

Summary
Dopamin utbyte farmakoterapi med användning av L-DOPA är den vanligast använda symptomatisk behandling av Parkinsons sjukdom, men åtföljs av biverkningar innefattande ofrivilliga onormala rörelser, benämnd dyskinesi
Abstract
MALDI imaging masspektrometri (IMS) är en kraftfull metod som underlättar den rumsliga analysen av molekyler i biologiska vävnadsprover 2 (Fig. 1). En 12 ^ m tunn vävnad sektionen är täckt med ett MALDI-matris, vilken underlättar desorption och jonisering av intakta peptider och proteiner som kan detekteras med en massanalysator, typiskt med användning av en MALDI-TOF / TOF-masspektrometer. Generellt hundratals toppar kan bedömas på ett enda råtta hjärnvävnad avsnitt. I motsats till vanliga avbildningstekniker, kräver detta tillvägagångssätt inte förkunskap av molekylerna av intresse och möjliggör oövervakad och omfattande analys av flera molekyler samtidigt som en hög molekylär specificitet och sensitivitet 2. Här beskriver vi ett MALDI IMS-baserad metod för att belysa regionspecifika fördelningsprofilerna av neuropeptider i råtthjäman av en djurmodell av Parkinsons sjukdom (PD).
PD är en vanlig neurodegenerativ sjukdom med en prevalens på 1% för personer över 65 ålder 3,4. Den vanligaste symtomatisk behandling bygger på dopamin utbyte med L-DOPA 5. Men detta åtföljs av allvarliga biverkningar inklusive ofrivilliga onormala rörelser, kallas L-DOPA-inducerad dyskinesi (LID) 1,3,6. En av de mest framträdande molekylära förändringar i locket är en uppreglering av opioiden föregångaren prodynorfin mRNA 7. De dynorfin peptider modulera neurotransmissionen i områden i hjärnan som främst är involverade i rörelsekontroll 7,8. Har dock hittills exakt opioida peptider som härstammar från bearbetning av neuropeptid föregångaren inte karakteriserats. Därför använde vi MALDI IMS i en djurmodell av experimentell Parkinsons sjukdom och L-DOPA-inducerad dyskinesi.
MALDI avbildning masspektrometri visade sig vara särskilt fördelaktig med avseende på neuropeptid characterizaning, eftersom vanliga antikroppsbaserade metoder riktar kända peptidsekvenser och tidigare observerats posttranslationella modifieringar. Däremot MALDI IMS kan reda ut nya produkter peptidbearbetning och därmed visa på nya molekylära mekanismer för neuropeptid modulering av neuronala överföring. Medan den absoluta mängden av neuropeptider inte kan bestämmas genom MALDI IMS, den relativa förekomsten av peptid joner kan avgränsas från masspektra, vilket ger insikter om hur du ändrar nivåer i hälsa och sjukdom. I exemplen som presenteras här, var toppintensiteterna av dynorfin B, alfa-neoendorphin och substans P visade sig vara betydligt större i dorsolaterala, men inte dorsomediala, striatum hos djur med svår dyskinesi innefattar ansikts-, bål och orolingual muskler (Fig. 5). Dessutom visade MALDI IMS ett samband mellan dyskinesi svårighetsgrad och nivåerna av des-tyrosin alfa-neoendorphin, som representerar en tidigare okänd mekanism för funktionell inactition av dynorfiner i striatum som avlägsnandet av N-terminala tyrosinen reducerar dynorfin s opioid-receptor-bindande förmåga 9. Detta är den första studien om neuropeptid karakterisering i locket med MALDI IMS och resultaten belysa de möjligheter som tekniken för tillämpning inom alla områden av biomedicinsk forskning.
Protocol
Protokollet justeras för att statistisk analys av MALDI IMS data från flera avsnitt råtta hjärna, typiskt 20-30 sektioner och består av fem olika steg som omfattar vävnader förberedelser, matris ansökan MALDI-TOF MS-analys, data utvärdering och neuropeptid identifiering. Förfarandena beskrivs och beskrivs mer detaljerat nedan:
1. Vävnadspreparat
Detta förfarande omfattar insamling av respektive vävnadsprover samt mjukpapper sektionering för IMS analys. En särskild mål protein och peptid analys är att undvika proteolytisk nedbrytning. Därför är det viktigt att arbeta snabbt och flitig under vävnad dissektion.
