

Summary
神经肌肉接头(NMJ)
Abstract
果蝇幼虫神经肌肉接头(NMJ)突触的结构和功能研究的一个很好的模型。果蝇是缓解强大的遗传操作和幼虫的神经系统已被证明不仅在学习的正常功能特别有用,但也扰动伴随一些神经系统疾病(Lloyd和泰勒,2010)。在果蝇中发现的许多关键的突触分子还发现,在哺乳动物中,最喜欢在哺乳动物中枢神经系统兴奋性突触,果蝇NMJ是谷氨酸和演示活动依赖的重塑(Koh等 , 2000)。此外, 果蝇的神经细胞可以单独确定,因为他们的神经支配模式都是千篇一律和重复,使我们能够研究确定的突触终端,如运动神经元和体壁肌纤维之间,他们支配(Keshishian和Kim ,2004)。进化上保守的突触组件以及与遗传和体质操纵缓解的存在, 使得果蝇调查的内在机制,突触功能的(Budnik 1996)的理想模型。
在突触终端的活动区域特别感兴趣,因为这些神经递质释放的网站。 NC82是一种单克隆抗体,能够识别的果蝇蛋白Bruchpilot(BRP),CAST1/ERC家庭成员的活动区(Wagh 等人 ,2006年),是一个重要组成部分。 BRP是直接形成活动区的T -酒吧,并负责有效聚类的Ca 2 +通道下方的T -酒吧密度(富凯等人,2009年) 。 BRP突变体减少的Ca 2 +通道密度,郁闷诱发囊泡释放,并改变短期的可塑性( 基特尔等,2006) 。活跃区的改建已发现在果蝇的疾病模型。例如,使用的NC82抗体的免疫荧光显示,活跃的区域密度下降在肌萎缩性脊髓侧索硬化症和皮特霍普金斯综合征(Ratnaparkhi等,2008; Zweier等,2009)。模型。因此,活跃的区域,或其他疾病的果蝇幼虫模型,突触蛋白的评价可能会提供一个宝贵的初步线索,一个突触缺陷的存在。
准备整个安装解剖果蝇幼虫的免疫分析NMJ需要一些技巧,但可以稍加练习,大多数科学家所完成。是提供多个幼虫在同一解剖盘解剖和免疫染色方法,限制各基因型之间的环境差异,并提供足够的动物重现性和统计分析的信心。
Protocol
1。对免疫的筹备工作:
- 要创建一个解剖面,倒入一个小的组织培养板Sylgard 184硅橡胶基地。确保不来填充板完全使清扫面积低于轮辋。
- 修剪清扫引脚易于操纵增加了约3.5毫米长度,并确保至少有6%的幼虫引脚。
- 您将需要钝钳,让你掌握清扫引脚。
2。解剖的幼虫:
- 选择流浪三龄幼虫蠕动着小瓶双方还没有开始化蛹。
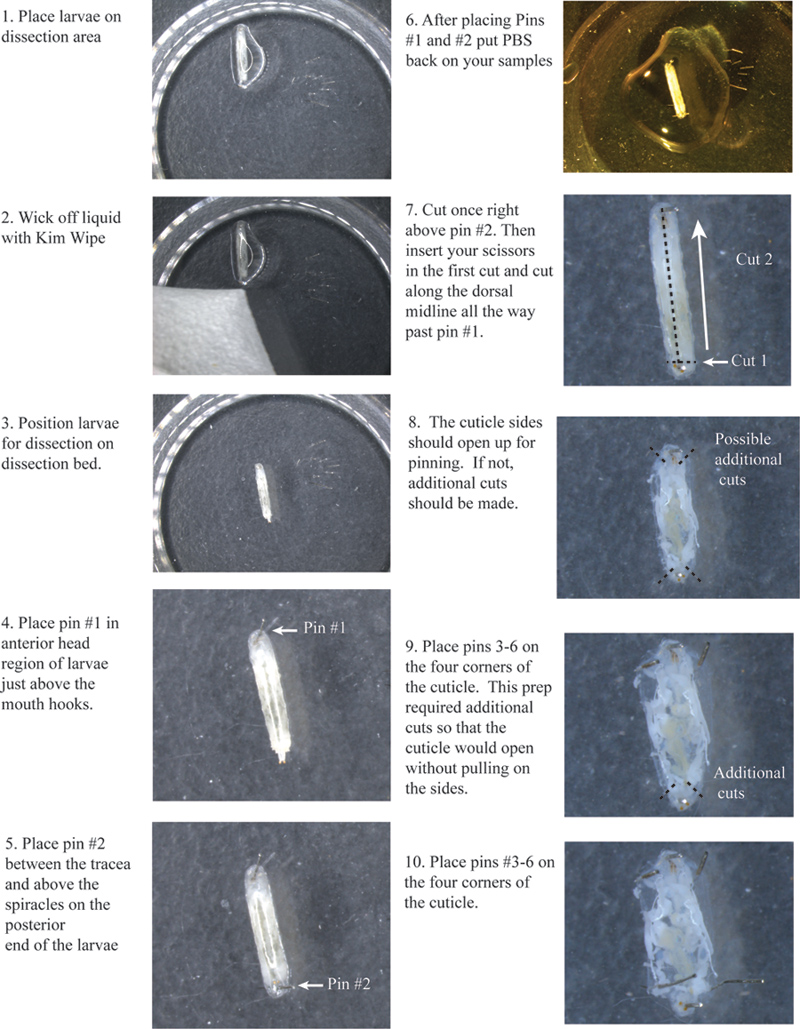
- 广场上的立体显微镜下解剖面的幼虫。威克走多余的潮湿使用Kimwipe。
- 安排每个幼虫,使背侧朝上。您将能够看到两个气管,白线沿背侧幼虫。中心在中间尽可能多的尝试,因为这标志着背中线。
- 将针,只是上面的嘴钩#1(图1)。
- 广场针在正上方气孔之间的气管的尾巴#2。
- 添加少量的PBS幼虫,以防止干燥的角质层。
- 与所有的幼虫,重复上述步骤,确保放置各基因型之间的引脚的空间分隔的基因型。
- 跨越气管(图1)#2针以上的小横削权。
- 插入你的剪刀剪切和沿背中线之间的气管切开了过去针一路#1。
- 您将需要在每一侧的角质层非常的顶部和底部的小伤口,使左,右两侧会开放让您针没有角质层拉下来。
- 放置接脚#“第3,4,5,6,并在每个角落。
- 修正了4%的煤灰样品在室温为20-25分钟
- 倒出的煤灰和3-4次,用PBS漂洗,每次调迁。
3。去除组织:
- 开始删除,因为通常你要保护的区域位于大脑后部组织。
- 而使用steromicroscope下的PBS中的幼虫,先抢后气管结束,并将其拉出,可能会删除一些与脂肪体。重复第二气管。
- 抓住后脂肪体或肠道和拉出来的预习。一旦后组织的大部分已被删除,着眼于消除大脑周围的小组织。如果你只在肌肉,或形态的NMJ感兴趣,可以去除大脑。
- 去除大脑可以更好地肌肉31日,这是一个有用的标记腹段取向的可视化。
4。为活动区域的免疫:
- 添加阻断剂(5%正常山羊血清的PBS)在解剖盘的解剖PREPS。在室温下1小时座。如果您想封锁和抗体稀释液,不仅使一个解决方案,包括0.1%TritonX - 100你阻止组织通透性的代理。
- 应用的主要抗体阻断0.1%TritonX - 100的试剂稀释。活动区抗体是通过发育研究杂交瘤细胞银行(DSHB,http://dshb.biology.uiowa.edu/)达到。使用引用抗体(如辣根过氧化物酶Cy3标记物)标签NMJ膜和轴突。
- 在4 ° C过夜培养的主要抗体。因此,冰箱干燥不会蒸发的抗体解决方案,创建湿度室,把一些水在使用枪头框的底部。半透明的盖子盖用铝箔,以减少荧光漂白。放置在湿度室的清扫幻灯片,并在4℃孵化器把整个中过夜。
