

Summary
De neuromusculaire junctie (NMJ) van
Abstract
De Drosophila larven neuromusculaire junctie (NMJ) is een uitstekend model voor de studie van synaptische structuur en functie. Drosophila staat bekend om het gemak van krachtige genetische manipulaties en het larvale zenuwstelsel heeft bewezen vooral nuttig bij het bestuderen van niet alleen de normale functie, maar ook storingen die gepaard gaan met een aantal neurologische aandoeningen (Lloyd en Taylor, 2010). Veel belangrijke synaptische moleculen gevonden in Drosophila zijn ook gevonden in zoogdieren en zoals de meeste CNS synapsen excitatoire in zoogdieren, de Drosophila NMJ is glutamaat en vertoont ook een activiteit-afhankelijke remodeling (Koh et al.., 2000). Bovendien kan Drosophila neuronen worden individueel geïdentificeerd omdat hun innervatie patronen stereotiepe en zich herhalende waardoor het mogelijk is om de vastgestelde synaptische terminals, zoals die tussen neuronen en het lichaam-muur spiervezels dat ze innerveren (Keshishian en Kim, 2004). Bestuderen Het bestaan van evolutionair geconserveerd synaps componenten, samen met het gemak van genetische en fysieke manipulatie maken het Drosophila model ideaal is voor het onderzoek naar de onderliggende mechanismen van synaptische functie (Budnik, 1996).
De actieve zones op synaptische terminals zijn van bijzonder belang, omdat deze zijn de sites van neurotransmitters. NC82 is een monoklonaal antilichaam dat het Drosophila-eiwit Bruchpilot (BRP), een CAST1/ERC familielid, dat is een belangrijk onderdeel van de actieve zone (Wagh et al., 2006). Herkent. BRP bleek direct de vorm van de actieve zone T-bar en is verantwoordelijk voor het effectief clustering Ca 2 +-kanalen onder de T-bar dichtheid (Fouquet et al.., 2009). Mutanten van Brp zijn verminderd Ca 2 + kanaal dichtheid, depressief opgeroepen blaasje release, en veranderde op korte termijn plasticiteit (Kittel et al., 2006).. Wijzigingen in actieve zones zijn waargenomen in Drosophila ziekte modellen. Bijvoorbeeld, immunofluorescentie met behulp van de NC82 antilichaam bleek dat de actieve zone dichtheid was afgenomen in modellen van amyotrofe laterale sclerose en Pitt Hopkins-syndroom (Ratnaparkhi et al., 2008;. Zweier et al., 2009.). Zo kan de evaluatie van actieve zones, of andere synaptische proteïnen, in Drosophila larven van de ziekte van een waardevolle eerste aanwijzing voor de aanwezigheid van een synaptische defect.
Voorbereiden hele-mount ontleed Drosophila larven voor immunofluorescentie analyse van de NMJ vereist enige vaardigheid, maar kan worden bereikt door de meeste wetenschappers met een beetje oefening. Gepresenteerd, is een methode die zorgt voor meerdere larven worden ontleed en immunostained in dezelfde dissectie gerecht, het beperken van het milieu de verschillen tussen elk genotype en zorgen voor voldoende dieren voor vertrouwen in de reproduceerbaarheid en statistische analyse.
Protocol
1. Voorbereiding voor immunofluorescentie:
- Voor het maken van een dissectie oppervlak, giet Sylgard 184 Siliconen Elastomeren Base in een kleine weefselkweek plaat. Zorg ervoor dat niet op de plaat te vullen volledig, zodat de dissectie gebied lager is dan de velg.
- Trim dissectie pinnen tot een lengte van ongeveer 3,5 mm voor meer gemak van manipulatie, en zorg ervoor dat ten minste 6-pins per larve hebben.
- U moet stompe tang waarmee u begrijpen de pinnen voor de dissectie.
2. Dissectie van Larven:
- Selecteer zwervende derde instar larven die kruipen rond de zijkanten van de flacon en zijn niet begonnen om zich te verpoppen.