- Offra råttor (typiskt 250-300 g) genom halshuggning, avlägsna råtthjärna inom högst post-mortem tid <30s och frysning av pulveriserad torris innan de överförs till -80 ° C frys. Snabbare frysning med flytande kväveöka risken för microtears i hjärnvävnad, som negativt påverkar matrisens kristallisation och därigenom reducera MS kvalitet (fig. 2D). Hela hjärnor kan lagras i flera år innan snittning utan förlust av MS signalkvalitet.
- Skära frusen vävnad på ett kryostatmikrotom till 12 | im snitt och tining-mount vävnadssektioner på ledande MALDI glasskivor (indiumtennoxid belagda objektglas, Bruker Daltonics) eller MALDI-målet (fig. 2A-C).
- Torra delar under 15 minuter under vakuum och glider lagra vid -80 ° C tills vidare användning. Vävnadssnitt bör analyseras inom kortast möjliga tid efter snittning, även om den förvaras vid -80 ° C. Vi finner att MS signalkvaliteten påtagligt kommer att minskas efter ett år i lager. För att minska oxidation av proteiner och peptider, kan luften i lagringsbehållaren ersättas med en inert gas (t.ex. argon eller kväve).
2. Matrisen ansökan
Matrisen ansökansteg har en betydande inverkan på spektrumet kvalitet och kräver optimering av multipla parametrar beror på den typ av vävnad samt till analyten av intresse. Dessa faktorer inkluderar kemiska parametrar såsom typ av matris, matris koncentration, pH, vävnad tvätt och organiska modifierare samt instrumentala inställningar inklusive deponering volym, lateral upplösning och antal vittnesmål 10 (Fig. 2D). För storskaliga experiment, är det av stor vikt att minska variationen, exempelvis genom applicering av matrisen till alla sektioner inom en dag och av samma operatör. Även om det finns många strategier för att tillämpa matrislösning såsom genom sublimering eller genom sprutning, den automatiserade avsättning av nät av små droppar matris, ca 100-150 pikoliter i storlek, har använts med framgång för analys av små proteiner och neuropeptider i olika vävnader inklusive hjärnsektioner 9, 10,11, 12, 13.
- Upptining sektionerna i en exsickator under 1 hvår.
- Se till att experimentet förblindad av en annan person än operatören. Re-märka alla prover.
- Tvätta sektioner 1x i 70% etanol (EtOH, vid rumstemperatur, RT) under 10 sek och två gånger i 95% EtOH (RT) under 10 sek. För stora experiment utföra tvättningen för alla glas glider tillsammans med användning av en kyvett för att minimera variationen.
- Torka de sektioner i en exsickator under 10 min.
- Utvärdera vävnadssnitt i mikroskop och kontrollera vävnader distorsion, microtears och små sprickor som försämrar MALDI MS kvalitet (Fig. 2D).
- Framställ färsk matris bestående av 50 mg / ml DHB i 50% metanol, 10% 150 mM ammoniumacetat (AMAC) och 0,3% trifluorättiksyra (TFA) i vatten.
- Matrisen ansökan utförs genom diskreta droppavsättningsanordning i ett rektangulärt mönster med användning av en skrivare kemisk bläckstråleskrivare (CHIP, Shimadzu). Det första steget är att optimera de experimentella parametrar matrix ansökan om neuropeptid analys including antalet droppar per genomgång, antal passeringar. Detta experiment utförs genom att applicera flera matrix matriser med olika applikationsparametrar på samma vävnadssnitt, samtidigt se till att varje uppsättning täcker liknande hjärnan regioner som corpus callosum, cortex och striatum. Samma experiment måste utföras varje gång parametrar ändras, inklusive olika hjärnstrukturer, olika matriser riktade specifika analyter, och om olika matris lösningsmedel behövs för utvinning av specifika analyter.
- Skanna innehavaren glasskiva med vävnaden sektionen och justera hållaren. Definiera din array för matrix program på vävnadssnittet och ange den rumsliga plats upplösningen, dvs att upptäcka avstånd. Applicera matris med den optimerade protokollet om kemiska bläckstråleskrivare. För detta experiment använde vi en optimerad protokoll för peptid avbildning med följande utskrift parametrar: 10 droppar (100 PL / drop), passerar 10 ansökan och en plats att upptäcka distance av 300 pm.