- 第二天早上,洗脸用PBST(0.2%TRITON - X - 100的PBS)的5-10倍,每5-10分钟。
- 孵育的二级抗体(如Invitrogen公司Alexa的山羊抗鼠- 488)在0.1%TritonX - 100阻断试剂在室温覆盖湿度试验箱2-4小时稀释。包括在二次核染色(如DAPI)的,如果你想确定在肌肉或脑细胞核的位置。
- 用PBST(0.02%TRITON - X - 100的PBS),洗净的5-10倍,每5-10分钟。
5。在幻灯片上安装的幼虫样品:
- 倒出和灯芯最后PBST洗涤。
- 移液器约1毫升80%甘油,以牵制的幼虫毛皮。
- 删除5 6引脚在所有的幼虫。
- 当你准备把幻灯片上的第一基因型,删除最后一个引脚,所有的幼虫,genotyPE。
- 把一个干净的幻灯片上两滴您的安装介质的准备“洗”的幻灯片。这一步很重要的,洗掉的幼虫水解决方案,安装和更清晰的成像。
- 抓住您的钝钳尾一幼虫,幼虫在甘油拖动周围,然后沿夹层菜干边,除去大多数的甘油。放置在第一个安装介质下降的幼虫。
- 同一基因型的幼虫,重复步骤6。
- 一旦所有的幼虫都在第一次安装介质下降,抢到第一幼虫的尾巴,转危为安下降,沿干边的菜,然后删除了大多数中等。然后将安装在第二个中期下降的幼虫。
- 所有具有相同基因型的幼虫重复步骤8。
- 准备好你最后的幻灯片,标签,并在幻灯片上放置一个小的“T”型安装介质的形状。
- 在步骤5.8中,抓住的尾巴的幼虫;转危为安第二安装介质下降,然后沿“洗”幻灯片干燥双方的皮毛。大多数介质脱落后,幼虫皮上安装介质中的最后一张幻灯片。
- 重复所有的幼虫毛皮。
- 确保所有的幼虫毛皮安排需要确保角质层方下来,裸露的肌肉朝上。
- 将幻灯片上的盖玻片覆盖解剖的幼虫,在“T”上线边缘
- 盖玻片放在一个小,重量轻(如9伏电池),确保安装介质传播的掩护下滑完全。
- 为了减少可能的荧光猝灭,放置在抽屉或黑暗的空间幻灯片,直到安装介质已治愈。
- 清理安装介质溢出盖玻片一柄。盖玻片或用70%乙醇的幻灯片上,擦去多余的干安装介质。
- 要创建一个最终的密封,指甲油密封。
6。代表性的成果:
解剖和免疫染色的幼虫例如幻灯片,如图2所示。当幼虫解剖,污渍,并安装得当,用户可以识别A2 - A6段肌肉(图3)幼虫的利益。晃等提供了肌肉位置的详细描述,以及具体的突触boutons(晃和千叶,2001年)的表征。使用高功率的目标用户可以专注于一个单一的突触和采取的Z - Stack的形象。突触6 / 7的一部分的一个代表最高的投影图像(图4)所示。然后用图像处理软件,活跃的区域,可定量评估,尤其是您的突变表型特征。可能在一个特定的突变表型的改变的功能的例子包括的总数量,flourescence强度,活动区域之间和间距。
潜在的隐患:
幼虫神经肌肉交界处的免疫评价是一个有价值的的系统,可以到突触生物学提供了宝贵的见解,虽然有潜在的缺陷,可以限制这一制度的效用。例如,它常常是可取的图像在同一样品的两个或两个以上的主要抗体染色模式。选择适当的主要抗体,适当的荧光基团,和相应的阴性对照的是取得成功的关键。
如果成像两个抗原在实验中,可以肯定,使每一个感兴趣的蛋白质,可以唯一一个荧光标记的第二抗体确定不同物种的主要抗体。一个推荐的方式,以减少背景是使用阻断剂含有从中派生的二级抗体(如用正常山羊血清作为阻断剂时,使用羊抗兔或抗鼠二抗)的物种的正常血清。
规划的小学和中学的抗体在实验中,您将使用后,你还必须考虑包括在您的染色的控制设置,以确保您看到的荧光信号是真实的,不只是背景。如果可能的话,包括阴性对照样品,不包含感兴趣的蛋白质。中的外源基因的过度表达的研究,这是很容易做到,包括野生型动物,但在某些情况下可以使用一个你感兴趣的蛋白质的纯合子突变的完成。另一个重要的负面控制,用于测量您的荧光信号的背景水平,是不包括主使用的二级抗体。成像时产生的PREPS,重要的是理解的荧光信号,你看到的所有可能不是真正的的,可能从哟背景UR二次抗体或内源性组织背景。考虑背景荧光,荧光强度之间的变化化验时,您可能需要将您的图像的正常化。
荧光的选择也必须小心。每个荧光基团和能力的唯一检测每个信号的激发和发射光谱的代价是必要的,以避免渠道流失,通过的。如果有任何的不确定性,一个简单的方法来确定是否出血的程度,通过免疫染色与荧光标记的单一抗体的时间和检查,每个通道的图像,你会在你最后的实验中使用。这应该发现,如果通过流血与您的显微镜过滤器设置发生。

图1。角质层的切割和安置的针脚1-6。流程图点击这里查看大图

