Mitte des Bohrlochs hervorgehoben wurde. Durch sorgfältiges Pipettieren unter Verwendung von Pipettenspitzen mit breiter Öffnung wurden Sphäroide in die Sphäroidplatten übertragen und zufällig durch Schwerkraftelution über die Bohrlochoberflächen verteilt (Abbildung 7B). Wo Sphäroide vorsichtig mittels Schwerkraftelution übertragen wurden, konnten die meisten Sphäroide typischerweise in den Zonen 1-2 der Mikrotiterplatte gefunden werden, wobei die empfohlenen Übertragungstechniken des Herstellers verwendet wurden. Wo Sphäroide durch Aspiration aus der Pipettenspitze gedrückt wurden, wurden Sphäroide oft außerhalb dieser Zonen platziert und konnten mit Hilfe der Mikroskopie nicht gesehen werden.
Um die Positionen der Sphäroidplatzierung zu vergleichen, wurden MCF-7-Sphäroide in die Sphäroidassay-Mikroplatten in den ausgewiesenen Zonen 1-3 oder außerhalb der Zone übertragen (Abbildung 7A). Diese 4 Vertiefungen wurden durch eine kinetische Experiment-OCR zu Studienbeginn und nach Zugabe von Oligomycin, BAM15 oder Rotenon-Antimycin A verfolgt (Abbildung 7C). Die OCR wurde aus dem Mittelwert von drei zyklischen Messwerten vor jeder Injektion berechnet (Abbildung 7B). Die OCR wurde kinetisch über 200 min in den 4 ausgewählten Bohrlöchern gemessen (Abbildung 7C) und zu Studienbeginn korrigiert (Abbildung 7D). Wo Sphäroide in Zone 3 oder außerhalb der Zone platziert wurden, waren die Baseline-OCRs signifikant niedriger als die der Sphäroide in den Zonen 1 und 2 (Abbildung 7C). Die Auswirkungen der respiratorischen Verbindungen Oligomycin, BAM15 und Rotenon-Antimycin A unterschieden sich auch dramatisch zwischen den Sphäroiden in den Zonen 1 und 2 im Vergleich zu Zone 3 und Regionen außerhalb der Zone. Ein Anstieg der OCR wurde mit Oligomycin in Sphäroiden beobachtet, die in Zone 3 oder außerhalb der Zone platziert waren (Abbildung 7E). Darüber hinaus zeigten Sphäroide, die in Zone 3 oder außerhalb der Zone platziert wurden, eine übermäßig hohe Reaktion auf BAM15 mit OCR, die höher als der Ausgangswert nach Rotenon-Antimycin-A-Injektion war (Abbildung 7E). Trotz eines fast zweifachen Anstiegs der basalen OCR (Abbildung 7C) mit Sphäroiden in Zone 2 gegenüber Zone 1 waren die Faltenänderungen als Reaktion auf alle respiratorischen Verbindungen sehr ähnlich (Abbildung 7E), was darauf hindeutet, dass Unterschiede in der basalen OCR zwischen Sphäroiden in den Zonen 1 oder 2 wahrscheinlich nicht das Ergebnis der Platzierung innerhalb des Bohrlochs sind.

Abbildung 7: Die Platzierung von Sphäroiden innerhalb der Sphäroid-Assay-Mikroplatte diktiert basale OCR- und mitochondriale Modulatoreffekte unter Verwendung der XF-Technologie. MCF-7-Sphäroide wurden an 4 × 10 3 Zellen / Well ausgesät und über3 Tage kultiviert, bevor sie in die Vertiefungen der Sphäroid-Mikrotiterplatte, die XF RPMI enthielt, gegeben und mit dem Agilent Seahorse XFe96-Analysator auf OCR ± mitochondriale Modulatoren untersucht wurden. (A) Photomikrographien der Positionen der Sphäroidzone in Sphäroid-Assay-Mikrotiterplatten nach der Assay-Dauer; Skalenbalken = 500 μm und OCR, die von entsprechenden Bohrlöchern im Laufe der Zeit erfasst wurden, ausgedrückt als pmol/min-1/Well-1 (B) oder % basal (C). (D) Mitochondriale Modulatoreffekte von MCF-7-Sphäroiden, die in verschiedenen Zonen innerhalb der Sphäroidassay-Mikroplatte platziert sind; Daten, ausgedrückt als Faltungsänderung von basal. (E) Beispiel kinetischer Spuren, die hervorhebt, welche OCR-Datenmessungen (rote Kreise) verwendet werden, um die Reaktion jedes mitochondrialen Modulators auf die in E dargestellten Daten zu berechnen. Die gezeigten Daten stammen aus individuellen Bohrlochantworten. Abkürzung: OCR = Sauerstoffverbrauchsrate. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Die Auswahlkriterien für den Hintergrund sind von hoher Bedeutung; Die Verwendung von äußersten Vertiefungen zur Hintergrundkorrektur ist nicht repräsentativ für alle Mikrotiterplattenvertiefungen, was dazu führen kann, dass falsche Datenannahmen getroffen werden und falsche Datenschlussfolgerungen aufgrund von Kanteneffekten über die Sphäroid-Mikrotiterplatte gezogen werden. Um diese Beobachtung zu bewerten, wurden MCF-7-Sphäroide verwendet, um die Assay-Korrekturverfahren zu vergleichen, um OCR-Werte als Reaktion auf die Zugabe einer Fahrzeugkontrolle, Oligomycin, BAM15 oder Rotenon-Antimycin A abzuleiten (Abbildung 8). Alle respiratorischen Verbindungen ergaben die erwarteten kinetischen OCR-Profile für die ausgewählten Verbindungen, was eine durchschnittliche stetige Basalatmungsrate von 20-30 pmol/min/well ergab (Abbildung 8A). Wenn die Assay-Daten jedoch mit den äußersten Vertiefungen zur Korrektur der Hintergrundtemperatur (A1, A12, H1 und H12) analysiert wurden, waren die Werte für OCR nach der Zugabe von Atemwegsverbindungen besonders niedrig; OCR ergab negative Werte für Rotenon-Antimycin A. Als Reaktion auf diese Beobachtungen wurde eine alternative Analyse mit einer Reihe von leeren Vertiefungen durchgeführt, die zufällig über die Sphäroid-Mikrotiterplatte verteilt waren, als Hintergrundtemperaturkorrektur-Wells (Abbildung 8B). Wenn eine alternative Hintergrundkorrektur angewendet wurde, waren alle relativen zusammengesetzten Effekte auf die OCR in beiden Analysesätzen gleich; Die absoluten OCR-Werte stiegen jedoch um ca. 10 pmol/min/well (Abbildung 8). Diese Daten unterstreichen die Leistungsfähigkeit und Bedeutung der Hintergrundtemperaturkorrektur auf Sphäroid-Assay-Mikrotiterplatten und unterstreichen die Bedeutung der Benutzeroptimierung für die XF-Analyse.

