

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Esses protocolos ajudarão os usuários a sondar o metabolismo de energia mitocondrial em esferoides derivados de células cancerígenas 3D usando a análise de fluxo extracelular do Seahorse.
Abstract
Agregados celulares tridimensionais (3D), denominados esferoides, tornaram-se a vanguarda da cultura celular in vitro nos últimos anos. Em contraste com as células cultivadas como monocamadas bidimensionais e unicelulares (cultura 2D), a cultura celular esfóide promove, regula e suporta arquitetura fisiológica celular e características que existem in vivo, incluindo a expressão de proteínas de matriz extracelular, sinalização celular, expressão genética, produção de proteínas, diferenciação e proliferação. A importância da cultura 3D tem sido reconhecida em muitas áreas de pesquisa, incluindo oncologia, diabetes, biologia de células-tronco e engenharia de tecidos. Na última década, métodos aprimorados foram desenvolvidos para produzir esferoides e avaliar sua função metabólica e destino.
Analisadores de fluxo extracelular (XF) têm sido usados para explorar a função mitocondrial em microtissues 3D, como esferoides usando uma placa de captura de ilhotas XF24 ou uma microplaca esferoide XFe96. No entanto, protocolos distintos e a otimização da sondagem do metabolismo de energia mitocondrial em esferoides usando a tecnologia XF não foram descritos em detalhes. Este artigo fornece protocolos detalhados para sondar o metabolismo de energia mitocondrial em esferoides 3D únicos usando microplapes esferoides com o analisador XFe96 XF. Usando diferentes linhas de células cancerígenas, a tecnologia XF é demonstrada ser capaz de distinguir entre a respiração celular em esferoides 3D não apenas de tamanhos diferentes, mas também diferentes volumes, números celulares, conteúdo de DNA e tipo.
As concentrações compostas de efeito mitocondrial ideal de oligomicina, BAM15, rotenona e antimicina A são usadas para sondar parâmetros específicos do metabolismo de energia mitocondrial em esferoides 3D. Este artigo também discute métodos para normalizar dados obtidos a partir de esferoides e aborda muitas considerações que devem ser consideradas ao explorar o metabolismo esferoide usando a tecnologia XF. Este protocolo ajudará a impulsionar a pesquisa em modelos esferoides in vitro avançados.
Introduction
Os avanços em modelos in vitro em pesquisa biológica progrediram rapidamente nos últimos 20 anos. Tais modelos agora incluem modalidades organ-on-a-chip, organoides e esferoides microtissue 3D, todos os quais se tornaram um foco comum para melhorar a tradução entre estudos in vitro e in vivo. O uso de modelos in vitro avançados, particularmente esferoides, abrange diversos campos de pesquisa, incluindo engenharia de tecidos, pesquisa de células-tronco, câncer e biologia de doenças 1,2,3,4,5,6,7, e testes de segurança, incluindo toxicologia genética 8,9,10, toxicologia de nanomateriais11, 12,13,14, e testes de segurança e eficácia de medicamentos 8,15,16,17,18,19.
A morfologia celular normal é fundamental para o fenótipo biológico e a atividade. A colheita de células em esferoides microtissue 3D permite que as células adotem uma morfologia, função fenotípica e arquitetura, mais semelhantes às observadas in vivo , mas difíceis de capturar com técnicas clássicas de cultura celular monocamada. Tanto in vivo quanto in vitro, a função celular é diretamente impactada pelo microambiente celular, que não se limita à comunicação e programação celular (por exemplo, formações de junção celular-célula, oportunidades de formação de nichos celulares); exposição celular a hormônios e fatores de crescimento nos ambientes imediatos (por exemplo, exposição à citocina celular como parte de uma resposta inflamatória); composição de matrizes físicas e químicas (por exemplo, se as células são cultivadas em plástico de cultura tecidual rígida ou um ambiente de tecido elástico); e, mais importante, como o metabolismo celular é impactado pela nutrição e acesso ao oxigênio, bem como pelo processamento de resíduos metabólicos, como o ácido láctico.
A análise do fluxo metabólico é uma maneira poderosa de examinar o metabolismo celular dentro de sistemas in vitro definidos. Especificamente, a tecnologia XF permite a análise de mudanças ao vivo e em tempo real nos bioenergésicos celulares de células e tecidos intactos. Dado que muitos eventos metabólicos intracelulares ocorrem dentro da ordem de segundos a minutos, abordagens funcionais em tempo real são primordiais para entender mudanças em tempo real no fluxo metabólico celular em células intactas e tecidos in vitro.
Este artigo fornece protocolos para o cultivo das linhas celulares derivadas do câncer A549 (adenocarcinoma pulmonar), HepG2/C3A (carcinoma hepatocelular), MCF-7 (adenocarcinoma mamário) e SK-OV-3 (adenocarcinoma ovariano) como modelos esferoides in vitro 3D usando abordagens de agregação forçada (Figura 1). Ele também (i) descreve em detalhes como sondar o metabolismo de energia mitocondrial de esferoides 3D únicos usando o analisador Agilent XFe96 XF, (ii) destaca maneiras de otimizar os ensaios XF usando spheroids 3D únicos, e (iii) discute considerações e limitações importantes de sondagem do metabolismo esferoide 3D usando essa abordagem. Mais importante, este artigo descreve como são coletados conjuntos de dados que permitem o cálculo da taxa de consumo de oxigênio (OCR) para determinar a fosforilação oxidativa e, portanto, a função mitocondrial em esferoides celulares. Embora não seja analisada para este protocolo, a taxa de acidificação extracelular (ECAR) é outro parâmetro que é medido ao lado de dados OCR em experimentos XF. No entanto, o ECAR é muitas vezes mal ou incorretamente interpretado a partir de conjuntos de dados XF. Nós fornecemos um comentário sobre as limitações de cálculo do ECAR seguindo abordagens básicas do fabricante de tecnologia.
Subscription Required. Please recommend JoVE to your librarian.
Protocol

Figura 1: Fluxo de trabalho gráfico para a geração de esferoides celulares, análise de fluxo extracelular e ensaios a jusante. Quatro linhas de células cancerígenas foram seletivamente cultivadas como monocamadas (A), separadas dos frascos da cultura tecidual, e semeadas em microplacas de 96 poços para formar esferoides (B). Carcinoma pulmonar A549, carcinoma hepg2/C3A, As células de adenocarcinoma ovariano SK-OV-3 e as células de carcinoma mamário MCF-7 foram semeadas em 1 × 103-8 × 103 células/bem e cresceram até 7 dias para formar esferoides únicos e otimizar a densidade e o tempo de cultivo de sementes e cultivo e esferoide por meio de observação contínua e medidas planimétricas. Uma vez formados, os sferóides únicos foram lavados em um meio XF livre de soro e cuidadosamente semeados em microplacas de ensaio esferoide, pré-revestidas com poli-D-lysine (C). Os spheroids foram submetidos à análise de fluxo extracelular usando o analisador XFe96 usando vários protocolos para abordar: (1) tamanho esferoide ideal para resposta à respiração mitocondrial basal; (2) titulação otimizada de inibidores respiratórios mitocondriais; (3) otimização da colocação esferoide dentro de poços de microplacão. (D) Análises pós-XF, microscopia de contraste de fase e quantificação de DNA esferoide foram utilizadas para normalização de dados e outros ensaios in vitro a jusante. Clique aqui para ver uma versão maior desta figura.
1. Cultivo de linhas de células cancerosas como esferoides in vitro 3D
| Linha celular | Descrição | Meio de cultura | Fonte |
| A549 | Linha celular de carcinoma pulmonar | RPMI 1640 | Coleção Europeia de Culturas Celulares Autenticadas (ECACC) |
| Piruvato de sódio (1 mM) | |||
| Penicilina- Estreptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| HepG2/C3A | Linha celular de carcinoma hepático, um derivado clonal da linha de células HepG2 pai | DMEM | Coleção Americana de Cultura de Tecidos (ATCC) |
| Penicilina- Estreptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| MCF7 | Linha celular de adenocarcinoma mamário | RPMI 1640 | Coleção Europeia de Culturas Celulares Autenticadas (ECACC) |
| Piruvato de sódio (1 mM) | |||
| Penicilina- Estreptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| SK-OV-3 | Linha celular adenocarcinoma ovariana | RPMI 1640 | Coleção Europeia de Culturas Celulares Autenticadas (ECACC) |
| Piruvato de sódio (1 mM) | |||
| Penicilina- Estreptomicina - (100 U/mL – 100 mg/mL) | |||
| 10 % (v/v) FBS | |||
| Componente | Meio de ensaio RPMI (volume final de 50 mL) | ||
| Meio base | Agilent Seahorse XF RPMI, pH 7.4 | ||
| Glicose (1 M estoque estéril) | 11 mM (solução de estoque de 0,55 mL) | ||
| L-glutamina (estoque estéril de 200 mM) | 2 mM (0,5 mL de solução de estoque) | ||
| Piruvato de sódio (estoque estéril de 100 mM) | 1 mM (0,5 mL de solução de estoque) |
Tabela 1: Mídia de linha celular cancerosa e composições de mídia XF.
- Cultize todas as linhas celulares usando a técnica padrão de cultura de tecido asséptico e confirme que elas estão livres de mycoplasma usando um kit de ensaio adequado.
- Cultura as linhas celulares em frascos de cultura tecidual T75 ou equivalente, utilizando o meio recomendado (Tabela 1). Cultume as linhas celulares para 65-80% de confluência e passagem-as regularmente até um máximo de 25 passagens.
- Enxágüe os frascos de cultura celular duas vezes na solução salina modificada de fosfato (DBPS) modificada de Dulbecco.
- Retire as células dos frascos com 3 mL do reagente de dissociação celular (ver a Tabela de Materiais) por 5 min a 37 °C e confirme o descolamento por microscopia.
- Aspire suavemente a suspensão da célula isolada para garantir uma suspensão unicelular e desativar o reagente de dissociação celular com 7 mL de meio completo de cultura tecidual.
- Recolher as células por centrifugação a 300 × g por 5 min, descartar o supernascedor e resuspensar as células em meio completo.
- Conte as células usando um hemótmetro ou um contador celular automatizado e titulação à densidade celular desejada necessária para semeadura.
