

Summary
酵母の遺伝学的研究は、細胞のDNA代謝におけるヒト遺伝子の分子や細胞機能を調べるために用いることができる。メソッドは、人間の遺伝的特性に記載されている
Abstract
遺伝的経路でヒトDNA修復タンパク質の役割を理解することは多くの研究者に大きな課題です。哺乳動物系における遺伝学的研究は、定義されている変異遺伝子の細胞株、規制発現系、および適切な選択マーカーを含むすぐに利用できるツールの不足により制限されてきた。これらの困難を回避するために、下等真核生物のモデル遺伝システムは、機能的に保存されたDNA修復タンパク質や経路の研究のための魅力的な選択肢となっています。我々は、ウェルナー症候群ヘリカーゼ - ヌクレアーゼの不十分な定義遺伝子機能を研究するためのモデルの酵母のシステムを開発した(
Protocol
1。酵母株
野生型SGS1 TOP3を有する株(WT、W303 - 1A、遺伝子型、MAT ADE2 - 1 CANL - 100 HIS3 - 11、15 LEU2 - 3、112 TRPL - L URA3 - 1)[2]、SGS1変異体(W1292 - 3C 、遺伝子型のMAT SUP4 - O::URA3 SGS1 - 25 ADE2 - 1 CAN1 - 100 HIS3 - 11、15 LEU2 - 3、112 TRP1 - 1 URA3 - 1 rad5 - 535)とSGS1トップ3の変異体(W1058 - 11C、遺伝子型、MAT SUP4 - O::URA3 SGS1 - 25トップ3 - 2::HIS3 ADE2 - 1 CAN1 - 100 HIS3 - 11、15 LEU2 - 3、112 TRP1 - 1 URA3 - 1 rad5 - 535)[3]を特徴づけられていると親切に提供されていました博士ロドニーロススタイン(コロンビア大学)による。
2。プラスミドDNAの構造
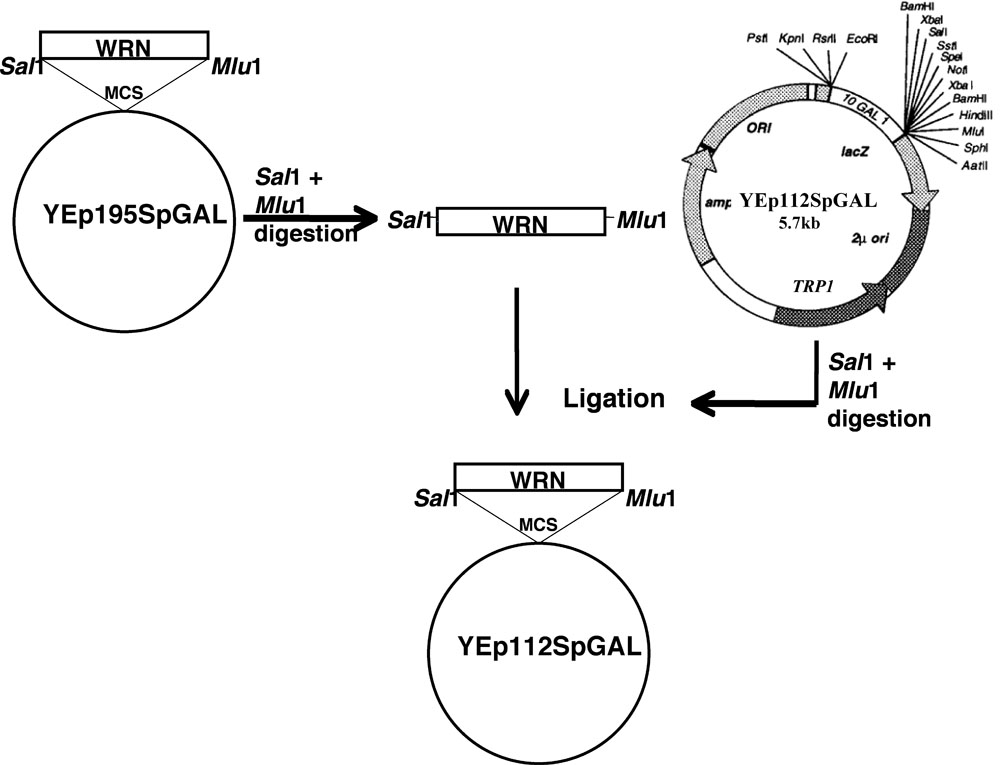
WRN遺伝子は、前述のスキームは、次のYEp112SpGALにクローニングした。
- プラスミドYEp195SpGAL - WRNのWRNの部位特異的変異(E84Aのエキソヌクレアーゼ死んだ、K577Mヘリカーゼ死んだ、R834C、そしてK1016A)[4]クイックチェンジII XL部位特異的突然変異誘発キット(Stratagene社製)から突然変異誘発プライマーと標準プロトコルを使用して構築されたLofstrandラボ(ゲイサーズバーグ、MD)によって。
- WRNとそれに関連する変異体をコードするDNA断片をゲルSalIとのMlu Iで二重消化し た後、それぞれのYEp195SpGAL - WRNのコンストラクトから精製した
- ゲル精製断片を、図1に示すように、GAL -誘導性プロモーターの制御下YEp112SpGAL WRNやWRNの変異体を構築するTRP1選択マーカーを含む2μmのマルチコピープラスミド、ベクターYEp112SpGALのSal IでのMlu I部位にクローニングし、 。
- このようにして得られたコンストラクトはYEp112SpGAL - WRN、YEp112SpGAL - WRN E84A、YEp112SpGAL - WRN K577M、YEp112SpGAL - WRN K1016A、およびYEp112SpGAL - WRN R834Cとして命名された。
3。変換
酵母の培養物は、標準プロトコルとの変換を用いて増殖させたGietz ら [5]により酢酸リチウムベースのプロトコルを用いて行った。簡単に言えば、次の手順を行った。