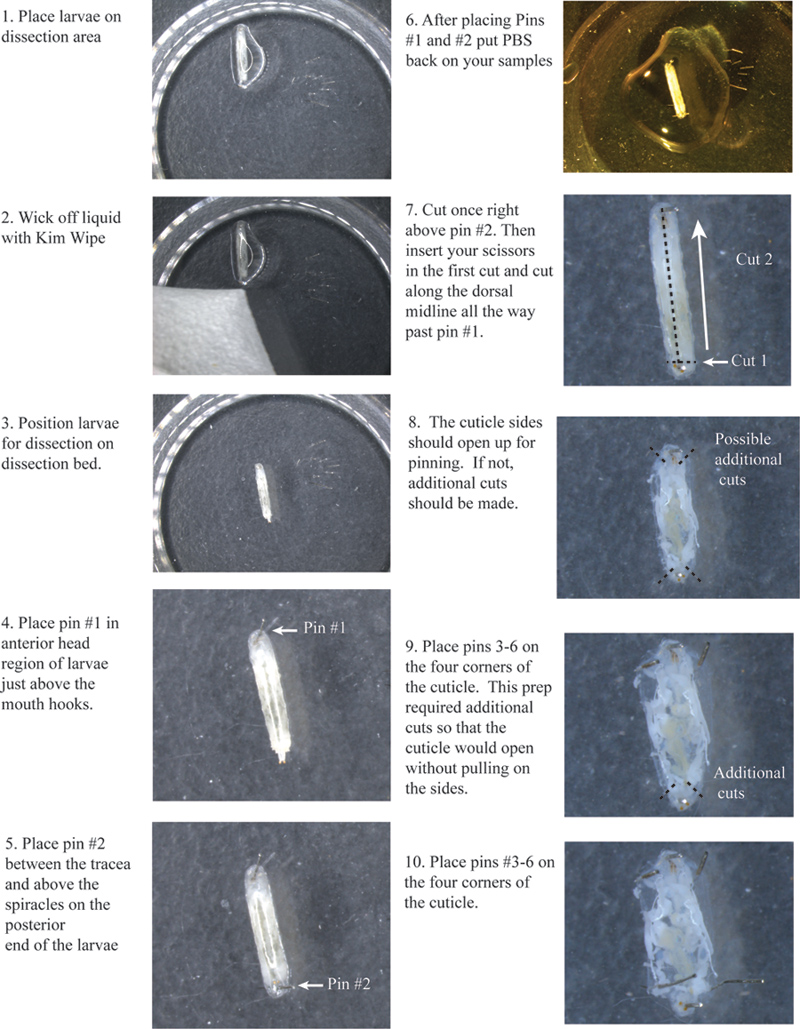
- Plaats een larve op de dissectie ondergrond en onder een stereomicroscoop. Wick overtollige vochtiger met behulp van een Kimwipe.
- Schik elke larve, zodat de dorsale zijde naar boven is gericht. Je zult in staat zijn om twee luchtpijp, witte lijnen te zien langs de dorsale zijde van de larven. Probeer het centrum van deze in het midden zoveel mogelijk want dit markeert de dorsale middellijn.
- Plaats pin # 1 net boven de mond haken (figuur 1).
- Plaats pin # 2 in de staart net boven de siphonen tussen de tracheeën.
- Voeg een kleine hoeveelheid van PBS om de larven te voorkomen dat het drogen van de cuticula.
- Herhaal de bovenstaande stappen met alle larven, zorg ervoor dat de genotypen ruimtelijk worden gescheiden door het plaatsen van een pin tussen elk genotype.
- Maak een kleine dwarse snede recht boven pin # 2 in zowel luchtpijpen (figuur 1).
- Steek uw schaar in de eerste snede en knip langs de dorsale middellijn tussen de luchtpijpen helemaal tot het verleden pin # 1.
- U moet andere kleine bezuinigen op de top en onderkant van elke kant van de nagelriem, zodat de linker-en rechterkant zal openen zodat u ze pinnen zonder trekken aan de nagelriem.
- Plaats pin # 's 3, 4, 5 en 6 op elke hoek.
- Bevestig het monster met 4% PFA voor 20-25 minuten bij kamertemperatuur
- Giet het PFA en spoelen met PBS 3-4 keer, decanteren per keer.