NOTA: Para semear uma placa inteira de 96 poços a 100 μL/well a 4 × 103 células/bem, as células devem ser tituladas a 4 × 104 células/mL em um volume recomendado de 12 mL. - Decante a suspensão da célula em um reservatório estéril e dispense 100 μL da suspensão celular em cada poço de uma microplaca repelente celular usando um pipettor multicanal.
NOTA: Apenas os 60 poços internos de uma microplaca devem ser semeados e o restante preenchido com DPBS. Isso formará uma barreira de evaporação, garantirá a homogeneidade esferoide através da placa e minimizará os efeitos da borda da placa. - Centrífugas microplatas esferoides a 300 × g por 15 minutos para forçar as células a agregados soltos.
- Incubar as placas a 37 °C, 5% de CO2 por um mínimo de 3 dias para garantir a formação de esferoides.
- Realizar microscopia de contraste de fase utilizando práticas laboratoriais padronizadas para monitorar o crescimento de esferoides. Reponha o meio de cultura celular a cada 3 dias ou duas vezes por semana, realizando uma troca média de meio volume.
2. Sondagem do metabolismo de energia mitocondrial de esferoides únicos usando tecnologia de fluxo extracelular (XF)
- Preparação de ensaios (um dia antes)
- Verifique a viabilidade esferoide usando um microscópio de luz invertida com contraste de fase na ampliação de 4x para garantir estrutura eferóide intacta, morfologia e uniformidade geral entre as amostras.
- Hidrate o cartucho do sensor.
- Alíquota ~20 mL do calibrante em um tubo cônico.
- Coloque o tubo cônico contendo o calibrante em uma incubadora não CO2 37 °C durante a noite.
- Remova o conteúdo do kit de ensaio.
- Remova o cartucho do sensor da placa do utilitário e coloque-o de cabeça para baixo na bancada ao lado da placa de utilidade.
- Pipeta 200 μL de ddH2O estéril em cada poço da placa de utilidade do cartucho do sensor usando uma pipeta P300 multicanal.
- Coloque o cartucho do sensor em cima da placa de utilidade.
- Verifique se o nível de água em cada poço é alto o suficiente para submergir as sondas do sensor.
- Transfira o cartucho do sensor montado para uma incubadora não-CO2 de 37 °C e deixe-o durante a noite.
NOTA: Esta etapa pode ser realizada 12-72 h antes do início do ensaio.
- Talo spheroid ensaio microplato
- Utilizando técnicas assépticas, adicione 30 μL/poço de solução estéril de Poli-D-Lysine (0,1 mg/mL) à microplacão e incuba-a por 30 minutos à temperatura ambiente.
- Aspire a solução de cada poço da microplacão esferoide, inverta a placa e bata-a firmemente no papel de tecido para remover qualquer solução residual.
- Lave a placa duas vezes com 200 μL/poço de ddH2O estéril.
- Após a lavagem final, inverta a microplacão e bata-a firmemente no papel de tecido para remover qualquer água residual.
- Deixe a placa secar ao ar por 30 minutos antes de usá-la ou armazená-la a 4 °C para uso futuro.
NOTA: A microplacão de ensaio esferoide deve ser revestida com um adesivo molecular para garantir que os esferoides sejam fixados na parte inferior da microplacão. Sem um adesivo molecular, os esferoides podem se desalojar e interferir com os resultados do ensaio. Outros adesivos moleculares também podem ser usados como uma alternativa à Poli-D-Lysine para pré-revestimento de placas. As placas pré-revestidas podem ser armazenadas a 4 °C, mas devem ser deixadas para equilibrar a temperatura ambiente antes do início do ensaio.
- Prepare o meio de ensaio XF
- Prepare o meio XF RPMI, conforme detalhado na Tabela 1, e filtro estéril com um filtro de seringa de 0,22 μm
- Preparação de ensaio (1h antes do ensaio)
- Pré-aquecimento do ensaio XF RPMI suplementado médio a 37 °C.
- Pré-aquecimento o ensaio esferoide revestido microplaca em uma incubadora não CO2 37 °C ou banho seco.
- Prepare o cartucho do sensor.
- Retire o tubo cônico contendo o calibrante e o cartucho do sensor da incubadora de ar.
- Remova o cartucho do sensor da placa do utilitário e coloque-o de cabeça para baixo na superfície de trabalho.
- Usando uma pipeta multicanal P300, aspire a água da placa de utilidade e descarte-a.
- Despeje a solução calibrante em um reservatório de reagente estéril e adicione 200 μL/poço do calibrante pré-caçado à placa de utilidade usando uma pipeta multicanal P300.
- Pegue o cartucho do sensor e coloque-o de volta em cima da placa de utilidade, garantindo que os sensores estejam bem submersos no calibrante.
- Transfira o cartucho de sensor montado de volta para a incubadora não-CO2 37 °C até estar pronto para carregar as soluções de injeção da porta.
- Lave os esferoides com o meio de ensaio.
- Remova a placa de cultura esferoide da incubadora de 37 °C, 5% de CO2 e observe os esferoides sob o microscópio para garantir sua integridade antes das etapas de transferência de esferoide.
- Carregue todos os poços da placa esferoide com 180 μL/poço de meio de ensaio pré-armado, incluindo quaisquer poços de correção de fundo.
- Encha parcialmente uma placa de Petri de 7 cm com 3 mL do meio de ensaio.
- Usando uma pipeta multicanal carregada com pontas de pipeta de orifício largo, transfira os esferoides da placa de cultura de 96 poços em pratos petri de 7 cm, colocando o pipettor em um volume de aspiração de 10-50 μL.
- Esferoides de sementes na microplacão de ensaio esferoide pré-revestido.
- Usando um microscópio de dissecção e um aparelho de caixa de luz, transfira os esferoides da placa de Petri para a microplaca de ensaio esferoide conforme detalhado abaixo.
- Defina o volume de um pipettor de um único canal equipado com uma ponta de pipeta de orifício largo para 20 μL e aspire cuidadosamente um único esferoide. Coloque a ponta diretamente no centro de cada poço da microplacão de ensaio esferoide e deixe que a gravidade esfolóide no centro de cada poço, ou seja, não expulse nenhum meio da ponta pipeta e permita que a ação capilar retire o esferoide da ponta pipeta. Para confirmar a eluição, o conteúdo do pipettor pode ser pipetado de volta para a placa de Petri de 7 cm sob o microscópio.
NOTA: A eluição de gravidade de um único esferoide normalmente leva 15-30 s, dependendo do tamanho/densidade esferoide. Durante este tempo, o pipettor não deve ser removido. Todos os poços de correção de fundo devem estar livres de esferoides e conter apenas o meio de ensaio. Sob o microscópio, confirme a posição de cada esferoide. Cada esferoide deve ser posicionado idealmente dentro do centro de cada poço. - Uma vez que todos os esferoides tenham sido transferidos para a microplaca de ensaio esferoide, transfira a placa para uma incubadora não-CO2 a 37 °C por um mínimo de 1h antes do ensaio.
- Defina o volume de um pipettor de um único canal equipado com uma ponta de pipeta de orifício largo para 20 μL e aspire cuidadosamente um único esferoide. Coloque a ponta diretamente no centro de cada poço da microplacão de ensaio esferoide e deixe que a gravidade esfolóide no centro de cada poço, ou seja, não expulse nenhum meio da ponta pipeta e permita que a ação capilar retire o esferoide da ponta pipeta. Para confirmar a eluição, o conteúdo do pipettor pode ser pipetado de volta para a placa de Petri de 7 cm sob o microscópio.
- Usando um microscópio de dissecção e um aparelho de caixa de luz, transfira os esferoides da placa de Petri para a microplaca de ensaio esferoide conforme detalhado abaixo.
3. Preparação e carregamento de compostos no cartucho do sensor para ensaios XF
| Estratégia de Injeção | Composto (Porta) | Volume de partida de microwell XFe96 (μL) | Concentração final de poços desejada | Volume da porta (μL) | Injeção final de microwell volume XFe96 (μL) | Concentração de estoque de trabalho |
| 1 | Oligomicina (A) | 180 | 3 ug/mL | 20 | 200 | 30 μg/mL |
| Rotenone (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimicina A (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| 2 | BAM15 (A) | 180 | 5 μM | 20 | 200 | 50 μM |
| Rotenone (B) | 200 | 2 μM | 20 | 220 | 22 μM | |
| Antimicina A (B) | 200 | 2 μM | 20 | 220 | 22 μM |
Tabela 2: Concentrações de compostos mitocondriais para sondar o metabolismo de energia mitocondrial de esferoides 3D únicos usando o Analisador XFe96.
- Prepare as concentrações de estoque de trabalho de cada composto, conforme observado na Tabela 2 usando um meio de ensaio XF RPMI totalmente suplementado e pré-equipado.
- Oriente a placa do cartucho (acoplado à placa de utilidade) em termos de coluna, 1-12 da esquerda para a direita.
- Se usar um guia de carga, coloque-o sobre a placa do cartucho de acordo com o procedimento de carregamento de poço, por exemplo, se a porta A for carregada primeiro, certifique-se de que A esteja visível no canto superior esquerdo da guia.
- Transfira a solução de funcionamento de cada composto para um reservatório adequado e, utilizando uma pipeta multicanal P100 calibrada, dispense 20 μL em todas as portas correspondentes. Repita cada composto nas portas restantes.
NOTA: Se não forem utilizadas as portas na placa do cartucho do sensor, estas podem ser deixadas vazias ou preenchidas com meio de ensaio. Se houver apenas uma seleção de uma letra de porta específica, certifique-se de que as outras portas correspondentes a essa letra estejam carregadas com meio de ensaio; caso contrário, o ar será injetado no poço, comprometendo os resultados nesses poços. - Após o carregamento da porta, remova as guias de carregamento da placa (se utilizada) e prepare o analisador para carregar o cartucho do sensor.
NOTA: Se o ensaio não estiver sendo executado imediatamente após o carregamento das portas, coloque a tampa de volta no cartucho do sensor e coloque a placa de volta na incubadora de ar de 37 °C até estar pronta para carregar na máquina.
4. Projeto de ensaio, estratégias de injeção e aquisição de dados
- Executando o ensaio
- Ligue o analisador e conecte-se ao controlador (computador).