- 上記菌株の培養液を5mlのYPD(酵母エキス - ペプトン - デキストロース)培地で一晩増殖させた。文化を、50ml YPD培地でreinoculatedされ、1.0のOD 600まで増殖させた- 1.2に到達した。
- 培養は20 mlの水で洗浄し、回収し、100mMの酢酸リチウム(LiAc)1mlに再懸濁した。細胞を30 ° Cで10分間インキュベートした。
- 細胞ペレットは、10の変換を行うのに十分である500μlの最終容量には、100 mMのLiAc中に再懸濁した。細胞懸濁液の50μlを分注し、細胞ペレットを遠心分離により回収した。
- 250μlの50%ポリエチレングリコール(PEG)、1 M LiAc、2 mg / mlのssDNAを(サケ精子DNA)、プラスミドDNAを5 mlの(100 ngの25μlの36μL:細胞をペレットは、次の順序で追加されました1000ng/mlの)、および50μlの水。細胞ペレットは、1分間攪拌した。
- 次に、細胞を30℃でインキュベートした42℃のヒートショック℃で20分間に続いて30分間。
- 細胞を採取し、1mlの水に再懸濁した。プラスミドYEp112SpGALが選択可能なマーカーとしてTRPを含んでいるので、構造を保有するすべての変異株はtrp堪能になる。従って、形質転換体は、SC(合成完全な)グルコース培地トリプトファンを欠くと植民地が表示されるまで3-4日間、30℃でプレートをインキュベートにサスペンションをプレーティングによって選択した。
4。 WRNの低成長の表現型の回復の遺伝学的解析はSGS1トップ3のひずみを変形させた。
1。ストリーク分析:
- WRN(W303 - 1A)親の野生型の成長への発現、SGS1、またはSGS1トップ3の菌株の効果を調べるために、YEp112SpGALまたはYEp112SpGAL - WRNと対応する株は、2%グルコース(いずれかを含むSCマイナスのTrpプレート上にストリークしたGLU)または2%ガラクトース(Gal)。対照として、SGS1トップ3 / YEp112SpGAL - SGS1が含まれていた。プレートを2日間30℃でインキュベートした。
- 低WRN蛋白質発現のレベルでSGS1トップ3の菌株の増殖速度に影響を及ぼすWRNの能力を決定するために、SGS1トップ3の菌株からGALの様々な濃度を含むYEp112SpGAL、YEp112SpGAL - WRNで形質転換、またはYEp112SpGAL - SGS1は SCマイナスTrpプレート上に画線した2パーセントという高い0.005%と低い。その後、プレートを2日間、30℃でインキュベートした。
- SGS1トップ3のバックグラウンドの低成長の表現型を復元するWRNの機能要件を評価するために、YEp112SpGAL - WRNで形質転換SGS1トップ3(E84A、K577M、R834C、そしてK1016Aは)STREいた指示された濃度でGluまたはGALのいずれかを含むSCマイナスのTrpプレートにaked。コントロールについては、YEp112SpGAL、YEp112SpGAL - WRNまたはYEp112Sp GAL - SGS1で形質転換SGS1トップ3の菌株が含まれていた。その後、プレートを2日間、30℃でインキュベートした。
2。液体培養の解析:
- 液体培養で形質転換したSGS1トップ3の菌株の成長を評価するために、酵母の細胞を30℃で一晩SC RAFマイナストリプトファンで増殖させた。培養物は、SC RAFマイナスのTrpにreinoculated、30℃で初期対数増殖期(〜0.5のOD 600)まで増殖させた。培養物を指示された時間間隔で吸光度を測定することによって続いていた2%のギャルと成長を含むSC培地でOD 0.05と reinoculatedれました。