3. Verwijderen van Tissue:
- Begin met het verwijderen van weefsel aan het achterste einde, want normaal gesproken wilt u het gebied waar de hersenen zich bevindt te beschermen.
- Hoewel het gebruik van een steromicroscope en met de larven onder een daling van PBS, eerst pak het achterste einde van een van de luchtpijp en trek het uit die een deel van het vet lichaam te verwijderen. Herhaal dit voor de tweede luchtpijp.
- Pak een deel van de achterste vet lichaam of darm en trek ze uit de prep. Zodra het grootste deel van de achterste weefsel is verwijderd, richten zich op het verwijderen van de kleine weefsels rond de hersenen. Als u alleen geïnteresseerd bent in de spieren, of de morfologie van de NMJ u kunt verwijderen van de hersenen ook.
- Het verwijderen van de hersenen zorgt voor een betere visualisatie van Muscle 31, dat is een nuttige marker voor abdominale segment oriëntatie.
4. Immunofluorescentie voor Active Zones:
- Voeg blokkerend middel (5% normaal Geit serum in PBS) aan de ontleed preps in de dissectie schotel. Blok voor 1 uur bij kamertemperatuur. Als u wilt een oplossing alleen voor het blokkeren en antilichaam verdunningen, onder meer 0,1% TritonX-100 om uw blokkerend middel voor weefsel permeabilisatie.
- Breng de primaire antilichaam verdund in het blokkeren van reagens met 0,1% TritonX-100. De actieve zone antilichaam is bereikbaar via de Developmental Studies Hybridoma Bank (DSHB, http://dshb.biology.uiowa.edu/). Gebruik een referentie-antilichaam (bijv. mierikswortelperoxidase-Cy3 conjugaat) om de NMJ membraan en axonen label.
- Incubeer de primaire antilichamen bij 4 ° C gedurende de nacht. Zodat de droogte van de koelkast niet zal het antilichaam oplossing verdampen, maak een bevochtigingskamer door er wat water op de bodem van een gebruikte pipetpunt box. Bedek de doorzichtige deksel met aluminium folie om tl verminderen bleken. Plaats de dissectie dia in de vochtigheid kamer en zet de hele doos in de 4 ° C incubator 's nachts.
- De volgende ochtend, wassen met PBST (PBS met 0,2% Triton-X-100) 5-10 keer gedurende 5-10 minuten elk.
- Incubeer met het secundaire antilichaam (bijv. Invitrogen Alexa geit anti-muis-488) verdund in het blokkeren van reagens met 0,1% TritonX-100 bij kamertemperatuur gedurende 2-4 uur in de overdekte luchtvochtigheid kamer. Onder meer een nucleaire vlek (bijv. DAPI) in de secundaire, als je zou willen om de locatie van de kern in de spieren of de hersenen te identificeren.
- Wassen met PBST (PBS met 0,02% Triton-X-100) 5-10 keer gedurende 5-10 minuten elk.
5. Montage van de larvale monsters op Dia's:
- Bezinken en lont uit de laatste PBST wassen.
- Pipet ongeveer 1 ml 80% glycerol aan het vastgepind larvale huiden te dekken.
- Verwijder 5 van de 6 pinnen in alle larven.
- Wanneer u klaar bent om de eerste genotype op een dia de laatste pin te verwijderen voor alle larven van die genotype.
- Bereid een "wash" slide door twee druppels van uw montage medium op een schoon objectglas. Deze stap is belangrijk om de oplossing van water wassen de larven voor het monteren en duidelijker beeldvorming.
- Pak een larve van de staart met je botte tang, sleept u de larve rond in de glycerol en dan langs de droge randen van uw dissectie schaal, tot een meerderheid van de glycerol te verwijderen. Plaats de larven in de eerste montage medium te laten vallen.
- Herhaal stap 6 voor alle van de larven van hetzelfde genotype.
- Zodra alle larven in de eerste montage medium laten vallen, de eerste larve bij de staart pakken en haal deze door de druppel en dan langs de droge randen van de schotel naar een meerderheid van het medium te verwijderen. Plaats dan de larven in de tweede montage medium te laten vallen.