- Scan slutliga matrisen såg sektioner och spara bilden för registrering innan MALDI datainsamling (steg 3,4).
- Lagra de sektioner tills vidare användning i en exsickator under vakuum.
3. MALDI MS datainsamling och bearbetning
MS-analys av neuropeptider utförs på ett MALDI time of flight instrument (Ultraflex II, Bruker Daltonics, Tyskland) som arbetar i reflektorn läge, med hjälp av programvara assisterad datainsamling från varenda matris plats 14. Därför exakt rumslig undervisning är absolut nödvändigt. Det är viktigt att MALDI optimering, förvärv och särskilt mål experimenten registrering utförs av samma operatör som helst skall förblindas de experimentella grupperna. I ett storskaligt experiment med flera glasskivor, kan MALDI experimenten utföras av en operatör medan en annan person använder skrivaren kemiska bläckstråleskrivare.
- Ladda glasskivor i masspektrometern.
- Kontrollera kalibreringen av MALDI förvärvsmetoden med ett lågmolekylärt standard kalibrering mix (Bruker Daltonics).
- Optimera förvärv parametrar.
- För att optimera MS-signal och för att undvika abiadering matrisen från angränsande matris avlagringar, bör storleken av lasern och den optimala inriktningen på vävnaden bestämmas.
- Lasern energin sätts att säkerställa maximal MS kvalitet från så många matris insättningar som möjligt utan att höja baslinjen, vilket minskar topp upplösning eller mättar detektorn.
- Bedöm det maximala antalet skott per matris plats tills bara brus upptäcks, ofta 1000-2000 skott. Uppskatta antal skott som ska ackumulerade och antalet bilder som förvärvats före lasern position inom en plats bör ändras. För att prova varje matris plats jämnt ackumuleras vi 600 bilder i 25 skott steg för totalt antal 24 stegen med en RANDom rörelsemönster, från varje matrixdeposition.
- Registrera genomsökning av alla fläckiga avsnitt till motoriska koordinater för MALDI scenen med hjälp av FlexImaging (v.2.0) 10 och utföra datainsamling i batch-läge genom att AutoXexuteBatchRunner.exe programvara.
- Process varje enskild spektra med hjälp av baseline subtraktion (konvex skrovet V3), utjämning och extern kalibrering (tillval), följt av export som en ascii-fil (*. Dat, *. Txt eller *. Csv-format) 15.
4. Data utvärdering
Slutlig analys av data består av uppgifter efterbehandling och data minskning med fokus bara på topp information, följt av statistisk analys.
- Som ett första steg, var MALDI IMS avsnitten utvärderas för overnormalization effekter. Detta kan lätt uppnås genom användning av data visualiseringsverktyg såsom FlexImaging (Bruker Daltonics) eller BioMap (Novartis). Som ett första steg de totala jon bilderna evaluated innan total jonström (TIC) normalisering, följt av manuell inspektion av enstaka bilder jon distribution av olika framträdande peptid toppar. Leta efter karakteristiska fördelningar toppintensitet och om de är relaterade till vävnad funktioner (skador), observation kvalitet eller normalisering effekter (Fig. 3).
- Avgränsa områden av intresse (t.ex. striatum) enligt histologiska funktioner och exportera motsvarande spektra i ASCII-format. Företrädesvis kan normalisering av spektra till den totala jonströmmen (TIC) utföras vid detta steg.
- Importera ASCII-filer i en databas hantering av programvara såsom Ursprung (V.8.1, Originlab), MATLAB (MathWorks, Natick, MA, USA) eller R 16. Toppdetektering kan utföras med maximal söktjänster som ingår i programmet, till exempel "peak analys" i Ursprung eller "mspeaks" i Matlab. Exportera peaklists från hela spektra som en enda tabbavgränsad textfil.