图2。安置的最后一张幻灯片上的幼虫。确保,幼虫PREPS暴露肌肉侧和角质层一面向下。在每个样本之间至少留出幼虫体宽度。

图3。成像时,找到利益的肌肉和段。常见的神经肌肉接头的特点支配肌肉的6 / 7,肌肉13 12,肌肉或肌肉4。在腹中线的对面,有一个镜像显示半段的肌肉结构。作为一个标记,以确定哪个段是成像肌肉31。相同的突触图片和段其余样品,收集许多突触%的基因型,可用于定量使用的图像。

图4。NMJ支配肌肉的6 / 7的一部分的一个有代表性的最大投影图像显示。 NC82(Bruchpilot)染色显示为绿色。 HRP的前突触染色显示为红色。注意活动区域punctae图像处理软件,可以进一步量化。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
神经元突触终端领域是至关重要的,是适当的沟通后,突触前细胞之间的桥梁。调查疾病模型的神经元的健康,是一种强有力的方式分析,免疫荧光蛋白质的突触终端。这里介绍的免疫荧光法,使研究者检查许多幼虫的同时,限制群体之间的环境差异。 果蝇三龄幼虫的中央神经系统有许多优点,包括谷氨酸的突触,突触末端,交通方便,重现性和遗传操作的权力映射和重复。特别在果蝇神经元疾病模型,活动区蛋白Bruchpilot的水平已经改变。由于大量的幼虫同时测定,活跃区分析使用的方法,使研究人员检测组之间的细微差别可以反映在健康的神经元的基本缺陷。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
没有利益冲突的声明。
Acknowledgments
我们感谢他们对这个手稿的有益的意见Nael阿拉米博士和博士南澈金。
Materials
| Name | Company | Catalog Number | Comments |
| Sylgard 184 Silicone Elastomer Base | Dow Corning | 68037-59-2 | After mixing allow for bubbles to rise slowly out by putting on slow rotator or allowing to sit for 30 minutes or more. |
| Stainless Steel Minutien PIns | Fine Science Tools | 26002-10 | Trim to approx. 3-4mm in length with regular scissors |
| Laminectomy Forceps (Blunt- Used for grasping pins) | Fine Science Tools | 11223-20 | Use as blunt forceps for grasping pins |
| Dissection Forceps | World Precision Instruments, Inc. | 501985 | |
| SuperFine Vannas Scissors, 8cm long | World Precision Instruments, Inc. | 501778 | |
| Mouse anti-Brp antibody | DSHB | NC82 | Use 1:50 dilution |
| Cy3 Affinipure Goat Anti-Horseradish Peroxidase | Jackson ImmunoResearch | 123-165-021 | Use at 1:200 dilution |
| Alexa Fluor 488 Goat anti-Mouse IgG | Invitrogen | A11001 | Use approx. 1:200 dilution |
References
- Budnik, V. Synapse maturation and structural plasticity at Drosophila neuromuscular junctions. Curr Opin Neurobiol. 6, 858-867 (1996).
- Fouquet, W., Owald, D., Wichmann, C., Mertel, S., Depner, H., Dyba, M., Hallermann, S., Kittel, R. J., Eimer, S., Sigrist, S. J. Maturation of active zone assembly by Drosophila Bruchpilot. J Cell Biol. 186, 129-145 (2009).