NOTA: Isso pode ser verificado pelo status de conexão de instrumento no painel de widget do software Wave Controller. - Navegue até a página de modelos no software WAVE, encontre o arquivo de modelo de ensaio para o experimento e clique duas vezes para abri-lo.
NOTA: Se o modelo de ensaio não aparecer na exibição Modelos , importe o arquivo de modelo para a pasta de modelo de uma unidade de rede compartilhada ou unidade flash USB. - Para iniciar o ensaio, clique na guia Executar ensaios .
NOTA: Se as definições de grupo tiverem sido corretamente alocadas dentro do mapa da placa, o ensaio estará pronto para ser executado conforme indicado pelo carrapato verde no lado direito da página. Nesta fase, qualquer informação adicional pode ser inserção na página de resumo do ensaio ou na página deixada em branco; prossiga para o próximo passo. Devido à penetração retardada de moduladores mitocondriais em esferoides microtissue 3D (Figura 2), utilize as informações do protocolo de medição descritas na Tabela 3.
- Ligue o analisador e conecte-se ao controlador (computador).
| Período de medição | Número de injeção e porta | Detalhes de medição | Duração do período (h:min:s) |
| Calibração | Não é aplicável | Os analisadores XF sempre realizam esta calibração para garantir que as medidas sejam precisas | 00:20:00 (esta é uma média e pode variar entre máquinas) |
| Equillibração | Não é aplicável | O equilíbrio ocorre após a calibração e é recomendado. | 00:10:00 |
| Basal | Não é aplicável | Ciclos = 5 | 00:30:00 |
| Mix = 3:00 | |||
| Espera = 0:00 | |||
| Medida = 3:00 | |||
| Oligomicina / BAM15 | Injeção 1 (Porta A) | Ciclos = 10 | 01:00:00 |
| Mix = 3:00 | |||
| Espera = 0:00 | |||
| Medida = 3:00 | |||
| Rotenona + antimicina A | Injeção 2 (Porta B) | Ciclos = 10 | 01:00:00 |
| Mix = 3:00 | |||
| Espera = 0:00 | |||
| Medida = 3:00 | |||
| Tempo total: | 03:00:00 | ||
Tabela 3: Configuração do protocolo para sondagem do metabolismo de energia mitocondrial de esferoides 3D únicos usando o Analisador XFe96.
- Clique em iniciar a execução para trazer a caixa de diálogo de localização de salvamento .
- Digite o local de salvamento para o arquivo de resultado e coloque o cartucho do sensor montado na bandeja térmica que aparece da porta na lateral do analisador. Aguarde que a bandeja térmica abra automaticamente e a tela exiba a mensagem de placa de utilidade do calibrador de carga. Antes de seguir as instruções na tela, certifique-se de que i) ajuste adequado do cartucho do sensor na placa utilitário, ii) a tampa é removida do cartucho do sensor, e iii) orientação correta do cartucho do sensor na placa do utilitário.
- Siga os comandos na tela para iniciar a calibração do cartucho do sensor.
NOTA: O tempo necessário para completar a calibração é de aproximadamente 10-20 min (para ensaios a 37 °C). - Após a calibração do cartucho do sensor, carregue a microplaca esferoide no analisador seguindo as instruções na tela no Controlador de Ondas para iniciar a etapa de equilíbrio de 12 minutos.
NOTA: Caixas verdes com carrapatos brancos indicam uma calibração "boa" para esse poço. Se algum poço não fornecer uma calibração "boa", eles serão indicados com uma caixa vermelha e cruz branca. Esses poços devem ser observados e excluídos de qualquer análise após a conclusão do ensaio usando a guia de ensaio de modificação . - Aguarde que o analisador comece automaticamente a adquirir medições de linha de base após a máquina ter concluído a etapa de equilíbrio (conforme descrito no protocolo do instrumento).
- Para completar o experimento, siga os comandos na tela no controlador WAVE.
NOTA: Uma vez que a microplacã esferoide tenha sido removida do analisador, descarte o cartucho do sensor e reserve a placa esferoide para análise posterior, se necessário (por exemplo, quantificação de DNA de dupla fita). Se a microplacão não for necessária para análise suplementar, ela pode ser descartada junto com o cartucho do sensor. - Aguarde que a caixa de diálogo de ensaio apareça e visualize os resultados ou retorne à exibição de modelos .
5. Estratégias de normalização e análise de dados - normalização pós-ensaio e ensaios a jusante (etapas opcionais)
- Normalização de dados
- Para normalizar os dados esferoides, consulte a série de protocolos pertinentes às estratégias de normalização de dados para calcular o tamanho e o volume e quantificar o DSDNA em ensaios esferoides. Estes foram incluídos como arquivos suplementares; ver arquivo suplementar 1 e arquivo suplementar 2.
- Análise de dados
- Para exportar dados para um dos geradores de análise automatizados, siga os comandos de exportação de dados no controlador WAVE e selecione o gerador de exportação que corresponde ao tipo de ensaio. Alternativamente, exporte o arquivo de dados e carregue-o para análises do Seahorse.
NOTA: A desvantagem dos geradores de relatórios e da análise de Seahorse é que a análise de dados está limitada à forma como o ensaio XF é projetado e não permite que médias sejam tomadas em todos os ciclos de medição. A exportação manual de conjuntos de dados do software de instrumento permite a preferência do usuário a esse respeito. Dado que a estratégia de injeção para avaliar a respiração mitocondrial de esferoides 3D provavelmente difere do de um teste típico de 'MitoStress', uma série de modelos de planilha foram desenvolvidos para ajudar a analisar esses conjuntos de dados, específicos para culturas de células 3D e serão fornecidos mediante solicitação. Esses arquivos de modelo de dados fornecerão dados sobre os principais parâmetros respiratórios mitocondriais detalhados e explicados na Figura 2. - Para analisar os dados, exporte os dados como um relatório de planilha do software do controlador WAVE e use um modelo de planilha independente para análise.
- Para exportar dados para um dos geradores de análise automatizados, siga os comandos de exportação de dados no controlador WAVE e selecione o gerador de exportação que corresponde ao tipo de ensaio. Alternativamente, exporte o arquivo de dados e carregue-o para análises do Seahorse.

Figura 2: Descritores esquemáticos para parâmetros derivados de análises de dados de fluxo extracelular. Abreviação: OCR = taxa de consumo de oxigênio. Clique aqui para ver uma versão maior desta figura.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Para obter spheróides bem formados e compactos, cada linha celular foi otimizada individualmente para a densidade de semeadura e duração do cultivo (Figura 3). As linhas celulares A549, HepG2/C3A e SK-OV-3 inicialmente formaram agregados soltos que não progrediram para esferoides redondos com perímetros claramente definidos até depois de 7 dias na cultura. Por outro lado, as células MCF-7 podem formar esferoides dentro de 3 dias. Houve uma clara correlação entre a densidade inicial de semeadura celular e o volume esferoide após o período de cultura para todos os modelos esferoides. O tamanho e a morfologia esferoides foram otimizados para a densidade de semeadura. A morfologia e a circularidade começaram a diminuir com o aumento do tamanho do esferoide em todos os modelos. As estratégias de semeadura para linhas celulares foram otimizadas em 4 × 103 células/poço para células A549 e SK-OV-3; As células HepG2/C3A foram previamente otimizadas em outros lugares para 1 × 103 células/bem, e as células MCF-7 foram usadas em 4 × 103 células/bem em todos os ensaios. Em estratégias otimizadas de semeadura, o volume de spheroid ficou entre 5,46 × 107 μm3 (SK-OV-3) e 1,45 × 108 μm3 (A549) (Figura 3B). Todos os tipos de esferoides apresentaram correlação linear entre a densidade inicial de semeadura e o volume esferoide onde A549 e HepG2/C3A tinham valores R2 de 0,957 e 0,947, respectivamente. Os volumes de spheroid MCF-7 e SK-OV-3 apresentaram maior correlação com a densidade inicial de semeadura, R2=0,977 (Figura 3A).
A circularidade esferoide foi calculada usando planimetria de imagem dentro do software de análise FIJI usando os diâmetros spheroid longos e curtos. Simetria esferoide perfeita teve circularidade = 1,0; desvio de 1,0 indicou perda de circularidade (Figura 3C). A circularidade foi maior nos esferoides MCF-7 do que em outros modelos onde a circularidade foi mantida entre 0,83 e 0,9 em todas as densidades de semeadura. Em comparação, o perímetro externo dos esferoides SK-OV-3 não foi tão claramente definido, e o volume de esferoides foi significativamente menor mesmo após 7 dias na cultura, produzindo esferoides com uma circularidade máxima de 0,61 a uma densidade de semeadura de 4 × 103/bem. As células hepG2/C3A também foram encontradas para formar esferoides apertados e bem formados com uma morfologia uniforme em toda a área de superfície de todos os esferoides, com circularidade mantida em 0,79 para células semeadas a 1 × 103 células/bem. As células A549 pareciam seguir uma tendência em que a circularidade e a morfologia e a morfologia e a morfologia esferoides foram aumentadas com densidade de semeadura; no entanto, a circularidade não foi superior a 0,63 na densidade utilizada nesses experimentos.
A respiração mitocondrial basal foi calculada como OCR medido a partir de esferoides semeados em 1 × 103, 2 × 103, 4 × 103, ou 8 × 103 células/poço em microplacas de cultura de spheroid de apego ultralow (Figura 3D). Para todos os tipos de esferoides, o OCR aumentou com o tamanho esferoide e foi linearmente correlacionado ao volume esferoide com R2 mais alto em esferoides MCF-7 a 0,988 e menor em spheroids SK-OV-3 em 0,744 (Figura 3E). O OCR medido foi estatisticamente diferente entre todos os grupos experimentais. A549 teve o OCR mais baixo, alcançando apenas 18 pmol/min/well no maior tamanho esferoide (Figura 3D). Por outro lado, os spheroids MCF-7 produziram um OCR semelhante no menor tamanho esferoide após apenas 3 dias na cultura, atingindo um OCR máximo de linha de base de 53 pmol/min/well para o maior tamanho esferoide (Figura 3D). O HepG2/C3A produziu dados OCR altamente consistentes com tamanho e morfologia e tamanho eféride. Em esferoides HepG2/C3A semeados de 1 × 103 células/bem, o OCR de linha de base atingiu uma média de 15 pmol/min/well, aumentando para um máximo de 52 pmol/min/well nos maiores esferoides (Figura 3D). OCR em esferoides SK-OV-3 só foi significativo entre os esferoides cultivados de 4 × 103 células/poço e 8 × 103 células/bem, com pouca diferença vista em esferoides cultivados entre 1 × 103, 2 × 103, ou 4 ×103 células. Apesar das diferenças de tamanho, os dados de OCR foram altamente semelhantes entre os esferoides HepG2/C3A e MCF-7 em todos os pontos de tamanho. Em relação ao tamanho do esferoide (μm3), o OCR de linha de base por esferoides MCF-7 era comparável ao dos esferoides HepG2/C3A cultivados ao longo de 7 dias a partir de 1.000 células por poço.