5。ヒドロキシまたは酵母株を形質転換したWRNのmethylmethane -スルホン酸感受性の遺伝学的解析。
- SGS1またはSGS1トップ3の菌株のMMSとHU感受性に関するWRNの発現の効果を判断するには、YEp112SpGALまたはYEp112SpGAL - WRNで形質転換株を、30℃SC RAFマイナストリプトファンで増殖させた℃の
- 培養物は、SC RAFマイナスのTrpにreinoculated、30℃で初期対数増殖期(〜0.6から0.8のOD 600)まで増殖させた。
- これらの株の10倍連続希釈をトリプトファンを欠くSC培地を用いて調製した。指数関数的に成長している文化から、5 × 10 6細胞を採取し、10,000倍にシリアルまで希釈した。各希釈の4つのマイクロリットルのHUやMMSの指示された濃度が含まれているSCギャルマイナスTrpプレート上にスポットした。プレートを30℃でインキュベートした。
- コントロールとして、野生型親株(W303 - 1A)はYEp112SpGALまたはYEp112SpGAL - WRN、またはSGS1トップ3 / YEp112SpGAL - SGS1で形質転換上記のように処理した。
6。 WRNの細胞周期分布はSGS1トップ3のひずみを変形させた。
- 勉強するWRNの細胞周期分布はSGS1トップ3の菌株、YEp112SpGALで形質転換SGS1トップ3の文化を変えて、YEp112SpGAL - WRN、またはYEp112SpGAL - SGS1、およびベクトル変換野生型親株(W303 - 1A)は、SC RAFで増殖させた30℃マイナスTRP℃で一晩のためのC。
- 培養はOD 0.05でreinoculatedと早期ログの段階(0.5 -0.6のOD 600)ゴマ増殖させた。WRNの発現を最終濃度2%にGALを追加することによって誘導した。培養物を30℃に成長させた℃で6時間インキュベートする。
- 培養物をDAPI(4'、6 - ジアミジノ-2 - フェニル)染色のために処理した。
- 培養物は、室温(RT)で10分間5800 × gで遠心分離によって回収した。
- 細胞は、1Xリン酸緩衝生理食塩水で一回洗浄した(PBS)、その後室温で20分間70%エタノールで固定した。
- 固定された細胞を1 X PBSで2回洗浄し、最後に1X PBSで再懸濁した。
- 細胞をスライド上に配置され、DAPI(1μg/ mlの)(ベクトル、米国)で培地をマウントVectashieldにマウントされた。と複合微分干渉コントラスト(DIC)、細胞は、Axiovert 200 M顕微鏡(100倍レンズツァイス)で調べた。蛍光画像はAxioVisionの、バージョン3.0のプログラム(ツァイス)を用いて分析した。
7。ウエスタンブロット分析。
1。 WRNのタンパク質発現や形質転換SGS1またはSGS1トップ3の菌株におけるWRNバリアントを決定するために、溶解物を、以下の方法とimmunblottingで分析したタンパク質によって調製した。
- 変換されたSGS1またはSGS1トップ3の菌株は、℃で一晩、30でSC RAFマイナストリプトファンで増殖させた。
- 培養は30℃reinoculatedとインキュベートした℃の初期対数増殖期(0.5のOD 600から 0.8)になるまで、次に最終濃度2%にGALを追加することによって誘導される。培養物を30℃に成長させた℃で6時間インキュベートする。
- 細胞(3 ml)を遠心分離により回収し、PBSで洗浄した。細胞ペレットは、アルカリ溶解バッファーに再懸濁し[50mMのNaOHを、pHが10.5、2mMのEDTA、1mMのフェニルメチルスルホニルフルオリド(PMSF)、2%SDS、10%グリセロール、5%2 - メルカプトエタノールおよびプロテアーゼ阻害剤(ロシュダイアグノスティックス)] 、遠心分離によって清澄化5分間、煮沸、および1M HClで中和した。