- Hoang, B., Chiba, A. Single-Cell Analysis of Drosophila Larval Neuromuscular Synapses. Developmental Biology. 229, 55-70 (2001).
- Keshishian, H., Kim, Y. -S. Orchestrating development and function: retrograde BMP signaling in the Drosophila nervous system. Trends in Neurosciences. 27, 143-147 (2004).
- Kittel, R. J., Wichmann, C., Rasse, T. M., Fouquet, W., Schmidt, M., Schmid, A., Wagh, D. A., Pawlu, C., Kellner, R. R., Willig, K. I., Hell, S. W., Buchner, E., Heckmann, M., Sigrist, S. J. Bruchpilot Promotes Active Zone Assembly, Ca2+ Channel Clustering, and Vesicle Release. Science. 312, 1051-1054 (2006).
- Koh, Y. H., Gramates, L. S., Budnik, V. Drosophila larval neuromuscular junction: Molecular components and mechanisms underlying synaptic plasticity. Microscopy Research and Technique. 49, 14-25 (2000).
- Lloyd, T. E., Taylor, J. P. Flightless flies: Drosophila models of neuromuscular disease. Ann N Y Acad Sci. 1184, e1-e20 (2010).
- Ratnaparkhi, A., Lawless, G. M., Schweizer, F. E., Golshani, P., Jackson, G. R. A Drosophila model of ALS: human ALS-associated mutation in VAP33A suggests a dominant negative mechanism. PLoS One. 3, e2334-e2334 (2008).
- Wagh, D. A., Rasse, T. M., Asan, E., Hofbauer, A., Schwenkert, I., Dürrbeck, H., Buchner, S., Dabauvalle, M. -C., Schmidt, M., Qin, G., Wichmann, C., Kittel, R., Sigrist, S. J., Buchner, E. Bruchpilot, a Protein with Homology to ELKS/CAST, Is Required for Structural Integrity and Function of Synaptic Active Zones in Drosophila. Neuron. 49, 833-844 (2006).
- Zweier, C., Jong, E. K. de, Zweier, M., Orrico, A., Ousager, L. B., Collins, A. L., Bijlsma, E. K., Oortveld, M. A., Ekici, A. B., Reis, A., Schenck, A., Rauch, A. CNTNAP2 and NRXN1 are mutated in autosomal-recessive Pitt-Hopkins-like mental retardation and determine the level of a common synaptic protein in Drosophila. Am J Hum Genet. 85, 655-666 (2009).

{kind=link}