- Herhaal stap 8 voor alle larven met hetzelfde genotype.
- Bereid uw laatste dia door etikettering van het en het plaatsen van een kleine "T" vorm van montage medium op de dia.
- Net als in stap 5.8, pak de larve van de staart, de vacht te trekken door de tweede montage medium laten vallen en dan langs de droge zijden van de "Wash" slide. Na het grootste deel van het medium is losgeraakt, plaats de larvale vacht op de laatste dia van de montage medium.
- Herhaal dit voor alle larven pelzen.
- Zorg ervoor dat alle larvale huiden zijn gerangschikt naar wens ervoor te zorgen dat de cuticula naar beneden is en de blootgestelde spier naar boven is gericht.
- Plaats een dekglas op de dia om de ontleed larven te dekken, met het uitgangspunt rand in de bovenste lijn van de "T"
- Zet een klein gewicht (bijv. 9 volt batterij) op de omslag slip en zorg ervoor dat de montage medium volledig verspreidt onder het dekglas.
- Een vermindering van de tl-quenching, plaats de dia in een lade of donkere ruimte totdat de montage medium is uitgehard.
- Het schoonmaken van de montage medium dat de randen van de kap slip overgelopen. Schone overtollige droog montage medium op de cover slip of dia met 70% ethanol.
- Om een definitieve afdichting te creëren, afdichting met nagellak.
6. Representatieve resultaten:
Een voorbeeld glijbaan van ontleed en immunostained larven is weergegeven in figuur 2. Wanneer de larven dissecties, vlekken, en montage correct zijn uitgevoerd, kan de gebruiker de larven spier van belang (figuur 3) te identificeren in segmenten A2-A6. Hoang et al.. Geeft een gedetailleerde beschrijving van de spier positie als karakterisering van specifieke synaptische boutons (Hoang en Chiba, 2001). Het gebruik van een hoog vermogen het doel van de gebruiker kan zich richten op een enkele synaps en neem z-stack beeld. Een vertegenwoordiger maximale projectie beeld van een deel van de synaps 6 / 7 wordt getoond (figuur 4). Vervolgens met behulp van imaging-software, kan de actieve zones kwantitatief worden beoordeeld op kenmerken die specifiek zijn voor uw mutant fenotype. Voorbeelden van functies die kunnen worden veranderd is op een bepaalde mutant fenotype omvatten het totale aantal, fluorescentie-intensiteit, en de afstand tussen de actieve zones.
Mogelijke valkuilen:
Immunofluorescentie evaluatie van de larvale neuromusculaire overgang is een waardevol systeem dat waardevol inzicht kunnen geven in de synaptische biologie, al zijn er potentiële valkuilen dat het nut van dit systeem kunnen beperken. Bijvoorbeeld, vaak is het wenselijk om het imago van de kleuringspatroon van twee of meer primaire antilichamen in hetzelfde monster. De keuze van de juiste primaire antilichamen, passende fluoroforen, en passende negatieve controles is essentieel voor succes.
Als imaging twee antigenen in uw experiment, er zeker van zijn dat de primaire antilichamen zijn van verschillende soorten, zodat elk eiwit van belang kan uniek worden geïdentificeerd met een fluorescent gelabelde secundaire antilichaam. Een aanbevolen manier om de achtergrond te verminderen is een blokkerend middel te gebruiken met de normale serum van de soorten waarvan de secundaire antilichamen zijn afgeleid (bv. normale geit serum te gebruiken als een blokkerend middel bij het gebruik van geit anti-konijn of anti-muis secundaire antilichamen).