Abbildung 8: Zufällige Auswahl von Vertiefungen zur Hintergrundkorrektur zur Verbesserung der Kontrolle von Temperaturgradienten über die Sphäroid-Assay-Mikrotiterplatte. OCR-Daten, extrapoliert aus Abbildung 2A unter Verwendung empfohlener Vertiefungen für die Hintergrundkorrektur (A) im Vergleich zu zufällig zugewiesenen Vertiefungen für die Hintergrundkorrektur (B). Abkürzung: OCR = Sauerstoffverbrauchsrate. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Im Gegensatz zu Zellmonoschichten stellen Sphäroide eine heterogene Aggregation von Zellen in einem 3D-Raum dar und erfordern daher eine gründliche Betrachtung in Bezug auf die Analyse, insbesondere bei der Normalisierung dieser Daten. Dieses Papier stellt drei Ansätze zur Normalisierung von XF-Daten vor, die von MCF-7-Sphäroiden erfasst wurden (Abbildung 9). Bei Unnormalisierung korreliert OCR positiv (R2 = 0,98) mit der Sphäroidgröße (bestimmt durch die anfängliche Zellaussaatdichte), wenn sie statistisch mit dem Pearson-Korrelationskoeffizienten P = 0,0057 verglichen wird (Abbildung 9A). Diese lineare Beziehung wird verringert, wenn die OCR auf die anfängliche Zellaussaatdichte (R2 = 0,78) normalisiert wird und nicht mehr signifikant mit der Sphäroidgröße korreliert (P = 0,117, Abbildung 9B). Dies ist auch der Fall, wenn auf das Sphäroidvolumen (R2 = 0,77; Pearson-Korrelationskoeffizient P = 0,120, Abbildung 9C) und Kern-dsDNA-Gehalt (R2 = 0,58; Pearson-Korrelationskoeffizient P= 0,233, Abbildung 9D). Diese Daten unterstreichen die Bedeutung der Normalisierung von XF-Daten bei der Untersuchung des mitochondrialen Stoffwechsels von Sphäroiden, insbesondere wenn sie unterschiedlich groß sind.

Abbildung 9: Normalisierung von extrazellulären Flussdaten, die von zellulären Sphäroiden gewonnen wurden. (A) OCR-Rohdaten wurden aus MCF-7 gewonnen, die über 3 Tage kultiviert wurden, und unter Verwendung des Pearson-Modells aufgetragen, um einen Korrelationskoeffizienten zwischen Sphäroid-Seeding-Dichte und OCR zu erhalten; P-Wert auf 0,05 festgelegt. (B) OCR-Rohdaten wurden gegen die anfängliche Sphäroid-Seeding-Dichte normalisiert; C) MCF-7-Sphäroidvolumen, das aus der Mikroskopie-Planimetrie gewonnen wird; und (D) Kern-DS-DNA-Gehalt unter Verwendung des Pearson-Korrelationskoeffizienten. Abkürzungen: OCR = Sauerstoffverbrauchsrate; ds DNA = doppelsträngige DNA. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
| Zelllinie | Aussaatdichte (Brunnen) | Sphäroidwachstum (Tage) | Endgültiges Sphäroidvolumen (μM3) | Basale OCR (pmolO2/min/Well) | Sensitivität für basale OCR erfüllt (JA/NEIN) |
| SKOV | 1000 | 5 | 9,52E+06 | 28 ± 3,5 | Ja |
| SKOV | 2000 | 5 | 2,38E+07 | 26 ± 1,4 | Ja |
| SKOV | 4000 | 5 | 4,92E+07 | 36 ± 3,1 | Ja |
| SKOV | 8000 | 5 | 1,11E+08 | 50 ± 7,9 | Ja |
| HepG2 | 1000 | 5 | 1,11E+07 | 15 ± 0,7 | Nein |
| HepG2 | 2000 | 5 | 2,88E+07 | 23 ± 1,8 | Ja |
| HepG2 | 4000 | 5 | 5,46E+07 | 31 ± 1,7 | Ja |
| HepG2 | 8000 | 5 | 1,21E+08 | 52 ± 2,8 | Ja |
| A-549 | 1000 | 5 | 2.11E+07 | 30 ± 2,5 | Ja |
| A-549 | 2000 | 5 | 3,57E+07 | 41 ± 1,6 | Ja |
| A-549 | 4000 | 5 | 6,93E+07 | 53 ± 7,2 | Ja |
| A-549 | 8000 | 5 | 1,44E+08 | 65 ± 8,4 | Ja |
| MCF-7 | 1000 | 3 | 1,60E+07 | 29 ± 0,8 | Ja |
| MCF-7 | 2000 | 3 | 2,52E+07 | 37 ± 1,7 | Ja |
| MCF-7 | 4000 | 3 | 6.00E+07 | 46 ± 1,7 | Ja |
| MCF-7 | 8000 | 3 | 1,06E+08 | 66 ± 2,9 | Ja |
Tabelle 4: Optimierte Parameter zur Bestimmung basaler OCR-Messungen in einzelnen 3D-Sphäroiden. Abkürzung: OCR = Sauerstoffverbrauchsrate.
Ergänzende Datei 1: Analyse der Sphäroidgröße und des Sphäroidvolumens. Bitte klicken Sie hier, um diese Datei herunterzuladen.