Figura 3: Parâmetros de crescimento esferoide determinam a respiração mitocondrial da linha de base. (A) A549, HepG2/C3A, Os spheroids MCF-7 e SK-OV-3 foram otimizados para a densidade de semeadura e seu crescimento monitorado em 1 × 103, 2 × 103, 4 × 103 e 8 × 103 células/bem em cada um dos fotomicrógrafos, da esquerda superior para a direita inferior, respectivamente; barras de escala = 500 μm. (B) O tamanho do esferoide foi calculado utilizando dados planimétricos de fotomicógrafos coletados e comparados com a estatística de correlação de Pearson; as linhas pontilhadas representam a distribuição de 95% do intervalo de confiança. (C) A morfologia esferoide foi comparada pelo cálculo da circularidade. (D) OCR foi medido em 5x, após o qual a rotenona-antimicina A foi adicionada para responder pela taxa respiratória não-iamtocondrial usando o analisador Agilent Seahorse XFe96. Ocrbasal medido como OCR - OCRr/a foi comparado entre densidade de semeadura (D) e volume esferoide (E). Os dados são médias ± SEM de 5-8 bem replica tipo esferoide per per e densidade de semeadura celular. Abreviaturas: OCR = taxa de consumo de oxigênio; OCRbasal = respiração mitocondrial basal; OCRr/a = OCR após a adição de rotenone-antimicina A. Clique aqui para ver uma versão maior desta figura.
A concentração e o curso de tempo para exposição a moduladores respiratórios na análise XF é um passo crítico na otimização do ensaio. Os compostos moduladores respiratórios-oligomicina, BAM15, uma mistura de rotenona-antimicina A-ou um controle de veículo DMSO foram sequencialmente injetados através das portas de injeção do cartucho de sensor em poços de microplaca contendo os esferoides MCF-7 (Figura 4A). Quatro ciclos de medição foram concluídos para determinar o OCR basal médio de 30-40 pmol/min/well para todos os grupos amostrais. Durante o restante da duração do ensaio, os moduladores respiratórios foram adicionados sequencialmente a cada 5 ciclos de medição para alcançar concentrações finais de poços de 0,5 μM (0,5 μg/mL oligomicina) após a injeção 1; 2,0 μM (2 μg/mL oligomicina) após a injeção 2; 5 μM (5 μg/mL oligomicina) após a injeção 3; e, finalmente, uma concentração máxima de poço de 11 μM (11 μg/mL oligomicina) após a quarta adição sequencial. Os esferoides MCF-7 não responderam ao controle do veículo durante todo o experimento (Figura 4A). O OCR basal imediatamente mudou após a primeira injeção de cada composto respectivo na menor concentração de 0,5 μM ou 0,5 μg/mL oligomicina (Figura 4B). OCR em esferoides MCF-7 foi reduzido com oligomicina de 41 pmol/min/well para 23 pmol/min/well após 5 ciclos de medição após a primeira injeção de 0,5 μg/mL (Figura 4B).
Em resposta a 0,5 μm BAM15, o OCR foi aumentado de 33 para 41 pmol/min/bem antes da segunda injeção (Figura 4C). Comparativamente, a combinação de rotenona mais antimicina A baixou o OCR de 37 para 13 pmol/min/bem antes da segunda injeção (Figura 4D). Traços cinéticos revelaram ainda uma diminuição linear constante (oligomicina e rotenona-antimicina A) ou aumento (BAM15) em OCR. Para todos os regimes de dosagem composta, um OCR de estado estável foi alcançado dentro de 10-12 ciclos completos de medição (60-72 min) em uma concentração total de poços de 2 μM BAM15, 2 μM rotenona, 2 μM antimicina A e 2 μg/mL oligomicina (Figura 4A). A taxa de consumo de oxigênio atingiu um patamar de estado estável em ~19 pmol/min/well (oligomicina), 52 pmol/min/well (BAM15) e 10 pmol/min/well (rotenona-antimicina A) (Figura 4A). Aumento da concentração composta de oligomicina, BAM15, ou rotenona mais antimicina A ainda não teve efeito óbvio sobre o OCR, que permaneceu constante durante todo o restante do ensaio. Esses dados demonstram que tanto a concentração composta quanto o curso de tempo de exposição a compostos moduladores respiratórios devem ser considerados para otimização de ensaios ao usar esferoides 3D.

Figura 4: Titulação de compostos moduladores respiratórios como um passo importante para otimizar a análise de fluxo extracelular. (A) Os esferoides MCF-7 foram semeados em 4 × 103 células/poço e cultivados ao longo de 3 dias antes de serem colocados em poços de um microplaca de ensaio esferoide contendo XF RPMI e sondados para moduladores mitocondriais OCR ± usando o analisador XFe96. OCR foi medido em 5x, após o qual foram adicionadas titulações de controle de veículo, oligomicina (B), BAM15 (C) ou antimicina rotenone A (D) para inibir a distease ATP mitocondrial, determinar a capacidade respiratória máxima ou estabelecer taxa respiratória não-almitocondrial, respectivamente. A concentração de cada modulador mitocondrial foi aumentada ao longo de 4 estratégias individuais de injeção de titulação (0,5 μM, 1,5 μM, 3 μM e 6 μM; unidades para oligomicina são μg/mL) para determinar o OCR de estado estável máximo em resposta à concentração composta ideal. OCR foi medido para 5 ciclos de medição entre cada injeção. Os dados são médias ± SEM de 5 a 8 réplicas de poços individuais. Abreviação: OCR = taxa de consumo de oxigênio. Clique aqui para ver uma versão maior desta figura.
Um dos principais benefícios da tecnologia XF é a capacidade de sondar a função mitocondrial em células e tecidos intactos. Para examinar aspectos específicos da função mitocondrial em células e tecidos, moduladores mitocondriais são adicionados sequencialmente aos poços da amostra microplaca através das 4 portas de injeção disponíveis no cartucho do sensor. A sequência típica de moduladores usados para sondar parâmetros mitocondriais em ensaios XF são a oligomiecina, um protonophore (por exemplo, FCCP ou BAM15), e uma combinação de rotenone mais antimicina A, que são adicionados sequencialmente para inibir a disteligeste atp mitocondrial, determinar capacidade respiratória máxima e corrigir a taxa respiratória não-peritrudária, respectivamente. Esta sequência típica de adições moduladoras é chamada de teste MitoStress pelo fabricante de tecnologia de ensaio. Dado que a oligomicina pode inibir a respiração estimulada pelo desacoplador em algumas monocamadas de células20, examinamos isso com esferoides 3D derivados do câncer medindo OCR (OCRmax) estimulado sem acoplamento antes (único) e depois da injeção de oligomicina (Figura 5A-D) (Figura 5A-D). OCRmax não foi significativamente limitado pela adição de oligomicina em esferoides formados a partir de HEPG2/C3A ou SK-OV-3 (Figura 5E e Figura 5G). No entanto, o OCRmax foi significativamente reduzido em esferoides A549 e MCF-7 após uma injeção sequencial de BAM15 após oligomicina em comparação com o OCRmax obtido a partir de uma única injeção de BAM15 (Figura 5F e Figura 5H). Salvo ade ser conhecido de outra forma, recomenda-se, portanto, usar poços separados para tratar com oligomicina e uncoupler, com uma adição final de rotenona e antimicina A ao explorar o metabolismo de energia mitocondrial de esferoides 3D. Esta abordagem ainda permite o cálculo de todos os parâmetros mitocondriais como em um teste típico de MitoStress onde os compostos são adicionados sequencialmente.

Figura 5: Injeção única ou sequencial de compostos respiratórios mitocondriais. Eferóides derivados de células cancerígenas de MCF-7, HEPG2/C3A, SK-OV-3 e A549 foram colocados em poços de uma microplaca esferoide XFe96 em XF RPMI e sondados para OCR usando o analisador Agilent Seahorse XFe96. OCR foi medido 5x, após o qual 2 μg/mL oligomicina (porta de injeção A: traço verde) ou 5 μM BAM15 (porta de injeção A: traço azul ou porta de injeção B: traço verde) para inibir a synthase ATP mitocondrial e determinar a capacidade respiratória máxima, respectivamente. Os dados do OCR cinético são expressos como % basal (A-D). A capacidade respiratória máxima (OCRmax) foi calculada como um fator de OCR basal pela equação: OCRmax = OCRBAM15 / OCRbasal. OCRmax foi obtido a partir de médias OCR em ciclos de medição 8-10 pós injeção BAM15 com (barras verdes) e sem (barras azuis) oligomicina. Os dados são médias ± SEM de 3 a 8 réplicas de poços individuais em toda a microplacão de ensaio esferoide. Abreviaturas: OCR = taxa de consumo de oxigênio. Clique aqui para ver uma versão maior desta figura.
Utilizando densidades ideais de semeadura celular, concentrações compostas, estratégia de injeção e período de ciclo de medição determinados nesses experimentos de otimização (Tabela 3), desenvolvemos um protocolo detalhado para sondar com precisão a respiração mitocondrial basal: OCRbasal (Figura 6A), respiração de fósforo ADP( Respiração OCRADP (Figura 6B), respiração de vazamento: OCRomy (Figura 6C), eficiência de acoplamento de ADP (6D) ), capacidade respiratória máxima: OCRmax (Figura 6E) e capacidade respiratória sobressalente: OCR(Figura 6F) utilizando esferoides 3D derivados do câncer.