- För att bestämma bin gränser för upptäckt peptidtoppar är binning analys med hjälp av lämpliga programverktyg (t.ex. pBIN 17) eller in-house skrivit manus för MATLAB eller R. Här den enda textfil som innehåller all toppen plockade data laddas in i programmet och parametrarna för maximal gräns bestämma anges såsom hur ofta en toppen bör vara närvarande i spektra för att vara relevant för experimentet. Till exempel innehåller experimentet 2 grupper av djur, 5 djur i varje grupp, och 100 spektra insamlas från varje djur och regionen av intresse. Antar en topp är potentiellt intressant om det är närvarande i åtminstone huvuddelen av djuren i en grupp (3/5) och i åtminstone hälften av spektra för dessa djur (3x50 = 150 spektra), kommer detta att ge en total andel på 15% för 150 positiva spektra av totalt 1000 (2x5x100) spektra. Med hjälp av pBIN verktyget ger detta steg en enda binrange fil som innehåller alla bin bredder bestäms från de förvärvade data. För att kontrollera att bin gränserär lämpliga, är det lätt att visualisera facken i Origin tillsammans med de ursprungliga spektra spåren.
- Peak-området integration kan minska variansen som är viktig för statistisk analys. Vi använder en in-house skrivet manus för R för att beräkna arean under kurvan mellan topp gränser som bestäms i steg 4. Integrerade toppareor importeras till MS Excel (v.2007) och statistisk analys genom icke-parametrisk oparade tester med SAM verktyget utförs 18.
5. Peptididentifiering
Sekvens verifiering av de observerade peptid identiteter är nödvändig för att ingå biologiska relevans. Den mest exakta metoden omfattar sant top-down beslut direkt från vävnad med hjälp av peptidfragmentering med hjälp av tandem masspektrometri (MS / MS), även om höga peptidkoncentrationer krävs för denna typ av analys 12,13. För låga rikliga peptider eller flera peptider med closEM / z-värden (± 0,5%), är on-vävnadsanalys försämras och bort vävnad analys med användning av en peptidomic strategi utnyttjas som innefattar extraktion, separation och MS baserad identifiering av endogena neuropeptider. För experimentet presenteras här, var det centrala fokus på opioid peptid upptäckt, vilket är en särskild utmaning eftersom dessa peptider är ganska låg rikligt jämfört med andra neuropeptider i spektra. Dessutom har dessa peptider är ganska polära vilket gör dem relativt hydrofil och svåra att bibehålla med gemensam peptid extraktion och separationstekniker .. Därför har vi tillämpat en tidigare rapporterats protokoll för vävnader utvinning och opioid peptid prefraktionering i kombination med standard LC-MS/MS baserad peptid identifiering 9,19.
- Samla koronala delar av målstrukturer av intresse (nucleus accumbens, NAc, caudatus putamen, CPU). Montera frysta råtthjärna i en kryostatmikrotom och ta bort omgivande material från hjärnan (motsvarandetex, septum, corpus callosum) med en skalpell. Samla sektioner (30 m, n = 50) av dissekerades NAC och CPU och tina montera NAC och CPU delar av avsnitten om olika glasskivor.
- Extrahera peptider off vävnad genom att tillsätta 100 | il 5% ACN/0.1% TFA, inkubera under två minuter och uppsamlas i Eppendorf låg proteinbindning rör. Upprepa detta steg två gånger.
- Utföra peptid förfraktionering medelst kraftig katjonbyteskromatografi med användning stegvis (n = 4) eluering vid förhöjd jonstyrka 19. Torkar prover under vakuum med användning av en speedvac-koncentrator.
- Analysera peptidfraktioner genom nanoflow omvänd C18-fas-vätskekromatografi (1100, Agilent, Santa Clara, CA) förbunden med elektrospray tandemmasspektrometri (LC-MS/MS). MS Experimenten utfördes på en hybrid linjärt iontrap / Fouriertransform jon cyklotron resonans (FTICR) instrument (LTQ FT 7T, Thermo Scientific, Waltham, MA,) peptid fulscan spektra (m / z 150-2000) Förvärvades med FTICR analysatorn med hög massa upplösning (100000), följt av efterföljande fragmentering av de 5 mest intensiva peptid toppar i iontrap genom kollision dissociation (CID) 9.
- Peptidsekvensen identifiering utförs genom databasen matchning och kan kompletteras genom de-novo-sekvensering analys. För databassökning, kommersiellt tillgängliga sökmotorer (Mascot, XTandem eller Protein Prospector) används 20. Sökningarna är vanligtvis utförs mot databaser som innehåller sekvenser av kända eller förväntade neuropeptider och sekvenser av neuropeptid prekursorproteinerna 21.