Supplemental File 2: Quantification of double-stränged DNA from spheroids in the spheroid microplate. Bitte klicken Sie hier, um diese Datei herunterzuladen.
Ergänzende Datei 3: Empfehlungen für die Anzahl der Replikate, die erforderlich sind, um zuverlässige XF-Assay-Datensätze zu erhalten. Bitte klicken Sie hier, um diese Datei herunterzuladen.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Wichtigste Ergebnisse und Ergebnisse
Dieses Dokument enthält ein detailliertes Protokoll zur Untersuchung des mitochondrialen Energiestoffwechsels einzelner 3D-Sphäroide unter Verwendung einer Reihe von Krebszelllinien mit dem XFe96 XF Analyzer. Es wird ein Verfahren zur schnellen Kultivierung von A549, HepG2/C3A, MCF7 und SK-OV-3 zellulären Sphäroiden unter Verwendung von zellabweisenden Technologien zur erzwungenen Aggregation entwickelt und beschrieben. Dieses Protokoll befasst sich mit vielen Überlegungen zur Untersuchung des Sphäroidstoffwechsels mit der XF-Technologie, einschließlich (1) der Optimierung von Sphäroidkulturprotokollen und der Handhabung und Übertragung von Sphäroiden in spezifische Sphäroidassay-Mikrotiterplatten des Technologieherstellers aus seinen ursprünglichen Kultivierungsgefäßen; (2) die Konzentration der zu verwendenden Atemwegsverbindungen und die zeitliche Abhängigkeit der Penetration mit Verbindungen; (3) anzuwendende Injektionsstrategien; und (4) Möglichkeiten zur Normalisierung von Daten zwischen experimentellen Gruppen. All diese Überlegungen wurden im aktuellen Papier untersucht und werden im Folgenden näher erläutert. Diese Methoden werden als vereinfachte Ansätze zur Generierung konsistenter metabolischer Sauerstoffflussdaten unter Verwendung einzelner 3D-Sphäroide mit dem XFe96 Flux-Analysator vorgestellt. Dieser experimentelle Ansatz kann als Ausgangspunkt und Rubrik für die Verwendung in anderen Sphäroidmodellen verwendet werden, die in einer grundlegenden Laborumgebung leicht implementiert werden können.
Betrachtungen
Sphäroidwachstum, Größe und Empfindlichkeit der XF-Technologie
Um reproduzierbare Daten mit der XF-Technologie zu ermitteln, ist es unerlässlich, den Assay für das spezifische Modell zu charakterisieren und zu optimieren. Dieser Ansatz ist in einer grundlegenden Monoschicht von Zellen relativ einfach; Dies stellt jedoch zusätzliche Herausforderungen bei der Kultivierung von Zellen als 3D-Sphäroide dar. Während der hier vorgestellten Experimente wurde RPMI-Medium des Herstellers bei der Verwendung ergänzt. Während festgestellt wird, dass einige Zelllinien, nämlich HepG2 / C3A, in DMEM-Wachstumsmedium kultiviert wurden, hatte die Substitution mit RPMI-DMEM-Formulierungen während dieser relativ kurzen Assays (~ 3-5 h) nur begrenzte Auswirkungen auf die XF-Analyse. Die Formulierung der beiden Medien ist sehr ähnlich, und Benutzer könnten Seahorse RPMI-Medien durch Supplementierung, z. B. erhöhte Glukose, weitere Zugabe von Kohlenhydratquellen, an die Matrix ihrer Zellkulturmedien anpassen. Entscheidend für die endgültige Formulierung aller XF-Puffer und -Medien ist das Fehlen von Phenolrot, das wahrscheinlich die fluoreszierenden Sonden in der XF-Sondenkartuschenplatte stört, und Natriumbicarbonat, das aufgrund des Mangels an CO2-Pufferung in Zellkulturinkubatoren zu Alkalinität führt. Andere Medien und Puffer können im eigenen Haus gekauft und/oder hergestellt werden. Zum Beispiel ist Krebs Ringer HEPES-Puffer ein einfacher Puffer, der verwendet werden kann, um die Atmung in vielen verschiedenen Zellen, einschließlich Sphäroidmodellen, zu beurteilen. Benutzer von XF-Assays sollten jedoch beachten, dass eine Änderung des Mediums/Puffers und seine Ergänzung die Gesamtpufferkapazität ändern kann. Dies ist besonders besorgniserregend, wenn Benutzer an der Messung von ECAR interessiert sein könnten, bei der der Pufferfaktor des Mediums bewertet werden muss, um eine ECAR-Umwandlung in die Protonenaustrittsrate (PER) zu ermöglichen.
Da die mit der XF-Technologie gemessene zelluläre OCR proportional zur Zelldichte ist, wenn die Zellzahl im Bohrloch innerhalb der Empfindlichkeit des Systems liegt, war es wichtig, diese Beziehung mit einzelnen 3D-Sphäroiden zu untersuchen. Durch die Untersuchung der OCR einzelner 3D-Sphäroide, die aus 4 verschiedenen Krebszelllinien kultiviert wurden, die mit einer Dichte von 1.000, 2.000, 4.000 oder 8.000 Zellen pro Bohrloch ausgesät wurden, zeigen wir, dass der XFe96-Analysator empfindlich genug ist, um Veränderungen in der Rate der mitochondrialen Atmung zwischen 3D-Sphäroiden zu erfassen, die aus verschiedenen Zellaussaatdichten gezüchtet wurden (Abbildung 3). Wir zeigen, dass sich der optimale Bereich der Zellaussaatdichte und damit des Sphäroidvolumens für die Bildung von 3D-Sphäroiden zur Sondierung von OCR je nach Zelltyp unterscheidet. Dies zeigt die lineare Beziehung zwischen OCR und Seeding-Dichte oder Sphäroidvolumen (Abbildung 3). Für A549- und HepG2/C3A-Zellen lag die optimale Seedingdichte für die OCR-Empfindlichkeit zwischen 1.000 und 8.000 Zellen/Well; es waren 2.000-8.000 Zellen / Well für MCF-7 und 4.000-8.000 Zellen / Well für SK-OV-3 Zellen. Diese Daten zeigen, dass die Optimierung der Sphäroidgröße bei der Beurteilung der OCR mit der XF-Technologie von besonderer Bedeutung ist.