Na de planning van de primaire en secundaire antilichamen die u wilt gebruiken in je experiment, moet u ook rekening houden met controles in your vlekken sets om ervoor te zorgen dat het fluorescerende signaal dat je ziet is echt en niet alleen achtergrond. Indien mogelijk, onder meer negatieve controle monsters die niet bevatten het eiwit van belang. Dit is eenvoudig te doen in studies van exogene transgene overexpressie door het opnemen van een wild type dier, maar in sommige gevallen kan worden bereikt door middel van een homozygoot mutant van uw eiwit van belang. Een ander essentieel negatieve controle, nuttig voor het meten van de achtergrond niveaus van je fluorescerende signaal, is het secundaire antilichaam te gebruiken zonder de primaire. Wanneer de resulterende beeldvorming preps, is het belangrijk om te begrijpen dat alle van de fluorescerende signaal dat je ziet misschien niet echt zijn, en kan achtergrond van your secundaire antilichamen of van endogene weefsel achtergrond. Het kan nodig zijn om uw afbeeldingen te normaliseren op de achtergrond fluorescentie overwegen wanneer het testen voor de veranderingen tussen fluorescentie-intensiteit.
Keuze van de fluorofoor moet ook worden gemaakt met zorg. Behandeling van de excitatie-en emissie-spectra voor elke fluorofoor en de mogelijkheid om unieke sporen elk signaal is essentieel voor kanaal bloeden te vermijden door. Als er een onzekerheid, een eenvoudige manier om te bepalen of de omvang van bloeden door middel is om met een enkele fluorofoor-gelabeld antilichaam immunostain op een moment en het beeld controleren met ieder kanaal dat u gaat gebruiken in uw uiteindelijke experiment. Dit moet laten zien als bloeden door zich voordoet met uw microscoop filter in te stellen.

Figuur 1. Stroomdiagram van de cuticula te snijden en het plaatsen van pinnen 1-6. Klik hier om een grotere afbeelding te bekijken

Figuur 2. Plaatsing van larven op de laatste dia. Zorg ervoor dat de larven preps zijn spier-kant bloot op en neer cuticula kant. Laat op zijn minst een larvale lichaam breedte in elk monster.

Figuur 3. Bij beeldvorming, zoek de spieren en segmenten van belang. Voorkomende neuromusculaire verbindingen gekenmerkt innerveren de spieren 6 / 7, 13 spieren, spier-12, of spier 4. Aan de andere kant van de ventrale middellijn, is er een spiegelbeeld spier structuur van de getoonde hemi-segment. Gebruik Muscle 31 als een marker om te bepalen welke segmenten je bent beeldvorming. Het imago van de dezelfde synapsen en segmenten voor de rest van je monsters, het verzamelen van veel synaptische beelden per genotype die kunnen worden gebruikt voor de kwantificering.

Figuur 4. Een vertegenwoordiger maximaal projectie beeld van een deel van de NMJ innerveren spier 6 / 7 wordt getoond. NC82 (Bruchpilot) kleuring is in het groen weergegeven. HRP pre-synaptische vlek wordt getoond in het rood. Let op de actieve zone puncta die verder kunnen worden gekwantificeerd door imaging software.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Voor neuronen, de synaptische terminal gebied is van cruciaal belang, en is de brug voor een goede communicatie tussen de post-en pre-synaptische cellen. Een krachtige manier om de gezondheid van het neuron te onderzoeken bij de ziekte van modellen is aan eiwitten van de synaptische terminal te analyseren door immunofluorescentie. De immunofluorescentie methode hier gepresenteerde Hiermee kan de onderzoeker een groot aantal larven tegelijk onderzoeken, terwijl het beperken van de ecologische verschillen tussen de groepen. Het centrale zenuwstelsel van Drosophila derde instar larven heeft vele voordelen, waaronder glutamaat synapsen, in kaart gebracht en herhaalde synaptische terminals, toegankelijkheid, reproduceerbaarheid en de kracht van genetische manipulatie. Specifiek in Drosophila neuronale ziekte-modellen, heeft de concentratie van de werkzame zone eiwit Bruchpilot is gewijzigd. Vanwege het grote aantal larven tegelijkertijd getest, de actieve zone analyse met behulp van de gepresenteerde methode kan de onderzoeker te detecteren subtiele verschillen tussen de groepen die een fundamenteel gebrek in de gezondheid van de neuron zou kunnen geven.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Geen belangenconflicten verklaard.
Acknowledgments
Wij danken dr. Nael Alami en Dr Nam Chul Kim voor hun nuttige opmerkingen over dit manuscript.