6. Representativa resultat
MALDI avbildning masspektrometri av striatala vävnadssnitt som framställts enligt det protokoll som beskrivs här resulterat i upptäckten av mer än 1000 toppar som motsvarar ca 300 monoisotopiska molekylslag (genomsnitt spektra visasi Fig.. 1). Data visualisering för framstående molekyljonen toppar uppnåddes med hjälp av Flex Imaging programvara och visade karaktäristiska distributioner toppintensitet som är väl i linje med anatomiska funktioner (Fig. 3). Ett ytterligare särdrag hos MALDI IMS är dess relativa god reproducerbarhet. I detta experiment var den totala variationskoefficient för toppintensiteter för alla detekterade molekyler 30%, men många toppar uppvisade mycket låg variation och hög reproducerbarhet i behandlingsgrupper (fig. 4). De relativa toppintensiteten uppgifter av fyra olika regioner av intresse, inklusive dorsolaterala och dorsomediala del av både skadade och oskadade striata utsattes för statistisk analys. För att justera för multipla jämförelser samtidigt, var den statistiska analysen utförs med hjälp av icke-parametrisk test med hjälp av SAM verktyget 18. De mest framträdande förändringarna hittades i dorsolaterala delen av dopamin-denerverad, Parkinsons striatum. Här si gnificant förändringar mellan de olika behandlingsgrupperna observerades under två dynorfin peptider, dynorfin B och alfa-neoendorphin (fig. 5). I detalj, har en relativ ökning av både dynorfin toppintensiteter med 50-60% observerades i höga dyskinetiska djur jämfört med låga dyskinetiska djur och kontroller lesion (p <0,05, F (2, 15) = 12,8 DynB, F = 5,7 aNeo; FIG. 5).

. Figur 1 Genomsnitt MS spårar erhölls från två nära relaterade regioner av striatum; caudate putamen (CPu) och nucleus accumbens (NAc). De två regionerna visa olika MS profiler med några molekyler unikt uttryck i en region, eller vid olika maximala intensitetsnivåer (insert, m / z 2028). Rymdfördelningen mönster av varje topp kan visualiseras med hjälp av specialiserad bildprogram (nedre panelen).
2.jpg "/>
Figur 2 (A) Hjärnan är monterad på en kryostat patronen med hjälp av ett omslutande medium (OTC; pil). Dock försiktighet iakttas så att OTC inte förorenar området av hjärnan för att vara sektionerad eftersom OTC orsaken jon undertryckande av peptider. (B, C) Tunna sektioner (≈ 12 | im tjocklek) är tö-monterades på MALDI kompatibel glasskivor och torkades under några sekunder för att undvika frysning skador som ses i C. (D) Microtears kan vara svåra att detektera med blotta ögat men försämrar MALDI matris kristallisation och utplåna MALDI MS signal. Samma avsnitt färgas med kresylviolett avslöjar microtears och sprickor (nedre högra fotomikrografi).

Figur 3. Det första steget i analysen av data är att visualisera olika toppar över massområde analyseras (AI). Här var striatala delar från 9 möss avbildas med MALDI MS. Visualisering av den genomsnittliga totala ipå löpande kommer att avslöja områden med iögonfallande höga eller låga jonintensiteter (pilar). Dessa områden kan påverkas av över-eller under-normalisering effekter och uppgifter snedvrida analys kompromissa resultaten. Dålig anatomiska definition av topp distributioner visar avsnitt med generellt låg topp signal-till-brus, till exempel avsnitt 3 och 9, topparna F genom I.

Figur 4. MS reproducerbarhet mellan behandlingsgrupperna kan bedömas genom att beräkna den genomsnittliga MS spår och standardfel för varje m / z-värde (skär, m / z 722 och 1749). God reproducerbarhet ger giltigt statistisk analys.