Überlegungen zu minimalen und maximalen Sphäroidvolumina und basaler OCR
Im Allgemeinen wird es immer Mindest- und Höchstschwellen für messbare OCR-Parameter geben, die von der Herstellung für diese Experimente empfohlen werden. Für den XFe96-Analysator sind basale OCR zwischen 20 pmol O 2/min/well und 200 pmol O2/min/well die unteren bzw. oberen Grenzen. Dies ist bei Monolayer-Zellen und Sphäroiden der Fall, und wo das experimentelle Modell innerhalb dieses dynamischen OCR-Bereichs liegt, hängt von der Menge des verfügbaren biologischen Materials ab, z. B. der Anzahl der Zellen als Monoschicht oder der Größe der Sphäroide. Siehe Tabelle 4 für ein Beispiel dafür, wie OCR-Schwellenwerte von den hier verwendeten Sphäroidmodellen erreicht wurden. Es kann ratsam sein, den Sauerstoffgehalt innerhalb des Bohrlochs zu überprüfen, für den diese Daten auch aus diesen Messungen als Füllstandsdaten verfügbar sind. Dies sollte routinemäßig von jedem Experiment zu Qualitätskontrollzwecken betrachtet werden. Wenn es zu Sauerstoffmangel im Brunnen kommt, wird dies in den Daten deutlich. Sollte dies der Fall sein, kann eine Anpassung der Messzyklen innerhalb des Experiments erforderlich sein; Zum Beispiel das Erhöhen des Mischschritts, so dass der Sauerstoffgehalt im Bohrloch vor der nächsten Messperiode innerhalb des Messzyklus wiederhergestellt wird. Obwohl möglich, haben wir festgestellt, dass dies für Einzelsphäroidexperimente mit den beschriebenen Zelllinien sehr unwahrscheinlich ist.
Wahl der mitochondrialen Entkoppler für extrazelluläre Flussassays
Protonenionophore, wie Carbonylcyanid 4-(trifluormethoxy) phenylhydrazon (FCCP)21, Carbonylcyanid m-Chlorphenylhydrazon (CCCP)22 oder BAM15 23, sind potente niedermolekulare Chemikalien, die in der Lage sind, den elektrochemischen Protonengradienten über mitochondriale Membranen zu stören, die Produktion von ATP zu hemmen und letztendlich die mitochondriale Atmung zu entkoppeln 24 . Für diese Zwecke werden weiterhin neue kleine Moleküle entwickelt, insbesondere bei der Behandlung der Stoffwechselerkrankung25,26,27; Beziehen Sie sich auf zwei ausgezeichnete Bewertungen28,29. Umgekehrt wurde die Entkopplung der oxidativen Atmung mit einer unerwünschten Off-Target-Toxizitätin Verbindung gebracht 30. Innerhalb von zellulären In-vitro-Assays depolarisiert das Molekül FCCP jedoch das mitochondriale Membranpotential und übt Off-Target-Effekte wie die Depolarisation der Plasmamembran aus, wodurch der NA+-Ionenfluss31 gestört wird; Interferenz mit der zellulären Proteinverarbeitung32 und sogar Induktion der zellulären Seneszenz33. BAM15 wurde ursprünglich im Jahr 2013 als mitochondrialer Entkoppler mit minimalem Einfluss auf Plasmamembranen 23 eingeführt, mit protonophorer Aktivität im mikromolaren Bereich in ganzen Zellen und nanomolaren Bereich in isolierten Mitochondrien23,34.
Angesichts der Wirksamkeit von FCCP bei der Depolarisation der Plasmamembran ist BAM15 ein zuverlässigeres Protonophor zur Entkopplung der Atmung in intakten ganzen Zellen in extrazellulären Flussassays. Obwohl FCCP und sein Gegenstück, CCCP, seit über 50 Jahren zur Bestimmung der maximalen Atmungskapazität verwendet werden und in XF-Studien weiterhin häufig verwendet werden, unterschätzt die Verwendung dieser kleinen Moleküle oft die mitochondriale und zelluläre Stoffwechselkapazität. Dies hängt zum Teil damit zusammen, warum so viele Publikationen, die die XF-Technologie verwenden, in die Falle tappen, negative Ersatzbeatmungskapazitäten zu melden oder die tatsächlichen mitochondrialen Atmungskapazitäten zu unterschätzen, wenn FCCP verwendet wird. Die zusätzliche Wirksamkeit von FCCP in intakten Zellen und Geweben führt oft zu einer beeinträchtigten mitochondrialen Funktion, und Zellen können Schwierigkeiten haben, angemessen zu arbeiten, um eine maximale Atmungskapazität über mehrere Messzyklen nach ihrer Zugabe aufrechtzuerhalten, selbst bei sehr niedrigen Konzentrationen35. Daher kann die Reaktion der Zellen auf FCCP in vielen Studien gefunden werden, um nach der anfänglichen Messzyklusperiode abzufallen. Während FCCP routinemäßig für die XF-Analyse verwendet wurde, wird BAM15 bevorzugt in Fällen mit ganzen Zellen oder Sphäroidmodellen verwendet, da es eine maximale Atmungskapazität in vollständig depolarisierten Mitochondrien bei Konzentrationen von bis zu 10 μM 3 aufrechterhalten kann. Darüber hinaus induziert BAM15 Wirkungen auf die extrazelluläre Versauerung, die mit der der Nährstoffoxidation durch die Hydratation von CO2 zusammenfällt, um HCO 3- und H + in einem größeren Ausmaß als FCCP3 zu bilden. Im Falle von isolierten Mitochondrien und permeabilisierten Zellen sollte jedoch jeder dieser Entkoppler ebenso gut funktionieren wie BAM15 für die mitochondriale Entkopplung, wenn er in der richtigen Konzentration titriert wird.