- 細胞溶解液の等量からのタンパク質は、8から16パーセントのポリアクリルアミドSDSゲルで分離された。
2。ガラクトースの濃度を変化の影響下にあるWRNタンパク質の発現レベルを決定するために、培養物を、以下のように処理されました。
- 変換されたSGS1トップ3の株は一晩30℃でSC RAFマイナストリプトファンで増殖させた。
- し、誘導されたGAを追加することで-この培養物を° C初期対数増殖期(0.8 0.5のOD 600)まで、30℃reinoculatedとインキュベートした最終濃度2%までのl。培養物を30℃に成長させた℃で6時間インキュベートする。
- 細胞を遠心分離により回収し、緩衝液に再懸濁した(50mMのトリス- Cl、pH7.5の、1%デオキシコール酸ナトリウム、1%トリトンX - 100、0.1%ドデシル硫酸ナトリウム[SDS])を含むプロテアーゼ阻害剤(0.05%PMSF、0.05μgの/ロイペプチン)。
- ガラスビーズ(425〜600μm)を混合物に添加し、細胞をボルテックスにより破壊した。
- 得られたホモジネートを15000 × gで5分間遠心分離し、上清画分を回収した。
- ライセート画分中のタンパク質はBradford法により推定した。
- 別のライセートの分画からのタンパク質の等しい濃度は8から16パーセントのポリアクリルアミドSDSゲルで分離した。
3。 WRNやWRN変異体タンパク質の発現は、WRNの精製C末端フラグメントのエピトープに対して向けWRNのマウスモノクローナル抗体を用いたウェスタンブロットにより測定した[6](1:1000、スプリングバレー研究所)。
4。一次抗体のインキュベーションは、二次(西洋ワサビペルオキシダーゼ結合)抗体(1:5000)でインキュベートした。ブロットは、メーカーのプロトコルごとに、ECL Plusは西洋検出システムで処理した。
5。定量的なウェスタンブロット解析のために、His -タグ全長のWRNタンパク質がブロットの定量化により、標準的な線形曲線が生成されるようなゲルに含まれていた組換え精製の 濃度を増加させる。さらに、酵母ライセートサンプル中の細胞または20μgの相当額からの細胞抽出物は、WRNの濃度を推定するためにゲルにロードされました。
6。 ImageQuant分析は、ゲル上で実行され、標準的な線形曲線が生成されました。
7。さまざまなギャルの濃度でWRNタンパク質の濃度は、上記生成された標準的な線形曲線を用いて推定した。
8。代表的な結果
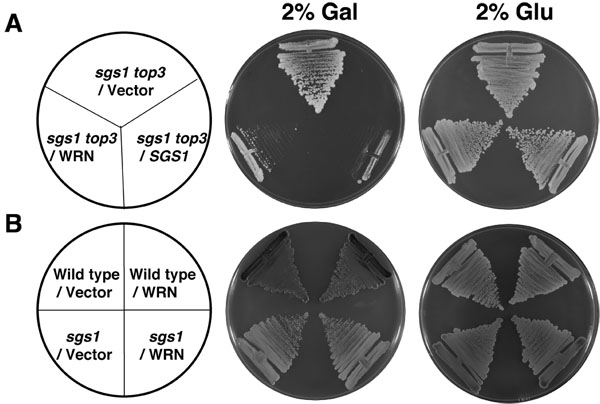
本研究で使用したすべての菌株は、トリプトファンの栄養要求です。 YEp112SpGALまたはYEp112SpGAL - WRNで形質転換従って、SGS1、SGS1トップ3と野生型W303 - 1A株はSCマイナスのTrp培地の存在下で増殖する能力に基づいて選択した。変換SGS1トップ3の変異細胞の増殖に対するWRNの発現の効果を調べるために、培養はWRN発現を誘導するために2%galでSCマイナスのTrp培地を含有するプレート上にストリークした。トップ3 decatenatesは、複製中にSGS1ヘリカーゼによって生成されたDNA分子[1、2]に絡み合っているため、トップ3の不在で、ねじり応力は、低成長と超組換えの結果を免れることができない。 SGS1の遺伝学的機能は、 トップ3の変異体の低成長の表現型を抑制することである。 WRNがトップ3との遺伝的相互作用にSGS1に代わることができれば、SGS1トップ3のトップ3の低成長の表現型の回復が期待される。