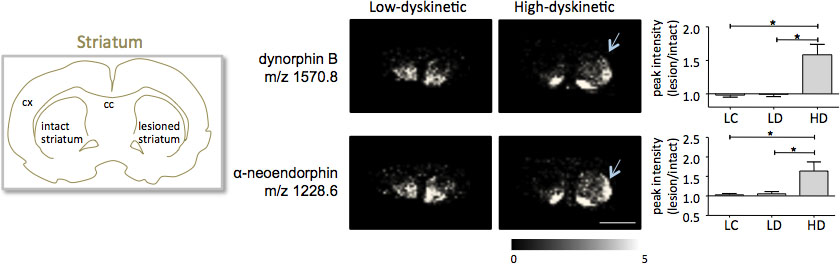
Figur 5. Dynorfin B och alfa-neoendorphin Toppintensiteterna signifikant förhöjd i 6-OHDA-Skadade, parkinsonistisk, striatum med hög dyskinetiska djur (HD, pilar) jämfört med låg-dyskinetiska (LD) och lesion kontrollgrupp (LC). Peptid toppintensiteter uttryckt som genomsnittlig utfällbara förändring av intakt sidan ± SEM (lesion / intakt sida). * P <0,05; cx kortex; cc corpus callosum. Skalstrecket 5 mm.
Discussion
Det finns flera fördelar med att anställa MALDI imaging masspektrometri i studien av neuropeptider. En objektiv analys av MS-data kan avslöja att endast specifika hjärnkärnor, eller som i de resultat som presenteras här, där endast den dorsolaterala del av striatum är associerad med en viss patofysiologiskt tillstånd. Genom att behålla den rumsliga information är det sedan möjligt att omdefiniera områden av intresse för att utföra statistisk analys med högre känslighet och lägre variabilitet jämfört med analys av hela hjärnan sektioner eller med traditionella peptidomics studier peptid extrakt. Dessutom är det viktigt att inse MALDI IMS lätt kan detektera tidigare okända posttranslationella modifieringar, men strukturella analyser måste följa för att bestämma de exakta aminosyrapositionerna som är modifierade.
Vanliga fallgropar i att visualisera MALDI IMS uppgifter omfattar att kartlägga den högsta toppen intensitet för att en linjär optisk skala från blACK (0%) till färg (100%) för varje sektion i den experimentella serie (fig 3), i stället för kartläggning alla längs en gemensam absolut skala där 100% är den maximala toppintensiteten av alla sektioner (figur 5) . Den senare metoden medger jämförelse av gruppdata och visualisering av skillnader mellan behandlingsgrupper.
Ett stort hinder för MALDI IMS analys är tilldelningen av peptider till specifika massa toppar. On-tissue tandem-masspektrometri är ibland möjligt, men ofta visar sig ganska svårt 13,14. Vi finner att en mer traditionell metod med en förberedande fraktionering av en stark katjonbytarkromatografi, följt av omvända fas LC-MS/MS kan användas för att framgångsrikt sekvens många neuropeptider och speciellt opioida peptider. Det är fortfarande inte ovanligt att få bra kvalitet MS / MS-spektra som inte matchade inga databasposter med vanliga sökmotorer såsom maskot. I dessa fall de novo-sekvensering för hand är only alternativ. Den ultimata beviset på topp identitet kan erhållas genom MALDI IMS av vävnadssnitt från lämplig knockout musen, men det är inte alltid tillgänglig eller möjlig. Ett alternativ är att validera resultaten genom en diametralt annan metod, till exempel genom Western immunoblotting eller immunohistokemi. Detta kan ofta inbegripa upplysning antikroppar och en betydande mängd arbete validera nya antikroppar.
Den allmänna strategin som presenteras i detta protokoll är optimerad för storskaliga neuropeptid MALDI IMS försök varav flera sektioner och experimentella förhållanden. Protokollet har optimerats speciellt för opioida peptider och kommer att få stor betydelse i framtida studier, som används inom olika forskningsområden, inklusive mekanismer som ligger bakom smärtan och den endogena responsen på läkemedel av missbruk.
Disclosures
Författarna har ingenting att lämna ut.
Acknowledgments
Vi tackar Hanna Warner för att bidra data för figur 3 och professor Jonas Bergquist för värdefulla synpunkter. Den svenska Vetenskapsrådet (Grant 522-2006-6416 (MA), 521-2007-5407 (MA), Den Åke Wibergs stiftelse (MA, JH), Kungliga Svenska Vetenskapsakademien MA (, JH) och den svenska kemiska Society (JH) är tacksamt känd för ekonomiskt stöd.