Kinetik der Compound-Penetration und des Assay-Zyklus
Die Konzentrationen, die Penetration und die kinetischen Profile chemischer Verbindungen, die zur Durchführung eines typischen MitoStress-Tests mit 3D-Sphäroiden mit dem XF-Analysator verwendet werden, sind komplexer zu behandeln. Angesichts der Tatsache, dass Sphäroide eine 3D-Struktur aufweisen, ist das Eindringen von Molekülen über den Durchmesser des Sphäroids ein unendlich komplexerer Prozess als über Zellmonoschichten. Zum Beispiel wurde die kinetische Penetration und damit die Empfindlichkeit gegenüber dem chemotherapeutischen Sorafenib durch das Sphäroidalter und damit die Größe in einem HepG2-Sphäroid Modell36 bestimmt. Die Fähigkeit niedermolekularer Chemikalien (z. B. Medikamente, Nanopartikel), ein biologisches Ziel zu erreichen, hängt von mehreren zugrunde liegenden Faktoren ab, einschließlich der Komplexität des Systems, das dynamisch durchdrungen und durch37,38 diffundiert werden soll. Dies gilt insbesondere für Medikamente, die auf Tumorgewebe abzielen39. Ähnlich wie beim Tumor-Targeting im Kontext eines 3D-Sphärooids können Größe, Kompaktheit und andere phänotypische Reaktionen wie die Expression von Wirkstofftransporterproteinen die Penetrationszeit und Konzentration einer Verbindung steuern, die erforderlich ist, um eine biologische Reaktion hervorzurufen.
In diesem Protokoll befassten wir uns mit dem Problem der Penetrationszeit und der Konzentration kleiner Moleküle als Reaktion auf den ATP-Synthase-Inhibitor Oligomycin, den Protonophor- und Mitochondrien-Entkoppler BAM15 und die Kombination der Komplex-I- und Komplex-III-Inhibitoren Rotenon und Antimycin A. Durch die Untersuchung der OCR einzelner MCF-7-Sphäroide, die mehreren Titrationen dieser häufigen Atemwegsverbindungen ausgesetzt waren, zeigen wir, dass die optimale Konzentration jeder Verbindung, die erforderlich ist, um eine stationäre Atemfrequenz zu induzieren, in einen ähnlichen Bereich wie bei Monolayer-Zellen liegt (Abbildung 4). Wichtig und anders als bei ihren monolagigen Gegenstücken wird gezeigt, dass die Erhöhung der Anzahl der Messzyklen zwischen den Injektionen der Schlüssel zum Erreichen einer stationären OCR in einzelnen 3D-Sphäroiden ist. Diese Daten unterstreichen die Bedeutung der Penetration von Verbindungen und ihrer jeweiligen kinetischen Profile bei der Untersuchung mitochondrialer Atmungsparameter von 3D-Sphäroiden mit diesen Ansätzen. Unter Verwendung von Sphäroidoptimierungseigenschaften, Konzentrationen von Verbindungen und Messzykluszeiten, die durch die in Abbildung 3, Abbildung 4 und Abbildung 5 dargestellten Daten informiert wurden, wurde ein validierter MitoStress-Test zur Untersuchung spezifischer Parameter des mitochondrialen oxidativen Stoffwechsels in einer Reihe von krebsabgeleiteten 3D-Sphäroiden erstellt (Abbildung 6). Von Bedeutung und wie bei einigen einschichtigen Krebszelllinien40 wurde die maximale Atmungskapazität (Rate der entkoppelten stimulierten Atmung) bestimmter krebsabgeleiteter 3D-Sphäroide durch Oligomycin gehemmt (Abbildung 5). Insbesondere zeigten 3D-Sphäroide, die entweder aus A549- oder MCF-7-Zellen gezüchtet wurden, eine signifikant niedrigere maximale Atmungsrate, wenn sie nach der Oligomycin-Injektion mit BAM15 entkoppelt wurden, verglichen mit der Entkopplung durch BAM15 ohne Oligomycin (Abbildung 5F und Abbildung 5H). Da dieser Effekt in anderen 3D-Sphäroidkulturen vorhanden sein kann, empfehlen wir, dass die maximale Atmungskapazität in 3D-Sphäroiden ohne Oligomycin geschätzt werden sollte, es sei denn, ein zuvor validiertes Protokoll wird verwendet.
Gleichzeitige Erfassung von ECAR-Daten als Maß für den glykolytischen Fluss in zellulären Sphäroiden
Wie typischerweise in der Literatur oder in Informationen des Technologieherstellers zu sehen ist, ist die glykolytische Rate von Sphäroiden, gemessen als ECAR, ein sekundärer Parameter, der neben OCR erfasst werden kann. Die Berechnung von ECAR allein ist in keinem XF-Experiment ein nützlicher oder aussagekräftiger Parameter, da sie nicht um die Pufferkapazität des XF-Assay-Puffers oder die Zugabe von mitochondrialer Versauerung korrigiert wird, die sich aus der Hydratation von CO2 zu HCO3- und H + ergibt. ECAR ist erst dann aufschlussreich, wenn diese Datenkorrekturen angewendet werden, wonach es möglich wird, genauere Rückschlüsse auf den glykolytischen Fluss zu liefern. Um die Pufferkapazität zu korrigieren, um aussagekräftigere PER-Daten zu erzeugen, muss man das Volumen der Mikrokammer für die Sphäroid-Mikroplatte kennen. Der Hersteller war nicht in der Lage, mit der Sphäroid-Mikrotiterplatte ein echtes Volumen dafür bereitzustellen, und daher können PER-Daten nicht leicht bestimmt werden. Obwohl diese Messungen empirisch durchgeführt werden konnten, sprengte dies den Rahmen dieses Manuskripts. Mit den entsprechenden Korrekturen und der Kenntnis des Volumens der Mikrokammer für eine gegebene Sphäroidgröße (z. B. Erhalten eines Maßes für die Sphäroiddichte) in der Vertiefung würden ECAR-Daten jedoch aussagekräftig werden, und Berechnungen der glykolytischen PER könnten durchgeführt werden. Daher könnten XF-Daten dann aussagekräftiger für die Untersuchung des glykolytischen und oxidativen Stoffwechsels in Sphäroiden sein, aber nur, wenn diese Parameter eingehend berücksichtigt würden.