図の(a)に示すように、WRNはSGS1トップ3の歪ベクトルと比較して有意にゆっくりと成長がSGS1トップ3の菌株を形質転換した。YEp112SpGAL - SGS1プラスミドは、それが野生型デモンストレーション、ポジティブコントロール(図2A)として含まれていたで形質転換SGS1トップ3を変換SGS1トップ3の変異体で表現SGS1は遺伝的に成長の表現型を補完することができた。 2日間(図2A)のために示されるように変換さSGS1トップ3の菌株は、GALがない場合にも同様に増加した。 WRNの発現は、細胞増殖に対するWRNの効果はSGS1トップ3の変異体バックグラウンドで観察される特異的であることを示す、親の野生型(W3031A)またはSGS1株(図2B)の成長に与える影響はありません。遺伝学的研究は、WRNヘリカーゼがないエキソヌクレアーゼ活性がトップ3の成長の表現型(図3B)の修復に必要であることを実証WRNのバリアントを使用して実行。ヘリカーゼ活性を妨害するWRNで、天然に存在するミスセンス多型は、 トップ3の低成長の表現型を復元する能力を廃止。
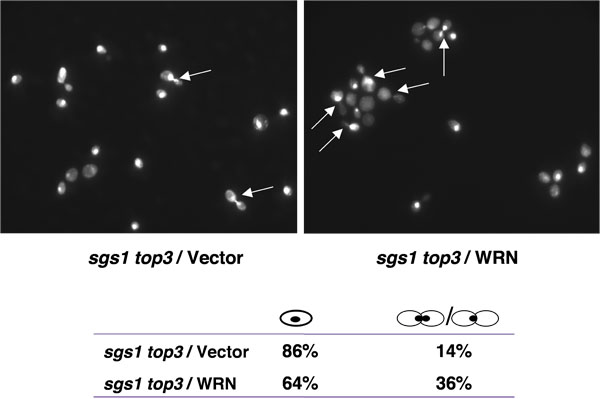
トップ3の変異株は細胞周期の後期S/G2の段階で遅れている[1]、それらの低成長を占めることが特徴。 トップ3のバックグラウンドでSGS1遺伝子の突然変異は、細胞周期のS/G2位相の遅れを抑制します。 WRNの発現はSGS1トップ3の細胞周期のS/G2位相の遅れを復元することができれば、分裂していない核をもつ大型出芽細胞の上昇の人口は、WRNの発現SGS1トップ3の変異体の細胞のために期待される。図4に示すように、大規模な出芽細胞の割合はS/G2ドの回復を示唆し、SGS1トップ3 /ベクトルよりSGS1トップ3 / WRNの方が高かった トップ3の特性を置く。
SGS1トップ3の二重変異体ではトップ3つの変異体[2]よりMMSまたはHUにはあまり敏感です。 SGS1トップ3のWRN影響を受ける細胞の成長以来、我々は次のメチルメタンスルホン酸(MMS、アルキル化剤)、ヒドロキシ(HU、複製の阻害剤)SGS1トップ3に匹敵する薬の両方に。SGS1トップ3 / WRN表示感度の感度に及ぼす影響を検討/ SGS1(図5)。 WRN亜種の遺伝学的解析は、WRNヘリカーゼ/ ATPアーゼではなく、WRNエキソヌクレアーゼ活性は、MMSとHUに感受性への影響のために必要とされることが明らかになった。

図1。 YEp112SpGALベクトルのクローニングWRN / WRNの亜種の模式図は。してくださいここをクリックして図1の拡大版のために。

図2。SGS1トップ3のWRNの発現は、 トップ3の低成長の表現型を復元します。 パネル YEp112SpGALまたはYEp112SpGAL WRNで形質転換、SGS1トップ3の菌株を2%Gluまたは2%のギャルのいずれかを含むSC - TRPプレート上にストリークした。 YEp112SpGAL SGS1で形質転換したコントロールSGS1トップ3株は、両方のプレート上にストリークしたとして。プレートを2日間30℃でインキュベートし、撮影した。 パネル YEp112SpGALまたはYEp112SpGAL WRNで形質転換されたB、野生型親株W303 - 1AまたはSGS1ひずみ 2パーセントGluまたは2%のgalを含むいずれかのSC - TRPプレート上にストリークした。プレートを4日間、30℃でインキュベートしてから撮影した。プレートの構成は、それぞれとパネルBパネルのようになった。してくださいここをクリックして図2の拡大バージョンを参照すること。