Figura 6: Sondando OCR com tecnologia XF para estabelecer o metabolismo de energia mitocondrial de esferoides derivados do câncer. Eferóides derivados de células cancerígenas de MCF-7, HEPG2/C3A, SK-OV-3 e A549 foram colocados em poços de uma microplaca de ensaio esferoide em XF RPMI e sondados para OCR usando o analisador Agilent Seahorse XFe96. OCR foi medido 5x, após o qual 2 μg/mL oligomicina, ou 5 μM BAM15, e RA foi adicionado para inibir a distease ATP mitocondrial, determinar a capacidade respiratória máxima e calcular a taxa respiratória nãomitocondrial, respectivamente. (A) A respiração mitocondrial basal (OCRbasal) foi calculada como a média do OCR dos 3 ciclos de medição antes da injeção da porta A. (B) A eficiência do acoplamento da fosforilação oxidativa foi aproximada por expressar ovazamento OCRADP (OCRbasal-OCR) como uma porcentagem dobasal OCR. (C) A respiração de fosforilação ADP (OCRADP) foi medida como OCR sensível à oligomicina, calculada a partir do OCR médio nos ciclos de medição 11-13 antes da injeção BAM15. (D) A respiraçãoomy de vazamento (vazamento de OCR) foi medida como OCR insensível à oligomicina, calculada a partir da média de OCR em ciclos de medição 11-13. (E) A capacidade respiratória máxima (OCRmax) foi medida como o OCR máximo médio medido após a injeção BAM15. (F) A capacidade respiratória sobressalente foi calculada expressando o OCRsobressal (OCRmax - OCRbasal)como uma porcentagem de basal OCR. A injeção OCR após a injeção de rotenona-antimicina A (OCRr/a) foi subtraída de todas as taxas para corrigir para OCR não-mitocondrial. Os dados são médias ± SEM de 3-8 réplicas de poços individuais através da placa esferoide XFe96. Abreviaturas: OCR = taxa de consumo de oxigênio; RA = 2 μM rotenone-2 μM antimicina A. Clique aqui para ver uma versão maior desta figura.
Os esferoides MCF-7 cultivados de 4 × 103 células/bem ao longo de 3 dias foram usados como modelo para determinar a melhor transferência, colocação e análise dentro de microplacas de ensaio esferoide. Utilizando as dimensões fornecidas para a microplacão esferoide do fabricante, a superfície do poço foi dividida em três áreas de zona para a colocação ideal de esferoide (Figura 7A), onde a zona 1 foi destacada como a zona ideal no centro do poço. Com tubos cuidadosos usando pontas de pipeta de orifício largo, os esferoides foram transferidos para as placas esferoides e distribuídos aleatoriamente pelas superfícies do poço por elução gravitacional (Figura 7B). Onde os esferoides foram cuidadosamente transferidos usando elução gravitacional, a maioria dos esferoides poderia ser tipicamente encontrada nas zonas 1-2 da microplacão, usando as técnicas recomendadas de transferência do fabricante. Onde os esferoides eram forçados a sair da ponta da pipeta por aspiração, os esferoides eram frequentemente colocados além dessas zonas e não podiam ser vistos usando microscopia.
Para comparar as posições de colocação esferoides, os esferoides MCF-7 foram transferidos para as microplapes de ensaio esferoide em zonas designadas 1-3 ou fora da zona (Figura 7A). Estes 4 poços foram rastreados através de um experimento cinético OCR na linha de base e após a adição de oligomicina, BAM15, ou rotenone-antimicina A (Figura 7C). OCR foi calculado a partir da média de três leituras de ciclo antes de cada injeção (Figura 7B). OCR foi medido cineticamente mais de 200 min nos 4 poços selecionados (Figura 7C) e corrigidos na linha de base (Figura 7D). Onde os spheróides foram colocados na zona 3 ou fora da zona, os OCRs de linha de base foram significativamente inferiores aos esferoides colocados nas zonas 1 e 2 (Figura 7C). Os efeitos dos compostos respiratórios oligomicina, BAM15 e rotenona-antimicina A também diferiram dramaticamente entre os esferoides colocados nas zonas 1 e 2 em comparação com as regiões da zona 3 e fora da zona. Um aumento no OCR foi visto com oligomicina em esferoides colocados na zona 3 ou fora da zona (Figura 7E). Além disso, os esferoides colocados na zona 3 ou fora da zona experimentaram uma resposta excessivamente alta ao BAM15 com OCR superior à linha de base após a injeção de rotenona-antimicina A (Figura 7E). Apesar de um aumento de quase duas vezes no OCR basal (Figura 7C) com esferoides colocados na zona 2 versus zona 1, as mudanças de dobra em resposta a todos os compostos respiratórios foram muito semelhantes (Figura 7E), sugerindo que diferenças no OCR basal entre esferoides colocados nas zonas 1 ou 2 dificilmente serão resultado de colocação dentro do poço.

Figura 7: A colocação de esferoides dentro da microplaca de ensaio esferoide dita efeitos basais de OCR e modulador mitocondrial usando a tecnologia XF. Os esferoides MCF-7 foram semeados em 4 × 103 células/bem e cultivados ao longo de 3 dias antes de serem colocados nos poços da microplaca esferoide contendo XF RPMI e sondados para moduladores mitocondriais OCR ± usando o analisador Agilent Seahorse XFe96. (A) Fotomicrogramas de posições de zona esferoide em microplacões de ensaio esferoide após a duração do ensaio; barra de escala = 500 μm e OCR capturados de poços correspondentes ao longo do tempo expressos como pmol/min-1/well-1 (B) ou % basal (C). (D) Efeitos moduladores mitocondriais de esferoides MCF-7 colocados em diferentes zonas dentro da microplaca de ensaio esferoide; dados expressos como mudança de dobra de basal. (E) Exemplo de traço cinético destacando quais medidas de dados OCR (círculos vermelhos) são usadas para calcular a resposta de cada modulador mitocondrial para dados apresentados em E. Os dados mostrados são de respostas individuais de poços. Abreviação: OCR = taxa de consumo de oxigênio. Clique aqui para ver uma versão maior desta figura.
Os critérios de seleção para o fundo são de alta importância; o uso de poços mais externos para correção de fundo não é representativo de todos os poços de microplacão, o que pode levar a suposições incorretas de dados sendo desenhadas e conclusões de dados errôneas devido a efeitos de borda em toda a microplato esferoide. Para avaliar essa observação, foram utilizados spheroids MCF-7 para comparar os procedimentos de correção de ensaios para derivar valores de OCR em resposta à adição de um controle veicular, oligomicina, BAM15 ou rotenone-antimicina A (Figura 8). Todos os compostos respiratórios produziram os perfis OCR cinéticos esperados para os compostos selecionados, revelando uma taxa média de respiração basal estável média de 20-30 pmol/min/well (Figura 8A). No entanto, quando os dados de ensaio foram analisados utilizando os poços mais externos para correção da temperatura de fundo (A1, A12, H1 e H12), os valores revelados para OCR após a adição de compostos respiratórios foram especialmente baixos; OCR rendeu valores negativos para rotenona-antimicina A. Em resposta a essas observações, a análise alternativa foi realizada utilizando-se uma série de poços vazios, distribuídos aleatoriamente através da microplapão esferoide, como poços de correção da temperatura de fundo (Figura 8B). Quando foi aplicada a correção de fundo alternativa, todos os efeitos compostos relativos no OCR foram os mesmos em ambos os conjuntos de análise; no entanto, os valores absolutos de OCR aumentaram aproximadamente 10 pmol/min/well (Figura 8). Esses dados destacam o poder e a importância da correção da temperatura em segundo plano em microplapes de ensaio e enfatizam a importância da otimização do usuário para a análise de XF.

Figura 8: Seleção aleatória de poços para correção de fundo para melhorar o controle de gradientes de temperatura em toda a microplacão de ensaio esferoide. Dados OCR extrapolados da Figura 2A usando poços recomendados para correção de fundo (A) versus poços atribuídos aleatoriamente para correção de fundo (B). Abreviação: OCR = taxa de consumo de oxigênio. Clique aqui para ver uma versão maior desta figura.
Ao contrário das monocamadas celulares, os esferoides representam uma agregação heterogênea das células em um espaço 3D e, portanto, requerem uma consideração minuciosa em relação à análise, particularmente quando normalizam esses dados. Este artigo apresenta três abordagens para normalizar os dados XF adquiridos dos esferoides MCF-7 (Figura 9). Quando não normalizado, o OCR correlaciona positivamente (R2 = 0,98) com o tamanho esferoide (determinado pela densidade inicial de semeadura celular) significativamente quando comparado estatisticamente com o coeficiente de correlação de Pearson, P = 0,0057 (Figura 9A). Esta relação linear é reduzida quando o OCR é normalizado à densidade inicial de semeadura celular (R2 = 0,78) e não se correlaciona significativamente com o tamanho do esferoide (P = 0,117, Figura 9B). Este também é o caso quando normalizado para volume esferoide (R2 = 0,77; Coeficiente de correlação de Pearson P = 0,120, Figura 9C) e teor de dsDNA nuclear (R2 = 0,58; Coeficiente de correlação de Pearson P= 0,233, Figura 9D). Esses dados destacam a importância da normalização dos dados XF ao sondar o metabolismo mitocondrial dos esferoides, especialmente se forem de tamanhos diferentes.