Sphäroidbildung, -handhabung, -transfer und -bewegung
Einige Zelllinien eignen sich besser für die Bildung von Sphäroiden als andere und bilden möglicherweise überhaupt keine Sphäroide, z. B. bilden MCF-7-Eierstockkrebszellen41,42 im Vergleich zu anderen Zelllinien stark kreisförmige Sphäroide (Abbildung 3). Als weiteres Beispiel wurde gezeigt, dass Capan-1-Bauchspeicheldrüsenkrebszellen bessere Sphäroide bilden als Panc-1 oder BxPC343. In ähnlicher Weise ist bekannt, dass Leberkarzinom-Zelllinien variable Fähigkeiten haben, kompakte Sphäroidezu bilden 5,44, mit einer beobachteten Veränderung des Phänotyps wie einem verbesserten Arzneimittelstoffwechsel oder der Produktion von Albumin, wie es bei HepG2 gegenüber HepG2 / C3A 9,45,46 oder HepaRG-Sphäroiden der Fall ist17,47,48 . Daher sollten Anwender die Sphäroid-Kultivierungstechniken entsprechend optimieren und Titrationsexperimente durchführen, um die optimale Aussaatdichte und den Kultivierungszeitverlauf zu bestimmen. Darüber hinaus wurde gezeigt, dass die Formulierung und Zusammensetzung von Assay-Medien die Sphäroidformulierung beeinflusst, einschließlich der Zugabe von Methylcellulose, die häufig zu Medien hinzugefügt wird, um die Matrixviskositätzu erhöhen 43,49,50. Daher sollte die optimale Zellmediumzusammensetzung für alle verwendeten Zelllinien empirisch bestimmt werden.
Die Anzahl der Mediumsaustausche während der Sphäroidkultur wird durch die verwendete Zelllinie bestimmt. In der Regel ist jedoch in den meisten Fällen ein halbvolumiger Mediumaustausch alle 2-3 Tage anwendbar, um Nährstoffe aufzufüllen. Wir verwendeten den Forced-Aggregate-Ansatz, um 3D-Sphäroide unter Verwendung von zellabweisenden Mikrotiterplatten aus kommerziell verfügbaren Quellen für die schnelle Entwicklung und den Einsatz von Sphäroidmodellen in XF-Analysestudien zu generieren. Alternative Plattformen können jedoch besser geeignet sein, um Sphäroide aus anderen Zelltypen zu erzeugen, z. B. Hanging-Drop- oder Matrix-Embedded-Ansätze. In ressourcenbegrenzten Labors sollten Anwender die Agarose-Flüssig-Overlay-Technik für die Bildung von zellabweisenden Mikroplattenoberflächen51,52 in Betracht ziehen, um die wirtschaftlichen Kosten der ersten Entwicklungsschritte der Sphäroidmethode deutlich zu senken. Die Bewegung von Sphäroiden zwischen Kulturgefäßen ist notwendig, um XF-Analysen und andere nachgeschaltete Assays durchzuführen. Die Leichtigkeit der Übertragung wird typischerweise durch die Sphäroidgröße und die Gesamtdichte bestimmt. Wir empfehlen die Verwendung einer P200- oder P1000-Pipettenspitze mit breiter Öffnung, um die Sphäroidintegrität zu erhalten. Pipettenspitzen mit kleinerer Bohrung riskieren eine mechanische Störung des Sphäroides, die im Handel gekauft oder mit Vorsicht durch einfaches Trimmen des Endes der Pipettenspitze hergestellt werden kann, um die Öffnung zu vergrößern. Dieser Ansatz kann jedoch dazu führen, dass der Kunststoff um das Ende der Spitze herum Furing verursacht, was zu mechanischen Störungen während der Handhabung führen kann. Die Verwendung einer Hintergrundbeleuchtung oder eines Leuchtkastens ist auch für die Handhabung und Beobachtung von Sphäroid unter einem Dissektionsmikroskop als wesentlicher Schritt nützlich, um den erfolgreichen Transfer von Sphäroiden in die Sphäroid-Assay-Mikroplatte sicherzustellen. Darüber hinaus ist die Sphäroidposition innerhalb der Vertiefung einer Sphäroid-Assay-Mikroplatte von besonderer Bedeutung und wirkt sich während eines typischen MitoStress-Tests direkt auf die OCR und den Compound-Effekt aus (Abbildung 7), höchstwahrscheinlich aufgrund der Beziehung zwischen der Sphäroidposition und den Fluorophoren der Sensorsonde.
Hintergrundkorrektur und Temperaturregelungsbrunnen
Die Verwendung von Assays auf Mikrotiterplattenbasis ist ein weit verbreiteter Ansatz in mehreren Forschungsbereichen; Ihre Verwendung stellt jedoch mehrere praktische Herausforderungen dar. Wie bei anderen experimentellen Ansätzen, insbesondere solchen, die das 96-Array-Format (oder höher) verwenden, können Mikroplattengeometrie und -positionierung Temperatur- und Gasaustauschgradienten über die Platte im Laufe der Zeit beeinflussen, was oft als "Kanteneffekte" bezeichnet wird53,54. Wir fanden heraus, dass das Gleiche für die Sphäroid-Assay-Mikrotiterplatte gilt. Gemäß den Richtlinien und Protokollen des Herstellers werden die äußersten Eckbohrungen: A1, A12, H1 und H12 immer als Hintergrundkorrektur- und Temperaturkontrollbrunnen für den XFe96-Analysator bezeichnet. Umgekehrt werden beim 24-Well-Array-Format A1 und D6 als Kontrollwellen bezeichnet, zusammen mit zwei anderen Wells, die gleichmäßig über die Mitte der Platte bei B4 und C3 verteilt sind. Bei der Durchführung der XF-Sphäroidanalyse stellten wir eine signifikante Abweichung in den Daten fest, die ursprünglich unter Verwendung der Anweisungen des Herstellers gesammelt wurden. Dies geschah trotz der Einbeziehung der notwendigen Schritte, um das Assay-Gleichgewicht zu Temperatur und CO2 -Gehalt vor Beginn der Datenerfassung sicherzustellen, was oft zu negativen Werten für OCR nach der Injektion bestimmter Atemwegshemmer führte (Abbildung 8).