References
- Obeso, J. A., Olanow, C. W., Nutt, J. G. Levodopa motor complications in Parkinson's disease. Trends Neurosci. 23, S2-S7 (2000).
- Caprioli, R. M., Farmer, T. B., Gile, J. Molecular imaging of biological samples: localization of peptides and proteins using MALDI-TOF MS. MALDI-TOF MS. Anal. Chem. 69, 4751-4760 (1997).
- Obeso, J. A. The evolution and origin of motor complications in Parkinson's disease. Neurology. 55, S13-S20 (2000).
- O, W. H. Noncommunicable Diseases and Mental Health Cluster, Noncommunicable Disease Prevention and Health Promotion Department, Ageing and Life Course. Active Ageing: A Policy framework. , (2002).
- Schapira, A. H. Movement disorders: advances in cause and treatment. Lancet Neurology. , 6-7 (2010).
- Obeso, J. A., Rodriguez-Oroz, M. C., Rodriguez, M., DeLong, M. R., Olanow, C. W. Pathophysiology of levodopa-induced dyskinesias in Parkinson's disease: problems with the current model. Ann. Neurol. 47, S22-S32 (2000).
- Cenci, M. A., Lee, C. S., Bjorklund, A. L-DOPA-induced dyskinesia in the rat is associated with striatal overexpression of prodynorphin- and glutamic acid decarboxylase mRNA. Eur. J. Neurosci. 10, 2694-2706 (1998).
- Andersson, M., Hilbertson, A., Cenci, M. A. Striatal fosB expression is causally linked with l-DOPA-induced abnormal involuntary movements and the associated upregulation of striatal prodynorphin mRNA in a rat model of Parkinson's disease. Neurobiol Dis. 6, 461-474 (1999).
- Hanrieder, J. Alterations of striatal neuropeptides revealed by imaging mass spectrometry. Molecular & Cellular Proteomics. , (2011).
- Cornett, D. S., Reyzer, M. L., Chaurand, P., Caprioli, R. M. MALDI imaging mass spectrometry: molecular snapshots of biochemical systems. Nat. Methods. 4, 828-833 (2007).
- Ljungdahl, Imaging Mass Spectrometry Reveals Elevated Nigral Levels of Dynorphin Neuropeptides in L-DOPA-Induced Dyskinesia in Rat Model of Parkinson's Disease. PLoS ONE. 6, e25653 (2011).
- Groseclose, M. R., Andersson, M., Hardesty, W. M., Caprioli, R. M. Identification of proteins directly from tissue: in situ tryptic digestions coupled with imaging mass spectrometry. J. Mass. Spectrom. 42, 254-262 (2007).
- Andersson, M., Groseclose, M. R., Deutch, A. Y., Caprioli, R. M. Imaging mass spectrometry of proteins and peptides: 3D volume reconstruction. Nat. Methods. 5, 101-108 (2008).
- Deininger, S. -O. Imaging Mass Spectrometry. Setou, M. , Springer. Japan. 199-208 (2010).
- Norris, J. L. Processing MALDI Mass Spectra to Improve Mass Spectral Direct Tissue Analysis. Int. J. Mass. Spectrom. 260, 212-221 (2007).
- Ihaka, R., Gentleman, R. R. A Language for Data Analysis and Graphics. Journal of Computational and Graphical Statistics. 5, 299-314 (1996).
- Mass Spectrometry Binning Software GAB. , Vanderbilt Center for Quantitative Sciences. Nashville, TN. Available from: http://www.vicc.org/biostatistics/software.php (2012).
- Tusher, V. G., Tibshirani, R., Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. U.S.A. 98, 5116-5121 (2001).
- Bergstrom, L., Christensson, I., Folkesson, R., Stenstrom, B., Terenius, L. An ion exchange chromatography and radioimmunoassay procedure for measuring opioid peptides and substance P. Life. Sci. 33, 1613-1619 (1983).
- Falth, M. Neuropeptidomics strategies for specific and sensitive identification of endogenous peptides. Mol. Cell. Proteomics. 6, 1188-1197 (2007).
- Falth, M. SwePep, a database designed for endogenous peptides and mass spectrometry. Mol. Cell. Proteomics. 5, 998-1005 (2006).