Figura 9: Normalização de dados de fluxo extracelular adquiridos de esferoides celulares. (A) Os dados de OCR bruto foram obtidos do MCF-7 cultivados ao longo de 3 dias e traçados usando o modelo de Pearson para obter um coeficiente de correlação entre densidade de sementes esferoides e OCR; Valor de P definido em 0,05. (B) Os dados de OCR bruto foram normalizados em relação à densidade inicial de semeadura esferoide; (C) Volume esferoide MCF-7 obtido a partir da planimetria de microscopia; e (D) conteúdo de DNA nuclear ds comparado usando o coeficiente de correlação de Pearson. Abreviaturas: OCR = taxa de consumo de oxigênio; ds DNA = DNA de dupla mente. Clique aqui para ver uma versão maior desta figura.
| Linha celular | Densidade de semeadura (bem) | Crescimento esferoide (dias) | Volume esferoide final (μM3) | Basal OCR (pmolO2/min/well) | Sensibilidade para OCR basal encontrado (SIM/NÃO) |
| SKOV | 1000 | 5 | 9.52E+06 | 28 ± 3,5 | Sim |
| SKOV | 2000 | 5 | 2.38E+07 | 26 ± 1,4 | Sim |
| SKOV | 4000 | 5 | 4.92E+07 | 36 ± 3.1 | Sim |
| SKOV | 8000 | 5 | 1.11E+08 | 50 ± 7.9 | Sim |
| HepG2 | 1000 | 5 | 1.11E+07 | 15 ± 0,7 | Não |
| HepG2 | 2000 | 5 | 2.88E+07 | 23 ± 1,8 | Sim |
| HepG2 | 4000 | 5 | 5.46E+07 | 31 ± 1,7 | Sim |
| HepG2 | 8000 | 5 | 1.21E+08 | 52 ± 2,8 | Sim |
| A549 | 1000 | 5 | 2.11E+07 | 30 ± 2,5 | Sim |
| A549 | 2000 | 5 | 3.57E+07 | 41 ± 1,6 | Sim |
| A549 | 4000 | 5 | 6.93E+07 | 53 ± 7.2 | Sim |
| A549 | 8000 | 5 | 1.44E+08 | 65 ± 8.4 | Sim |
| MCF-7 | 1000 | 3 | 1.60E+07 | 29 ± 0,8 | Sim |
| MCF-7 | 2000 | 3 | 2.52E+07 | 37 ± 1,7 | Sim |
| MCF-7 | 4000 | 3 | 6.00E+07 | 46 ± 1,7 | Sim |
| MCF-7 | 8000 | 3 | 1.06E+08 | 66 ± 2,9 | Sim |
Tabela 4: Parâmetros otimizados para determinar medições de OCR basais em esferoides 3D únicos. Abreviação: OCR = taxa de consumo de oxigênio.
Arquivo Suplementar 1: Análise do tamanho e volume esferoides. Clique aqui para baixar este Arquivo.
Arquivo Suplementar 2: Quantificação de DNA de dupla derivada de esferoides na microplacão esferoide. Clique aqui para baixar este Arquivo.
Arquivo Suplementar 3: Recomendações para o número de réplicas necessárias para obter conjuntos de dados de ensaio XF confiáveis. Clique aqui para baixar este Arquivo.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Principais descobertas e saídas
Este artigo fornece um protocolo detalhado para sondar o metabolismo de energia mitocondrial de esferoides 3D únicos usando uma série de linhas celulares derivadas do câncer com o XFe96 XF Analyzer. Um método é desenvolvido e descrito para o rápido cultivo de Esferoides celulares A549, HepG2/C3A, MCF7 e SK-OV-3 usando tecnologias repelentes celulares para agregação forçada. Este protocolo aborda muitas considerações sobre a sondagem do metabolismo esferoide com a tecnologia XF, incluindo (1) otimização de protocolos de cultura esferoide e o manuseio e transferência de esferoides em microplacas de ensaio spheróides específicos do fabricante de tecnologia de seus vasos originais de cultivo; (2) a concentração de compostos respiratórios a serem utilizados e a dependência do tempo de penetração composta; (3) estratégias de injeção a serem utilizadas; e (4) formas de normalizar os dados entre grupos experimentais. Todas essas considerações foram examinadas no artigo atual e são discutidas em mais detalhes abaixo. Esses métodos são apresentados como abordagens simplificadas para gerar dados consistentes de fluxo de oxigênio metabólico usando esferoides 3D únicos com o analisador XFe96 Flux. Esta abordagem experimental pode ser usada como ponto de partida e rubrica para uso em outros modelos esferoides que são facilmente implementados dentro de um ambiente básico de laboratório.
Considerações
Crescimento, tamanho e sensibilidade da tecnologia XF
Para estabelecer dados reprodutíveis com a tecnologia XF, é essencial caracterizar e otimizar o ensaio para o modelo específico. Esta abordagem é relativamente simples em uma monocamada básica de células; no entanto, isso apresenta desafios adicionais ao cultivar células como esferoides 3D. Durante os experimentos aqui apresentados, o meio RPMI do fabricante foi complementado no momento do uso. Embora se notie que algumas linhas de células, ou seja, HepG2/C3A, foram cultivadas em meio de crescimento DMEM, durante esses ensaios relativamente curtos (~3-5 h), a substituição por formulações de RPMI-DMEM teve impacto limitado na análise do XF. A formulação das duas mídias são muito semelhantes, e os usuários poderiam 'sintonizar' a mídia Seahorse RPMI para combinar com a matriz de seus meios de cultura celular através da suplementação, por exemplo, aumento da glicose, adição adicional de fontes de carboidratos. Fundamental para a formulação final de todos os buffers e meios XF é a ausência de fenol-vermelho, que provavelmente interferirá com as sondas fluorescentes dentro da placa do cartucho da sonda XF, e bicarbonato de sódio, o que levará à alcalinidade devido à falta de tampão de CO2 presente nas incubadoras de cultura celular. Outros meios de comunicação e buffers podem ser comprados e/ou feitos internamente. Por exemplo, o buffer Desferteiro Ringer HEPES é um buffer simples que pode ser usado para avaliar a respiração em muitas células diferentes, incluindo modelos esferoides. No entanto, os usuários de ensaios XF devem observar que uma mudança no meio/tampão e sua suplementação pode alterar sua capacidade global de buffering. Isso é particularmente preocupante quando os usuários podem estar interessados em medir o ECAR, no qual o fator tampão do meio precisa ser avaliado para permitir a transformação do ECAR para a taxa de efflux de prótons (PER).
Como o OCR celular medido pela tecnologia XF é proporcional à densidade celular quando o número de células no poço está dentro da sensibilidade do sistema, era importante investigar essa relação usando esferoides 3D únicos. Sondando OCR de spheróides 3D únicos cultivados a partir de 4 diferentes linhas de células cancerosas semeadas em densidades de 1.000, 2.000, 4.000, ou 8.000 células por poço, mostramos que o analisador XFe96 é sensível o suficiente para detectar alterações na taxa de respiração mitocondrial entre esferoides 3D cultivados a partir de diferentes densidades de semeadura celular (Figura 3). Mostramos que a faixa ideal de densidade de semeadura celular e, portanto, o volume esferoide para a formação de esferoides 3D para sondagem de OCR, diferem dependendo do tipo celular. Isso é mostrado pela relação linear entre OCR e densidade de semeadura ou volume esferoide (Figura 3). Para as células A549 e HepG2/C3A, a densidade ideal de semeadura para sensibilidade ao OCR foi entre 1.000 e 8.000 células/bem; foram 2.000-8.000 células/poço para mcf-7 e 4.000-8.000 células/poço para células SK-OV-3. Esses dados demonstram que a otimização do tamanho do esferoide é de particular importância ao avaliar o OCR usando a tecnologia XF.
Considerações sobre volumes esferoides mínimos e máximos e OCR basal
Em geral, sempre haverá limites mínimos e máximos para parâmetros OCR mensuráveis recomendados pela fabricação para esses experimentos. Para o analisador XFe96, o OCR basal entre 20 pmol O2/min/well e 200 pmol O2/min/well são os limites inferior e superior, respectivamente. Este é o caso de células monocamadas e esferoides, e onde o modelo experimental está dentro dessa faixa dinâmica de OCR dependerá da quantidade de material biológico disponível, por exemplo, o número de células como monocamadas ou o tamanho de esferoides. Consulte a Tabela 4 para um exemplo de como os limiares de OCR foram alcançados pelos modelos esferoides utilizados aqui. Pode ser prudente verificar o nível de oxigênio dentro do poço para o qual esses dados também estão disponíveis a partir dessas medidas como os dados de nível. Isso deve ser visto rotineiramente a partir de cada experimento para fins de controle de qualidade. Se houver esgotamento de oxigênio no poço, isso ficará evidente dentro dos dados. Caso este seja o caso, pode ser necessário ajustar os ciclos de medição dentro do experimento; por exemplo, aumentar a etapa de mistura de tal forma que o nível de oxigênio no poço seja recuperado antes do próximo período de medição dentro do ciclo de medição. Embora possível, descobrimos que isso é muito improvável para experimentos de esferoides únicos usando as linhas celulares descritas.
Escolha de unacouplers mitocondriais para ensaios de fluxo extracelular
Ionóforos de próton, como cianeto de carbonil 4-(trifluorometoxy) fenilhydrazone (FCCP)21, a hidrato de cianeto de ciriida de carbonil (CCCP)22 ou BAM1523, são potentes produtos químicos de pequenas moléculas capazes de interromper o gradiente de próton eletroquímico através de membranas mitocondriais, inibindo a produção de ATP e, finalmente, desacoplando a respiração mitocondrial24 . Novas moléculas pequenas continuam a ser desenvolvidas para esses fins, particularmente no tratamento da doença metabólica 25,26,27; referem-se a duas excelentes avaliações28,29. Por outro lado, o desacoplamento da respiração oxidativa tem sido associado com toxicidade fora do alvo indesejável30. No entanto, dentro de ensaios celulares in vitro, a molécula FCCP despolariza o potencial da membrana mitocondrial e exerce efeitos fora do alvo, como a despolarização da membrana plasmática, interrompendo o fluxo de íons NA+ 31; interferência com o processamento de proteína celular32, e até mesmo induzir senescência celular33. O BAM15 foi originalmente introduzido em 2013 como um uncoupler mitocondrial com influência mínima nas membranas plasmáticas23, com atividade prótonofórica na faixa de micromolar em células inteiras e faixa de nanomolar em mitocôndrias isoladas 23,34.