Materials
| Name | Company | Catalog Number | Comments |
| Sylgard 184 Silicone Elastomer Base | Dow Corning | 68037-59-2 | After mixing allow for bubbles to rise slowly out by putting on slow rotator or allowing to sit for 30 minutes or more. |
| Stainless Steel Minutien PIns | Fine Science Tools | 26002-10 | Trim to approx. 3-4mm in length with regular scissors |
| Laminectomy Forceps (Blunt- Used for grasping pins) | Fine Science Tools | 11223-20 | Use as blunt forceps for grasping pins |
| Dissection Forceps | World Precision Instruments, Inc. | 501985 | |
| SuperFine Vannas Scissors, 8cm long | World Precision Instruments, Inc. | 501778 | |
| Mouse anti-Brp antibody | DSHB | NC82 | Use 1:50 dilution |
| Cy3 Affinipure Goat Anti-Horseradish Peroxidase | Jackson ImmunoResearch | 123-165-021 | Use at 1:200 dilution |
| Alexa Fluor 488 Goat anti-Mouse IgG | Invitrogen | A11001 | Use approx. 1:200 dilution |
References
- Budnik, V. Synapse maturation and structural plasticity at Drosophila neuromuscular junctions. Curr Opin Neurobiol. 6, 858-867 (1996).
- Fouquet, W., Owald, D., Wichmann, C., Mertel, S., Depner, H., Dyba, M., Hallermann, S., Kittel, R. J., Eimer, S., Sigrist, S. J. Maturation of active zone assembly by Drosophila Bruchpilot. J Cell Biol. 186, 129-145 (2009).
- Hoang, B., Chiba, A. Single-Cell Analysis of Drosophila Larval Neuromuscular Synapses. Developmental Biology. 229, 55-70 (2001).
- Keshishian, H., Kim, Y. -S. Orchestrating development and function: retrograde BMP signaling in the Drosophila nervous system. Trends in Neurosciences. 27, 143-147 (2004).
- Kittel, R. J., Wichmann, C., Rasse, T. M., Fouquet, W., Schmidt, M., Schmid, A., Wagh, D. A., Pawlu, C., Kellner, R. R., Willig, K. I., Hell, S. W., Buchner, E., Heckmann, M., Sigrist, S. J. Bruchpilot Promotes Active Zone Assembly, Ca2+ Channel Clustering, and Vesicle Release. Science. 312, 1051-1054 (2006).
- Koh, Y. H., Gramates, L. S., Budnik, V. Drosophila larval neuromuscular junction: Molecular components and mechanisms underlying synaptic plasticity. Microscopy Research and Technique. 49, 14-25 (2000).
- Lloyd, T. E., Taylor, J. P. Flightless flies: Drosophila models of neuromuscular disease. Ann N Y Acad Sci. 1184, e1-e20 (2010).
- Ratnaparkhi, A., Lawless, G. M., Schweizer, F. E., Golshani, P., Jackson, G. R. A Drosophila model of ALS: human ALS-associated mutation in VAP33A suggests a dominant negative mechanism. PLoS One. 3, e2334-e2334 (2008).
- Wagh, D. A., Rasse, T. M., Asan, E., Hofbauer, A., Schwenkert, I., Dürrbeck, H., Buchner, S., Dabauvalle, M. -C., Schmidt, M., Qin, G., Wichmann, C., Kittel, R., Sigrist, S. J., Buchner, E. Bruchpilot, a Protein with Homology to ELKS/CAST, Is Required for Structural Integrity and Function of Synaptic Active Zones in Drosophila. Neuron. 49, 833-844 (2006).
- Zweier, C., Jong, E. K. de, Zweier, M., Orrico, A., Ousager, L. B., Collins, A. L., Bijlsma, E. K., Oortveld, M. A., Ekici, A. B., Reis, A., Schenck, A., Rauch, A. CNTNAP2 and NRXN1 are mutated in autosomal-recessive Pitt-Hopkins-like mental retardation and determine the level of a common synaptic protein in Drosophila. Am J Hum Genet. 85, 655-666 (2009).

{kind=link}