Wir fanden heraus, dass diese Beobachtungen wahrscheinlich auf Kanteneffekte über die Sphäroid-Assay-Mikrotiterplatte zurückzuführen sind. In Abbildung 8 fanden wir heraus, dass die XF-Daten bei der Neuverteilung der Hintergrundkontrollquellen über die Mikrotiterplatte ungefähr um das 2-fache angepasst wurden. Zwei der wahrscheinlichsten Gründe sind (1) auf Verdunstungseffekte an den Randbohrungen zurückzuführen, die zu einer kleineren Gesamtvolumenkammer für die XFe96-Sonde führen, aus der Proben entnommen werden kann, und (2) aus unzureichenden Temperaturgleichgewichten zwischen den für die Hintergrundkorrektur bestimmten Bohrlöchern und Probenbohrungen, was zu Datensätzen führt, die OCR entweder maskieren oder übermäßig aufblasen. Um solche Ergebnisse zu vermeiden, wird daher empfohlen, insbesondere im Rahmen der Sphäroidanalyse, dass die Benutzer die für die Hintergrundkorrektur vorgesehenen Vertiefungen über die gesamte Sphäroid-Assay-Mikroplatte verteilen und die notwendigen Schritte unternehmen, um ihren Assay vor der Erfassung von XF-Daten vorzuequilibrieren.
Normalisierung von Daten
Neben der Bereitstellung eines detaillierten Protokolls für die Untersuchung des mitochondrialen Energiestoffwechsels einzelner 3D-Sphäroide mit XF-Technologie werden in diesem Artikel auch Möglichkeiten zur Normalisierung mitochondrialer Atemfrequenzdaten vorgestellt, die mit 3D-Sphäroiden erhalten wurden. Unter Verwendung von Atemfrequenzdaten, die mit MCF-7-Sphäroiden erhalten wurden, die bei unterschiedlichen Zellaussaatdichten kultiviert wurden (Abbildung 3), zeigen wir basale mitochondriale Atemfrequenzen von MCF-7-Sphäroiden mit zunehmender Größe und Durchmesser, wenn sie auf die anfängliche Zellaussaatdichte, das Sphäroidvolumen und den dsDNA-Gehalt normalisiert werden (Abbildung 9). Die geeignete Normalisierungsmethode ist für die genaue Interpretation von XF-Datensätzen von größter Bedeutung, insbesondere beim Vergleich von In-vitro-3D-Sphäroidmodellen und verschiedenen Zelltypen. Eine schlechte Normalisierung kann zu fehlerhaften Ergebnissen führen, die einfach nicht zwischen Datensätzen verglichen werden können. Der Proteingehalt wird für die Normalisierung von Sphäroid XF-Daten nicht bevorzugt, da Vorbehandlungen die Proteinsyntheseraten beeinflussen können, ohne dass dies einen signifikanten Einfluss auf die Atemfrequenz hat. Darüber hinaus können signifikante, inkonsistente Proteinmengen bei der Zelllyse an sphäroide Mikroplatten binden, was zu einer Variation des Proteingehalts zwischen den Vertiefungen führt. Dies kann bei XF-Analysen mit Sphäroiden oder nicht adhärenten Zellen, für deren Bindung biomolekulare Klebstoffe erforderlich sind, die Protein enthalten können, noch komplizierter sein.
Im Gegensatz zum intrazellulären Proteingehalt ist der Kern-DNA-Gehalt unabhängig vom Zelltyp und proportional zur Zellzahl (Abbildung 9D) - ein genauerer und weniger zeitaufwendiger Ansatz als die Disaggregation von Sphäroiden zur Quantifizierung der Zellzahl. Umgekehrt fanden Yepéz et al.55, die XF-Analysen in Monoschichten von Fibroblastenzellen durchführten, heraus, dass die Normalisierung von XF-Daten auf die Zellzahl zu einer größeren Dispersion der Daten führte als vor der Normalisierung. Der Kern-DNA-Gehalt ist unabhängig vom differenzierten Zustand oder Phänotyp und daher für die Normalisierung von Sphäroiddaten in XF-Assays genauer als der Proteingehalt. Der DNA-Gehalt war auch eine bewährte Strategie für die Analyse anderer mit dem Stoffwechsel verbundener Datensätze56. Es ist jedoch wichtig zu beachten, dass der Kern-DNA-Gehalt aus allen im Sphäroid vorhandenen Zellen quantifiziert wird; Daher wird die Normalisierung auf den DNA-Gehalt nicht für XF-Datensätze empfohlen, bei denen Sphäroide Behandlungen unterzogen werden, die zu einem signifikanten Verlust der Zelllebensfähigkeit führen können. Für solche Datensätze wird, wenn möglich, die Normalisierung auf die Zelllebensfähigkeit bevorzugt, oder die Daten können zu einer Basalatmung korrigiert werden.