Dada a potência do FCCP na despolarização da membrana plasmática, BAM15 é um prótonophore mais confiável para desacoplamento da respiração em células inteiras intactas em ensaios de fluxo extracelular. Embora a FCCP e sua contraparte, CCCP, tenham sido usadas por mais de 50 anos para avaliar capacidades respiratórias máximas e continuarem a ser amplamente utilizadas em estudos de XF, o uso dessas pequenas moléculas muitas vezes subestima a capacidade metabólica mitocondrial e celular. Isso está em parte ligado à razão pela qual tantas publicações usando a tecnologia XF caem na armadilha de relatar capacidades respiratórias negativas ou subestimam as verdadeiras capacidades respiratórias mitocondriais quando a FCCP é usada. A potência adicional do FCCP em células e tecidos intactos muitas vezes leva à função mitocondrial comprometida, e as células podem lutar para operar adequadamente para sustentar uma capacidade respiratória máxima em vários ciclos de medição após sua adição, mesmo em concentrações muito baixas35. Portanto, a resposta das células ao FCCP pode ser encontrada em muitos estudos para cair após o período inicial do ciclo de medição. Embora a FCCP tenha sido usada rotineiramente para análise de XF, o BAM15 é usado preferencialmente em casos envolvendo células inteiras ou modelos esferoides, uma vez que pode manter uma capacidade respiratória máxima em mitocôndrias totalmente despolarizadas em concentrações de até 10 μM 3. Além disso, o BAM15 induz efeitos na acidificação extracelular, que coincide com a oxidação de nutrientes através da hidratação de CO2 para formar HCO3- e H+ em maior medida que o FCCP3. No entanto, no caso de mitocôndrias isoladas e células permeabilizadas, qualquer um desses desacopladores deve executar tão bem quanto BAM15 para desacoplamento mitocondrial se titulado na concentração correta.
Cinética da penetração composta e ensaio de ciclismo
As concentrações, penetração e perfis cinéticos de compostos químicos usados para realizar um teste típico de MitoStress com esferoides 3D usando o analisador XF são mais complexos de abordar. Dado que os esferoides apresentam estrutura 3D, a penetração de moléculas através do diâmetro do esferoide é um processo infinitamente mais complexo do que através de monocamadas celulares. Por exemplo, a penetração cinética e, portanto, a sensibilidade ao sorafenib quimioterápico foi determinada pela idade esferoide e, portanto, pelo tamanho em um modelo esferoide HepG236. A capacidade de produtos químicos de pequenas moléculas (por exemplo, drogas, nanopartículas) para atingir um alvo biológico depende de vários fatores subjacentes, incluindo a complexidade do sistema a ser penetrado dinamicamente e difundido através de37,38. Isso é particularmente verdadeiro para drogas que visam o tecido tumoral39. Semelhante ao alvo do tumor no contexto de um esferoide 3D, tamanho, compactação e outras respostas fenotípicas, como a expressão de proteínas de transporte de drogas podem reger o tempo de penetração e a concentração de um composto necessário para obter uma resposta biológica.
Neste protocolo, abordamos a questão em torno do tempo de penetração e da concentração de pequenas moléculas em resposta ao inibidor de sinóase ATP oligomiina, o protonophore e mitocondrial uncoupler BAM15, e a combinação dos inibidores complexos I e Complexo III rotenona e antimicina A. Ao sondar o OCR de spheróides mcf-7 únicos expostos a múltiplas titulações desses compostos respiratórios comuns, demonstramos que a concentração ideal de cada composto necessário para induzir uma taxa respiratória de estado estável se enquadra em uma faixa semelhante à das células monocamadas (Figura 4). É importante, e diferente de suas contrapartes de monocamadas, é demonstrado que aumentar o número de ciclos de medição entre injeções é fundamental para alcançar um OCR de estado estável em esferoides 3D únicos. Esses dados destacam a importância da penetração composta e seus respectivos perfis cinéticos ao explorar parâmetros respiratórios mitocondriais de esferoides 3D utilizando essas abordagens. Utilizando propriedades de otimização esferoides, concentrações de compostos e tempos de ciclo de medição informados pelos dados apresentados na Figura 3, Figura 4 e Figura 5, foi estabelecido um teste mitostress validado para a sondagem de parâmetros específicos do metabolismo oxidativo mitocondrial em uma gama de esferoides 3D derivados do câncer (Figura 6). De importância, e como algumas linhas de células cancerígenas monocamadas40, a capacidade respiratória máxima (taxa de respiração estimulada não acoplada) de certos esferoides 3D derivados do câncer foi inibida pela oligomicina (Figura 5). Especificamente, os esferoides 3D cultivados a partir de células A549 ou MCF-7 mostraram uma taxa máxima de respiração significativamente menor quando desacoplados com BAM15 após a injeção de oligomicina em comparação com ser desacoplado por BAM15 sem oligomicina (Figura 5F e Figura 5H). Dado que esse efeito pode estar presente em outras culturas esferoides 3D, sugerimos que, a menos que um protocolo previamente validado seja empregado, a capacidade respiratória máxima em esferoides 3D deve ser estimada sem oligomicina.
Coleta simultânea de dados ECAR como medida de fluxo glicóltico em esferoides celulares
Como normalmente visto na literatura ou informação do fabricante de tecnologia, a taxa glicóltica de esferoides, medida como ECAR, é um parâmetro secundário que pode ser capturado ao lado do OCR. O cálculo do ECAR por si só não é um parâmetro útil ou significativo em qualquer experimento XF, pois não é corrigido para a capacidade de buffer do tampão de ensaio XF ou a adição de acidificação mitocondrial, que surge da hidratação de CO2 a HCO3- e H+. O ECAR só é perspicaz quando essas correções de dados são aplicadas, após as quais torna-se possível fornecer conclusões mais precisas sobre o fluxo glicóltico. Para corrigir a capacidade de buffer para gerar dados PER mais significativos, é preciso saber o volume da microcâmara para a microplaca esferoide. O fabricante não foi capaz de fornecer um volume real para isso com a microplacão e, portanto, os dados PER não podem ser determinados facilmente. De fato, embora essas medidas pudessem ser alcançadas empiricamente, isso estava além do escopo deste manuscrito. No entanto, com as correções apropriadas e conhecendo o volume da microcâmara para um determinado tamanho esferoide presente (por exemplo, obtendo uma medida de densidade esferoide) no poço, os dados ECAR se tornariam significativos, e os cálculos do PER glicóltico poderiam ser feitos. Assim, os dados de XF poderiam então ser mais informativos para investigar o metabolismo glicolítico e oxidativo em esferoides, mas apenas se esses parâmetros fossem considerados em profundidade.
Formação, manuseio, transferência e movimento de spheroid
Algumas linhas celulares são mais adequadas à formação de esferoides do que outras e podem não formar esferoides, por exemplo, as células cancerígenas ovarianas MCF-741,42 formam esferoides altamente circulares em comparação com outras linhas celulares (Figura 3). Como outro exemplo, as células cancerígenas pancreáticas Capan-1 têm sido mostradas para formar melhores esferoides do que Panc-1 ou BxPC343. Da mesma forma, as linhas de células cancerígenas hepáticas são conhecidas por terem habilidades variáveis para formar esferoides compactos 5,44, com uma alteração observada no fenótipo, como o metabolismo de drogas aprimorada ou a produção de albumina, como é o caso de HepG2 versus HepG2/C3A 9,45,46 ou HepaRG spheroids 17,47,48 . Portanto, os usuários devem otimizar as técnicas de cultivo de esferoides de acordo e realizar experimentos de titulação para determinar a densidade de semeadura ideal e o curso de tempo de cultivo. Além disso, a formulação e composição dos meios de ensaio têm sido demonstradas para impactar a formulação de spheroid, incluindo a adição de metilcelulose, muitas vezes adicionada à mídia para aumentar a viscosidade matricial 43,49,50. Assim, a composição média da célula ideal deve ser determinada empiricamente para todas as linhas celulares utilizadas.
O número de trocas médias ao longo da cultura esferoide é determinado pela linha celular utilizada. No entanto, normalmente, trocas médias de meio volume a cada 2-3 dias são aplicáveis na maioria dos casos para repor nutrientes. Utilizamos a abordagem agregada forçada para gerar esferoides 3D usando microplacos repelentes de células de fontes comercialmente disponíveis para desenvolvimento rápido e implantação de modelos esferoides em estudos de análises XF. No entanto, plataformas alternativas podem ser mais adequadas para gerar esferoides de outros tipos de células, por exemplo, abordagens de queda suspensa ou embutidas por matriz. Em laboratórios limitados a recursos, os usuários podem querer olhar para a técnica de sobreposição agarose-líquido para a formação de superfícies microplacáveisrepelente celular 51,52 para reduzir significativamente os custos econômicos das etapas iniciais de desenvolvimento do método esferoide. O movimento de esferoides entre os vasos culturais é necessário para realizar análises XF e outros ensaios a jusante. A facilidade de transferência é tipicamente ditada pelo tamanho esferoide e densidade geral. Recomendamos o uso de uma ponta de pipeta de orifício largo P200 ou P1000 para manter a integridade esferoide; Pontas de pipeta de menor furo correm o risco de interrupção mecânica do esferoide, que pode ser comprado de origem comercial ou, com cuidado, feito simplesmente aparando a extremidade da ponta pipeta para aumentar o orifício. No entanto, essa abordagem pode ser passível de introduzir furring ao plástico ao redor da extremidade da ponta, o que poderia causar interrupção mecânica durante o manuseio. O uso de uma backlight ou lightbox também é útil para o manuseio e observação de spheroid sob um microscópio de dissecção como um passo essencial para garantir a transferência bem sucedida de esferoides para a microplaca de ensaio esferoide. Além disso, a posição esferoide dentro do poço de um microplaca de ensaio esferoide é de particular importância e impacta diretamente o OCR e o efeito composto durante um teste típico de MitoStress (Figura 7), provavelmente devido à relação da posição esferoide e da sonda do sensor fluoroforos.
Poços de correção de fundo e controle de temperatura
O uso de ensaios à base de microplacos é uma abordagem amplamente utilizada em diversas áreas de pesquisa; no entanto, seu uso apresenta vários desafios práticos. Como é verdade em outras abordagens experimentais, particularmente aquelas que usam o formato de matriz 96 (ou maior), a geometria e o posicionamento de microplaca podem influenciar os gradientes de temperatura e troca de gás em toda a placa ao longo do tempo, muitas vezes referidos como "efeitos de borda" 53,54. Achamos o mesmo como verdade para a microplacão de ensaio esferoide. De acordo com as diretrizes e protocolos do fabricante, os poços de canto mais externos: A1, A12, H1 e H12 são sempre designados como poços de correção de fundo e controle de temperatura para o analisador XFe96. Por outro lado, com o formato de matriz de 24 poços, A1 e D6 são designados como poços de controle, ao lado de dois outros poços distribuídos uniformemente pelo meio da placa em B4 e C3. Ao realizar a análise de spheroid XF, encontramos desvio significativo nos dados inicialmente coletados usando a orientação do fabricante. Isso foi apesar da inclusão das etapas necessárias para garantir a pré-calibração do ensaio à temperatura e ao teor deCO2 antes de iniciar a aquisição de dados, muitas vezes produzindo valores negativos para OCR após a injeção de certos inibidores respiratórios (Figura 8).