図3。WRN ATPアーゼ/ヘリカーゼではなく、エキソヌクレアーゼ活性は、SGS1トップ3のバックグラウンドでトップ3の低成長の表現型を復元するために必要です。SGS1 トップ3の株は(YEp195SpGAL WRN K577M)ATPアーゼ/ヘリカーゼ-死んだで形質転換、エキソヌクレアーゼデッド(YEp195SpGAL - WRN E84A)、RQC変異体(YEp195SpGAL - WRN K1016A)、または(YEp195SpGAL - WRN R834C)変異体ポリモーフィック型を、2%GLU(パネルC)または2%のギャル(パネルB)のいずれかを含むSC - Trpプレートにストリークした。プレートを2日間30℃でインキュベートし、撮影した。プレートの構成は、 パネルAのようにしてくださいでした、ここをクリックして図3の拡大バージョンを参照すること。

図4。WRNの発現は YEp112SpGAL、YEp112SpGAL WRN、またはYEp112SpGAL SGS1で形質転換SGS1トップ3の菌株の対数的に成長している文化が。SGS1トップ3の細胞内S/G2逮捕を誘発し、ベクトル変換野生型親株は2%ギャルの濃度で誘導された6時間培養物は、収穫などの材料および方法に記載のDAPI染色のために処理し、Axiovert 200 M顕微鏡(Zeiss社、100倍レンズ)を用いて観察されたされた。 YEp112SpGAL(左上)で形質転換SGS1トップ3のとYEp112SpGAL WRN(右上)とDAPI染色も示されています。矢印は、分割されていない核を持つ細胞を示す。 G1(単セル)とS/G2(出芽細胞)における細胞の分布は、下のパネルに示されています。してくださいここをクリックして図4の拡大バージョンを参照すること。

図5。 SGS1トップ3株のMMSとHU感度でWRNの発現の効果。YEp112SpGAL、YEp112SpGAL WRNで形質転換SGS1トップ3の菌株の対数的に成長する文化が、(YEp112SpGAL - WRN E84A) -死んだエキソヌクレアーゼは、ATPアーゼ/(YEp112SpGAL WRN K577M)ヘリカーゼ、死んだ、 RQC変異体は、(YEp112SpGAL - WRN K1016A)、変異体ポリモーフィック(YEp112SpGAL - WRN R834C)、YEp112SpGAL SGS1とベクトルは、親株はGluまたはgalおよびMMSやHUのいずれかを含むSC - Trpプレート上に十倍希釈で発見された野生型の変換指示された濃度で。プレートを30℃でインキュベートした後、2日間C(コントロールプレート)と4日(MMSとHUのプレート)と撮影。してください9/1639fig5.jpg">図5の拡大バージョンを見るにはここをクリックしてください。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
モデル系として、酵母を使用しての強みの一つは、酵母とヒトの間で保存されて定義されたDNAの複製と修復経路における変異体の可用性です。実験室株は、栄養要求性マーカーとの栄養要求性変異体とベクトルの容易に利用できるように特定の遺伝子を保有する形質転換体のさらなる選択が簡単で信頼性の高いです。これらのベクターを用いた遺伝子産物の発現は、誘導性プロモーターの制御(例えば、GALの誘導性プロモーター、等)の下に置くことで調節することができる。これらの利点を考慮し、我々は潜在的に人間と酵母の間で保存されて経路におけるウェルナー症候群における欠陥のある人間のWRN遺伝子の機能要件を研究するため、酵母のモデルシステムを開発した。説明するアプローチを使用して、我々は、WRNがトップ3に関連する表現型に影響を与える遺伝的経路で機能することができる最初の証拠を提供した。我々の観察では、WRNが機能的に細胞のDNA複製や組換えの間にヒトの細胞でTop3αと相互作用することを可能性のさらなる調査を促す。多分、BLMの不自由な状態で、WRNは部分的にトポイソメラーゼとの蛋白質のパートナーシップを通じてBLMのために代えることができる。