Nutzung der freien Atmungskapazität als Beispiel für die Bedeutung der Datennormalisierung
Die freie Atmungskapazität ist ein Maß für die Rate der maximalen mitochondrialen Atmungskapazität abzüglich der basalen mitochondrialen Atemfrequenz (Abbildung 6). Das Problem bei der Meldung von Daten dieser Art als Rate, d. H. PmolO2 / min / well in bestimmten Experimenten, besteht jedoch darin, dass die Daten keine Normalisierung aufweisen. Selbst wenn man Sphäroiddaten auf Zelldichte/DNA-Gehalt normalisiert, schließt dies oft den Schlüsselparameter aus, der für die mitochondriale Dichte innerhalb der Zellen normalisiert werden muss. Angesichts der Tatsache, dass eine Änderung der mitochondrialen Dichte zu einer proportionalen Änderung der basalen und maximalen Atmung führt, wird auch die freie Kapazität zunehmen. Wenn beispielsweise die sphäroide OCRbasal 200 und die maximaleOCR 400 beträgt, wird die freie Kapazität mit 200 angegeben. Wenn OCRbasal 100 und OCRmax ist, dann ist die freie Kapazität auch 100; Als Prozentsatz sind sie jedoch beide 50% von maximal (oder 100% von basal). Daher wird die freie Kapazität zwischen diesen beiden Beispielen nicht verändert, trotz Unterschieden in den Raten von 200 und 100, wenn sie als pmols O2 / min / well berechnet werden. Intern normalisierte Werte sind zuverlässiger und aufschlussreicher, um XF-Daten über Studien und Projekte hinweg vergleichbarer zu machen. Um dies für die freie Atemkapazität zu tun, haben wir uns entschieden, dies als Prozentsatz der maximalen Atmung anstelle einer absoluten Rate darzustellen. Dies könnte auch als Prozentsatz der Basalatmung dargestellt werden. Dies wäre der Fall, wenn mit Zellen oder Sphäroiden gearbeitet wird. Da jedoch die Lage des Sphäroids in der Mikrotiterplatte die absolute OCR verändern kann, nicht aber die relativen Veränderungen mit Inhibitoren oder Entkopplern, ist es wichtiger, intern normalisierte Reaktionen in Sphäroiden als Faltenänderung oder Prozentsätze zu betrachten.
Die hier generierten Sphäroidmodelle stellen eine Reihe von Zelltypen und Architekturen dar, die in klassischen 2D-Modellen nicht erfasst werden können. Dazu gehören heterogene, räumliche Anordnung von Zellen in drei Dimensionen, verbesserte Zell-Zell-Kontakte (z. B. Bildung von Gap Junctions und extrazellulären Matrizen) und biochemische Gradienten über den Sphäroiddurchmesser (z. B. pH-Gradienten, Sauerstoffdiffusionszugang zu Nährstoffen). Die Verwendung des extrazellulären Flusses zur Untersuchung der In-vitro-Sphäroidbiologie könnte es ermöglichen, optimale Ziele für medikamentöse Therapien durch metabolische Störungsbeobachtungen zu identifizieren. Diese könnten von In-vitro-Sphäroiden auf In-vivo-Tumoren extrapoliert werden und Wege identifizieren, die auf den Sphäroid-Tumor-Stoffwechsel abzielen können, z. B. die Kohlenhydratverwertung während des Sphäroidwachstums. Therapeutische Modalitäten können bei der Bekämpfung von Sphäroiden in frühen Wachstumsphasen wirksam sein, erweisen sich aber in den späteren Phasen des Sphäroidwachstums als weniger wirksam, wenn die Komplexität des metabolischen Netzwerks reift. Zusammenfassend lässt sich sagen, dass die Entwicklung von 3D-Zellkulturmodellen und ausgefeilten Analysetechnologien in der biologischen Forschung weiterhin ein dynamisches und sich schnell veränderndes Feld mit unübertroffenem Potenzial sein wird. Die extrazelluläre Flussanalyse von In-vitro-Zellkultur-Sphäroiden könnte als modernste Forschungsmethode eingesetzt werden, um Forschungsergebnisse voranzutreiben, die extrapoliert werden könnten, um die humanrelevante Biologie besser zu verstehen, die Verwendung von Tiermodellen in der Forschung zu reduzieren und die patientenzentrierte Forschung zu verbessern.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Die Autoren haben keine Interessenkonflikte zu erklären.
Acknowledgments
N.J.C wurde durch einen BBSRC MIBTP CASE Award mit Sygnature Discovery Ltd unterstützt (BB/M01116X/1, 1940003)
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
Krebsforschung Ausgabe 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
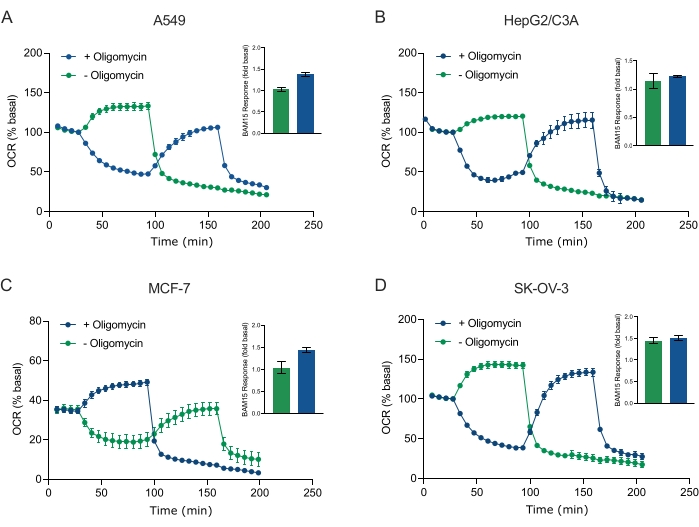
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
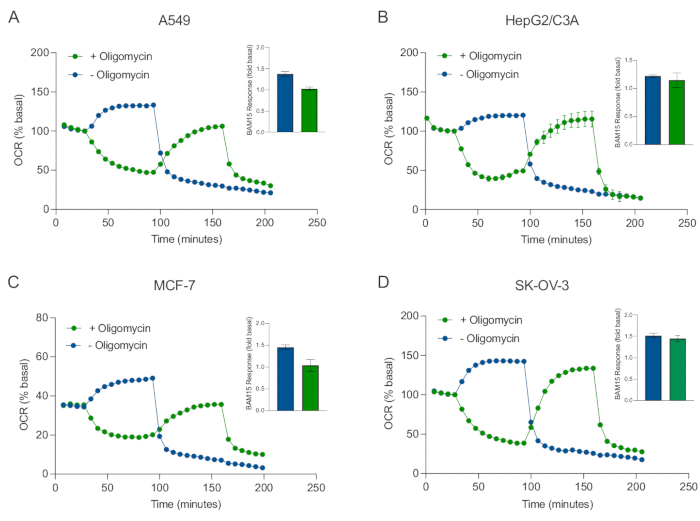
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