Descobrimos que essas observações provavelmente devem-se a efeitos de borda em toda a microplapão de ensaio esferoide. Na Figura 8, descobrimos que redistribuindo poços de controle de fundo através da microplacão, os dados XF foram ajustados aproximadamente 2 vezes. Duas razões mais prováveis são (1) devido aos efeitos de evaporação nos poços de borda, resultando em uma câmara de volume total menor para a sonda XFe96 para amostrar, e (2) de equilíbrios de temperatura inadequados entre os poços designados para correção de fundo e poços de amostra, resultando em conjuntos de dados que mascaram ou inflam excessivamente OCR. Para evitar tais resultados, recomenda-se, portanto, especialmente no contexto da análise esferoide, que os usuários redistribuam poços designados para correção de fundo em toda a microplacão de ensaio esferoides e tomem as medidas necessárias para pré-equilibrar seu ensaio antes de adquirir dados XF.
Normalização dos dados
Além de fornecer um protocolo detalhado para sondar o metabolismo de energia mitocondrial de esferoides 3D únicos com tecnologia XF, este artigo também apresenta possíveis maneiras de normalizar os dados de taxa respiratória mitocondrial obtidos com esferoides 3D. Utilizando dados de taxa respiratória obtidos com esferoides MCF-7 cultivados em diferentes densidades de semeadura celular (Figura 3), apresentamos taxas respiratórias mitocondriais basais de esferoides MCF-7 de tamanho e diâmetro crescentes quando normalizados para densidade inicial de semeadura celular, volume esferoide e conteúdo dsDNA (Figura 9). O método de normalização adequado é primordial para a interpretação precisa dos conjuntos de dados XF, particularmente quando se compara a modelos esferoides 3D in vitro e diferentes tipos de células. A má normalização pode levar a resultados errôneos que simplesmente não podem ser comparados entre conjuntos de dados. O teor de proteína não é preferido para a normalização dos dados de XF esferoides, pois os pré-tratamentos podem impactar as taxas de síntese proteica sem efeito significativo na taxa respiratória. Além disso, quantidades significativas e inconsistentes de proteína podem se ligar a microplacões esferoides sobre a lise celular, introduzindo variação no teor de proteínas entre os poços. Isso pode ser ainda mais complicado nas análises de XF usando esferoides ou células nãoadherentes que requerem colas biomoleculares para se ligar, que podem conter proteínas.
Ao contrário do teor de proteína intracelular, o conteúdo de DNA nuclear é independente do tipo celular e é proporcional ao número celular (Figura 9D)-uma abordagem mais precisa e menos demorada do que a desagregação de esferoides para quantificação de números celulares. Por outro lado, Yepéz et al.55, conduzindo análises de XF em monocamadas de células de fibroblastos, descobriram que a normalização dos dados XF para o número celular introduziu maior dispersão de dados do que antes da normalização. O conteúdo de DNA nuclear é independente de estado ou fenótipo diferenciado e, portanto, é mais preciso para a normalização dos dados esferoides em ensaios XF do que o conteúdo proteico. O conteúdo de DNA também tem sido uma estratégia comprovada para a análise de outros conjuntos de dados ligados ao metabolismo56. No entanto, é importante notar que o conteúdo de DNA nuclear é quantificado de todas as células presentes dentro do esferoide; portanto, a normalização do conteúdo de DNA não é recomendada para conjuntos de dados XF em que os esferoides passam por tratamentos que podem resultar em perda significativa da viabilidade celular. Para tais conjuntos de dados, se possível, a normalização da viabilidade celular é preferida, ou os dados podem ser corrigidos na linha de base para respiração basal.
Usando a capacidade respiratória sobressalente como exemplo para a importância da normalização dos dados
A capacidade respiratória sobressal é uma medida da taxa de capacidade respiratória mitocondrial máxima menos taxa respiratória mitocondrial basal (Figura 6). No entanto, o problema com a notificação de dados desse tipo como taxa, ou seja, pmolO2/min/well dentro de determinados experimentos, é que os dados são vazios de normalização. Mesmo que normalize os dados eferóides para a densidade celular/conteúdo de DNA, isso muitas vezes exclui o parâmetro chave que precisa ser normalizado para densidade mitocondrial dentro das células. Dado que uma mudança na densidade mitocondrial levará a uma mudança proporcional na respiração basal e máxima, a capacidade de reposição também aumentará. Por exemplo, se o spheroid OCRbasal é 200 e o OCRmax é 400, a capacidade de reposição é relatada como 200; se o OCRbasal é 100 e OCRmax, então a capacidade de reposição também é de 100; no entanto, como porcentagem, ambos são 50% do máximo (ou 100% do basal). Portanto, a capacidade de reposição não é alterada entre esses dois exemplos, apesar das diferenças nas taxas de 200 e 100 quando calculadas como pmols O2/min/well. Valores internalmente normalizados são mais confiáveis e perspicazes para tornar os dados XF mais comparáveis entre estudos e projetos. Para fazer isso para a capacidade respiratória de reposição, optamos por apresentá-lo como uma porcentagem de respiração máxima em vez de uma taxa absoluta. Isso também pode ser apresentado como uma porcentagem da respiração basal. Este seria o caso se trabalhasse com células ou esferoides. No entanto, dado que a localização do esferoide na placa de microwell pode alterar o OCR absoluto, mas não as alterações relativas com inibidores ou desacopladores, é mais importante olhar para respostas internalmente normalizadas em esferoides como mudança de dobra ou percentuais.
Os modelos esferoides gerados aqui apresentam uma gama de tipos de células e arquitetura que não podem ser capturados em modelos clássicos 2D. Estes incluem arranjo heterogêneo, espacial de células em três dimensões, contatos de células celulares aprimoradas (por exemplo, formação de junções de lacunas e matrizes extracelulares), e gradientes bioquímicos em todo o diâmetro esferoide (por exemplo, gradientes de pH, acesso à difusão de oxigênio a nutrientes). O uso de fluxo extracelular para estudar biologia esferoide in vitro poderia permitir que alvos ideais para terapias medicamentosas fossem identificados através de observações de perturbação metabólica. Estes podem ser extrapolados de esferoides in vitro para tumores in vivo e identificar caminhos que podem atingir o metabolismo spheroid-tumor, por exemplo, a utilização de carboidratos durante o crescimento esferoide. As modalidades terapêuticas podem ser eficazes na segmentação de esferoides em fases iniciais de crescimento, mas se mostram menos eficazes nas fases posteriores do crescimento esferoide à medida que a complexidade da rede metabólica amadurece. Para concluir, o desenvolvimento de modelos de cultura celular 3D e tecnologias de análise sofisticadas em pesquisa biológica continuará sendo um campo dinâmico e em rápida mudança com potencial insuperável. A análise de fluxo extracelular de esferoides de cultura celular in vitro poderia ser empregada como um método de pesquisa de ponta para avançar os resultados da pesquisa que poderiam ser extrapolados para entender melhor a biologia relevante humana, reduzir melhor o uso de modelos animais em pesquisas e melhorar a pesquisa centrada no paciente.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Os autores não têm conflitos de interesse para declarar.
Acknowledgments
N.J.C foi apoiado por um Prêmio BBSRC MIBTP CASE com a Sygnature Discovery Ltd (BB/M01116X/1, 1940003)
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
Pesquisa sobre câncer edição 180Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
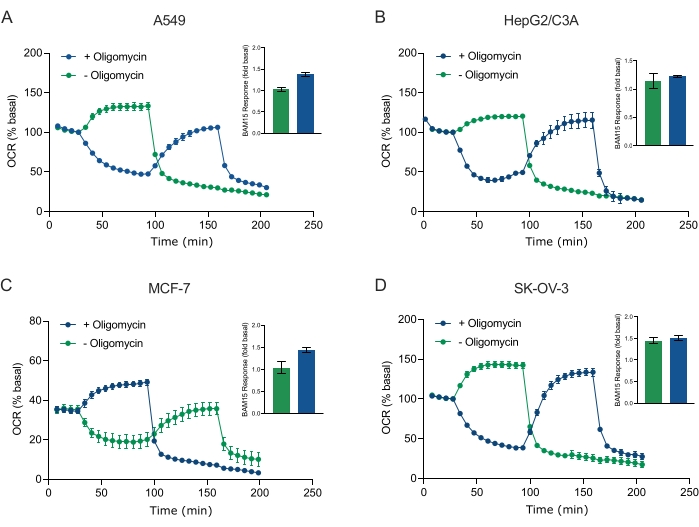
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
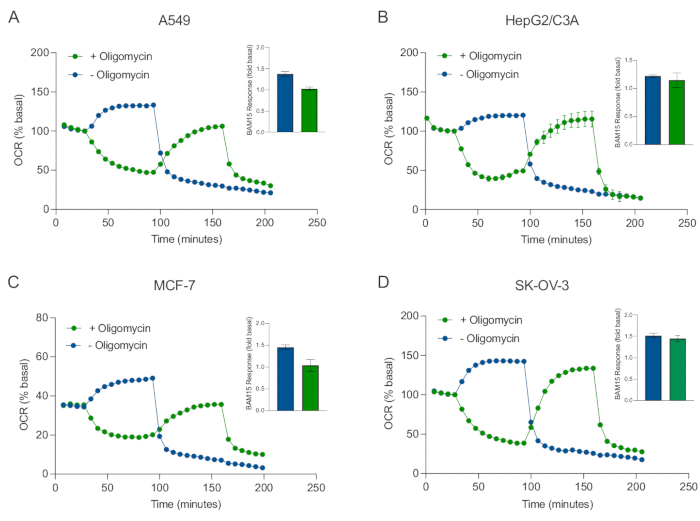
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