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
この作品は、老化NIH、国立研究所の学内研究プログラムによって完全にサポートされていました。我々は、プラスミドSGS1発現のための酵母菌株と博士ブラッドジョンソン(医学のペンシルバニア大学、フィラデルフィア、ペンシルベニア州)のために博士ロドニーロススタイン(コロンビア大学)を感謝。
References
- Aggarwal, M., Brosh, R. M. Jr WRN helicase defective in the premature aging disorder Werner Syndrome genetically interacts with Topoisomerase 3 and restores the top3 slow growth phenotype of sgs1 top3. Aging. 1, 219-233 (2009).
- Gangloff, S., McDonald, J. P., Bendixen, C., Arthur, L., Rothstein, R. The yeast type I topoisomerase Top3 interacts with Sgs1, a DNA helicase homolog: a potential eukaryotic reverse gyrase. Mol Cell Biol. 14, 8391-8398 (1994).
- Shor, E., Gangloff, S., Wagner, M., Weinstein, J., Price, G., Rothstein, R. Mutations in homologous recombination genes rescue top3 slow growth in Saccharomyces cerevisiae. Genetics. 162, 647-662 (2002).
- Sharma, S., Sommers, J. A., Brosh, R. M. Jr In vivo function of the conserved non-catalytic domain of Werner syndrome helicase in DNA replication. Hum Mol Genet. 13, 2247-2261 (2004).
- Gietz, R. D., Schiestl, R. H., Willems, A. R., Woods, R. A. Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast. 11, 355-360 (1995).
- von Kobbe, C., Thoma, N. H., Czyzewski, B. K., Pavletich, N. P., Bohr, V. A. Werner syndrome protein contains three structure specific DNA binding domains. J Biol Chem. 278, 52997-53006 (2003).

{kind=link}
{kind=link}
{kind=link}
{kind=link}